3.1. 光照强度对海滨雀稗生长与形态特征的响应
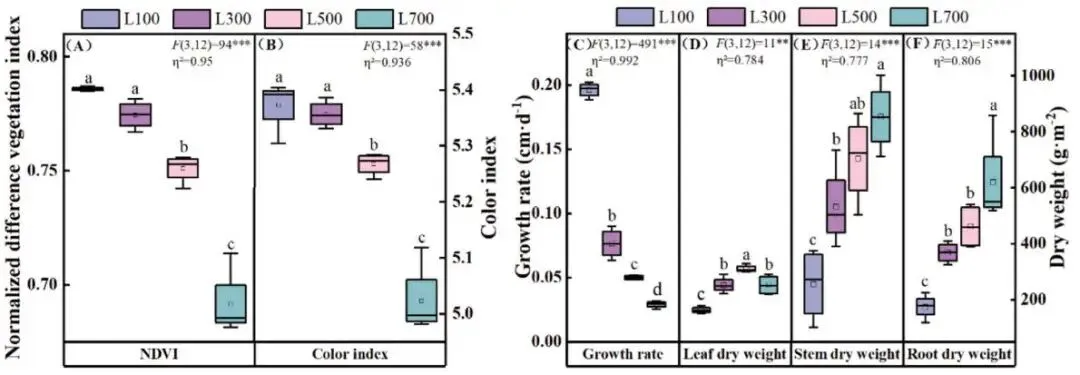
草坪质量:随光强增加,NDVI和颜色指数下降,草坪质量降低。
生长速率:随光强增加,生长速率显著下降。
生物量分配:叶干重在L500最高,茎和根干重在L700最高。
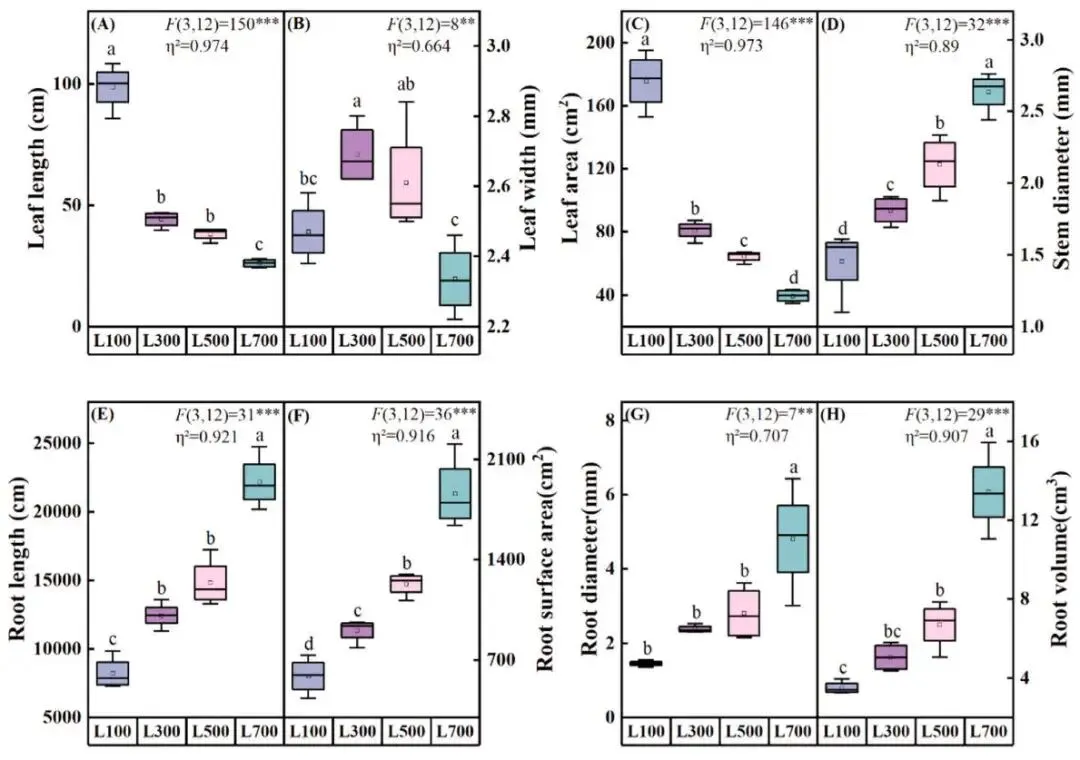
地上形态:叶长和叶面积随光强增加而减小,茎粗则增加。
地下形态:根长、根表面积、根直径和根体积随光强增加而显著增加。
随着光照强度从L100增加到L700,海滨雀稗的草坪质量(以NDVI和颜色指数衡量)和生长速率均呈现显著下降趋势。在生物量分配上,叶干重在L500光强下达到峰值,而茎干重和根干重则在L700光强下最高,表明高光促进了资源向支持性结构和地下部分的分配。形态学上,高光导致叶片变短、叶面积减小,但茎粗显著增加,以增强结构支撑。同时,所有根系参数(总根长、表面积、直径和体积)均随光强增加而显著增长,在L700下达到最大值,显示出强大的地下系统发育。L100的低光条件显著限制了植株各器官的生长 (Fig. 1, Fig. 2)。
图1. 不同光照强度下海滨雀稗的生长响应。草坪质量 (A),生长速率和干重 (B)。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
图2. 不同光照强度下海滨雀稗形态特征的变化。叶长 (A)、叶宽 (B)、叶面积 (C)、茎粗 (D)、根长 (E)、根表面积 (F)、根直径 (G)、根体积 (H)。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.2. 光照强度对海滨雀稗叶片解剖结构的响应
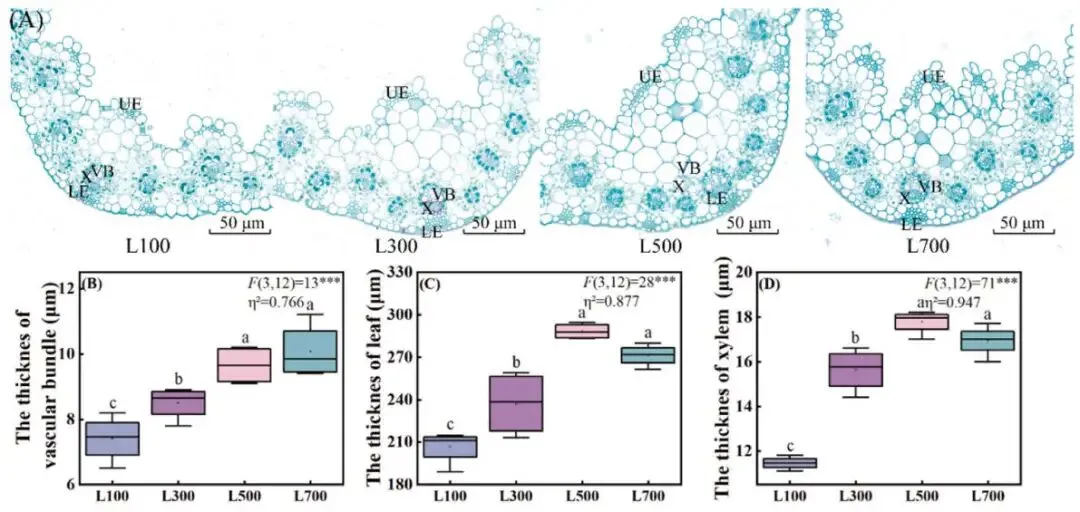
光照强度的增加引起了海滨雀稗叶片内部结构的适应性变化。维管束厚度随着光强的增加而持续增厚,在L700处理下达到最大值,相比L100增加了36.15% (P < 0.05)。而叶片厚度和木质部厚度则在L500处理下达到峰值,相较于L100分别显著增加了39.63%和55.24% (P < 0.05),随后在L700处理下略有下降。这表明中高光强(L500)最有利于叶片和木质部的结构性强化 (Fig. 3)。
图3. 不同光照强度下叶片解剖结构的变化。(A) 解剖结构图,(B) 维管束厚度,(C) 叶片厚度,(D) 木质部厚度。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.3. 光照强度对海滨雀稗叶片光合相关参数的响应
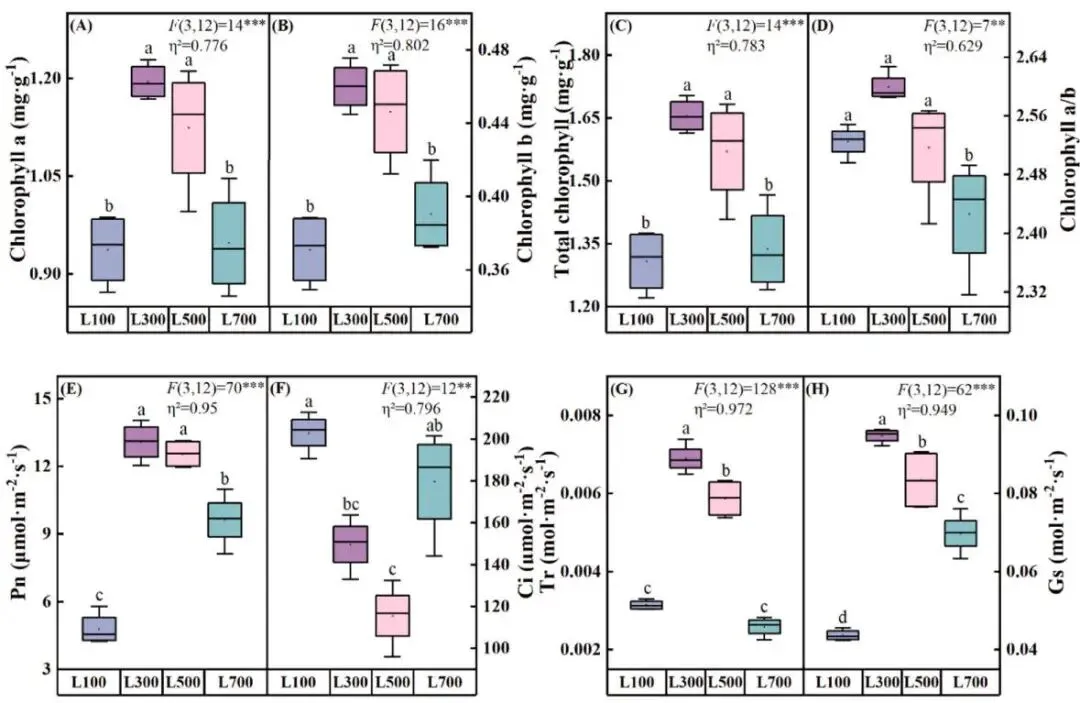
海滨雀稗的光合参数对光强变化响应显著。叶绿素a、叶绿素b及总叶绿素含量均在L300处理下达到峰值。在L700光强下,叶绿素a/b比值显著低于其他处理 (P < 0.05)。同样,净光合速率、蒸腾速率和气孔导度在L300处理下最高。值得注意的是,胞间CO₂浓度在L500处理下达到最低水平,表明此时CO₂利用效率最高。除蒸腾速率和气孔导度外,L300和L500处理间的光合相关参数无显著差异 (P > 0.05),说明海滨雀稗在L500光强下仍保持着较高的光合能力 (Fig. 4)。
图4. 不同光照强度下海滨雀稗叶绿素a (A)、叶绿素b (B)、总叶绿素 (C)、叶绿素a/b (D)以及净光合速率(Pn) (E)、胞间CO₂浓度(Ci) (F)、蒸腾速率(Tr) (G)和气孔导度(Gs) (H)的变化。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.4. 光照强度变化对海滨雀稗叶片碳氮浓度的响应
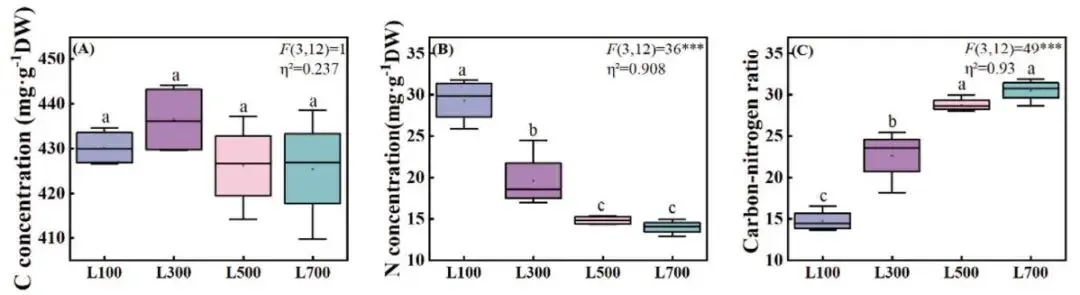
不同光强处理下,海滨雀稗叶片的碳(C)浓度没有显著差异 (P > 0.05)。然而,氮(N)浓度随光强的增加呈现出明显的下降趋势,从L100处理下的最高值(29.30 mg·g⁻¹)显著降低至L500和L700处理下的14.82和14.00 mg·g⁻¹。与此相反,碳氮比(C/N)随光强增加而显著上升,在L500和L700处理下分别比L100处理高出94.75%和106.26% (Fig. 5)。
图5. 不同光照强度下海滨雀稗叶片C浓度 (A)、N浓度 (B)、碳氮比 (C) 的变化。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.5. 光照强度对海滨雀稗叶片碳水化合物的影响
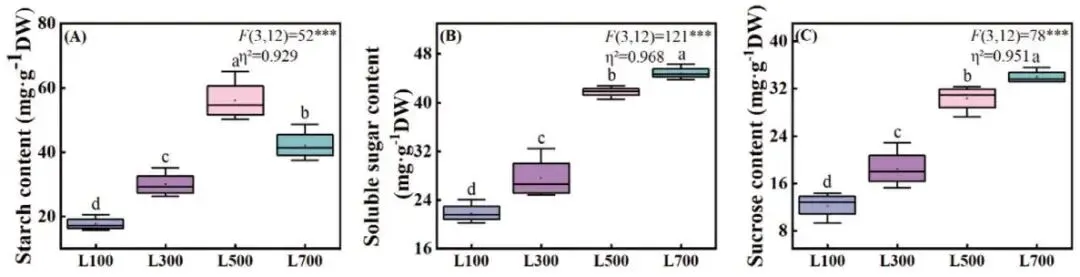
光照强度显著影响了海滨雀稗叶片中碳水化合物的分配。淀粉含量随着光强的增加呈现先升高后降低的趋势,在L500处理下达到最高值(56.04 mg·g⁻¹)。相反,可溶性糖和蔗糖的含量则从L100到L700持续显著上升 (P < 0.05),在L700处理下分别达到44.85和33.96 mg·g⁻¹,表明高光胁迫下碳水化合物由储存形式(淀粉)向渗透调节物质(可溶性糖)转化 (Fig. 6)。
图6. 不同光照强度下海滨雀稗叶片碳水化合物的变化。淀粉含量 (A)、可溶性糖含量 (B) 和蔗糖含量 (C)。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.6. 光照强度对海滨雀稗叶片细胞壁组分的响应
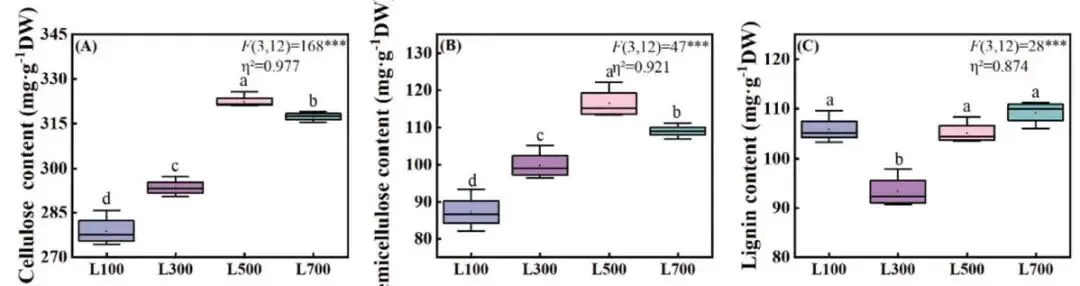
细胞壁结构组分的含量同样受光强调控。纤维素和半纤维素的含量随光强增加呈现先升高后降低的模式,均在L500处理下达到峰值,分别为322.38和116.37 mg·g⁻¹。在L700光强下,二者含量相比L500有显著下降 (P < 0.05)。此外,L300处理下的木质素含量显著低于其他三种光强处理 (P < 0.05),表明中等光强(L300)可能不利于木质化进程 (Fig. 7)。
图7. 不同光照强度下海滨雀稗叶片细胞壁组分含量的变化。纤维素含量 (A)、半纤维素含量 (B) 和木质素含量 (C)。所有数据以平均值 ± 标准差 (n = 4) 表示。柱上不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.7. 光照强度对海滨雀稗抗氧化酶的响应
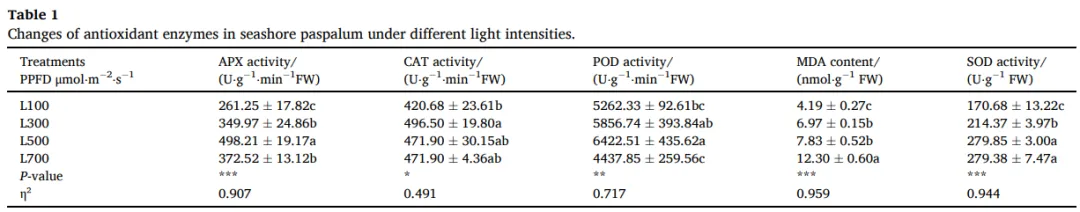
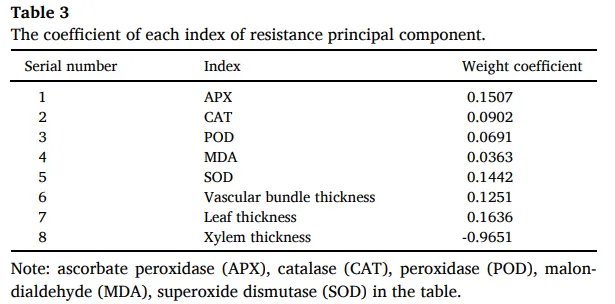
随着光强增加,海滨雀稗的抗氧化系统被激活。抗坏血酸过氧化物酶(APX)和过氧化物酶(POD)的活性在L500处理下达到峰值,而超氧化物歧化酶(SOD)的活性在L500和L700处理下显著高于低光处理。然而,在L700光强下,APX和POD活性出现显著下降,同时膜脂过氧化产物丙二醛(MDA)的含量达到最高水平,这表明在L700的强光胁迫下,抗氧化系统的清除能力已不足以应对过量的活性氧,导致了氧化损伤 (Table 1)。
表1. 不同光照强度下海滨雀稗抗氧化酶的变化。 注:APX(抗坏血酸过氧化物酶)、CAT(过氧化氢酶)、POD(过氧化物酶)、MDA(丙二醛)、SOD(超氧化物歧化酶)。所有数据以平均值 ± 标准差 (n = 4) 表示。同行不同的小写字母表示基于单因素方差分析和LSD事后检验,处理间存在显著差异 (P < 0.05)。星号表示单因素方差分析的显著性水平 (* P < 0.05, ** P < 0.01, *** P < 0.001)。η² 代表偏eta平方值,表示效应大小。
3.8. Spearman相关性分析
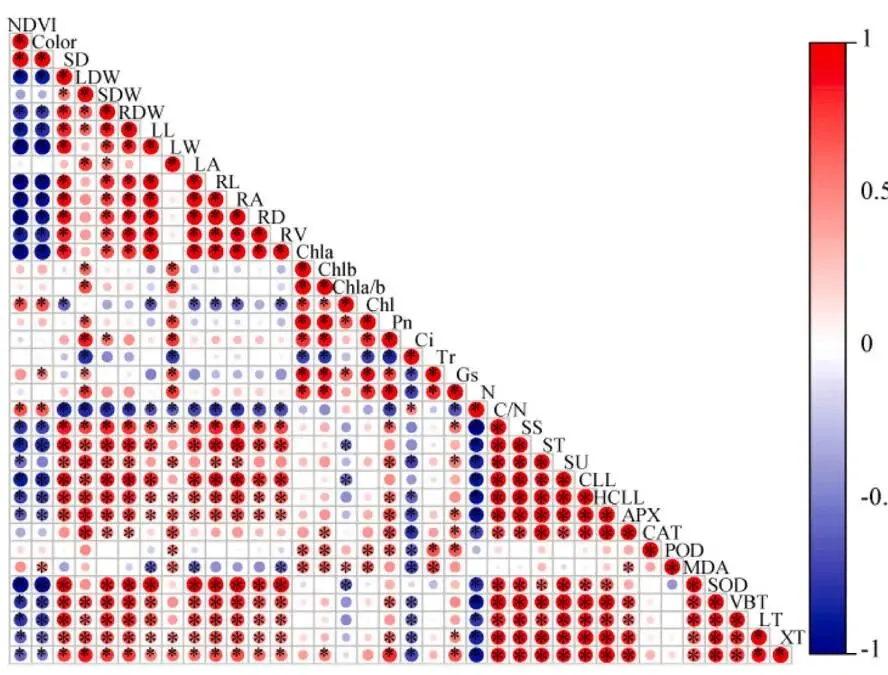
Spearman相关性分析揭示了各指标间的复杂关系。值得注意的是,草坪质量(NDVI, Color)与大部分生长指标(如干重、形态参数)和抗性指标(如碳水化合物、细胞壁组分、抗氧化酶、解剖结构)均呈显著负相关,但与总氮含量呈显著正相关。胞间CO₂浓度与多数其他指标呈负相关。此外,总氮含量与除草坪质量外的几乎所有指标均呈负相关,而碳氮比则呈现相反的模式。这些结果进一步支持了碳氮关系在调控海滨雀稗生长-抗性权衡中可能扮演关键角色的假设 (Fig. 8)。
图8. 不同光照强度下海滨雀稗各指标参数的Spearman相关性。NDVI (归一化植被指数), Color (颜色指数), SD (茎粗), LDW (叶干重), SDW (茎干重), RDW (根干重), LL (叶长), LW (叶宽), LA (叶面积), RL (根长), RA (根表面积), RD (根直径), RV (根体积), Chla (叶绿素a), Chlb (叶绿素b), Chla/b (叶绿素a/b), Chl (总叶绿素), Pn (净光合速率), Ci (胞间CO₂浓度), Tr (蒸腾速率), Gs (气孔导度), N (N浓度), SS (可溶性糖), ST (淀粉), SU (蔗糖), CLL (纤维素), HCLL (半纤维素), APX (抗坏血酸过氧化物酶), CAT (过氧化氢酶), POD (过氧化物酶), MDA (丙二醛), SOD (超氧化物歧化酶), VBT (维管束厚度), LT (叶片厚度), XT (木质部厚度), * 表示显著性 (P < 0.05)。
3.9. ‘生长’与‘抗性’的量化及其影响路径分析
主成分量化:通过PCA构建了‘生长’和‘抗性’两个主成分。
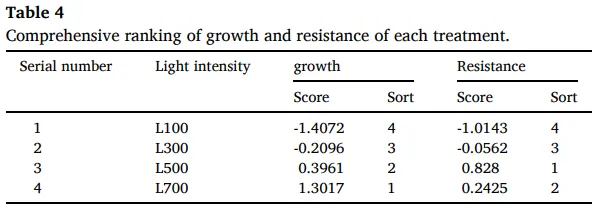
生长抗性权衡:生长主成分在L700最高,抗性主成分在L500最高。
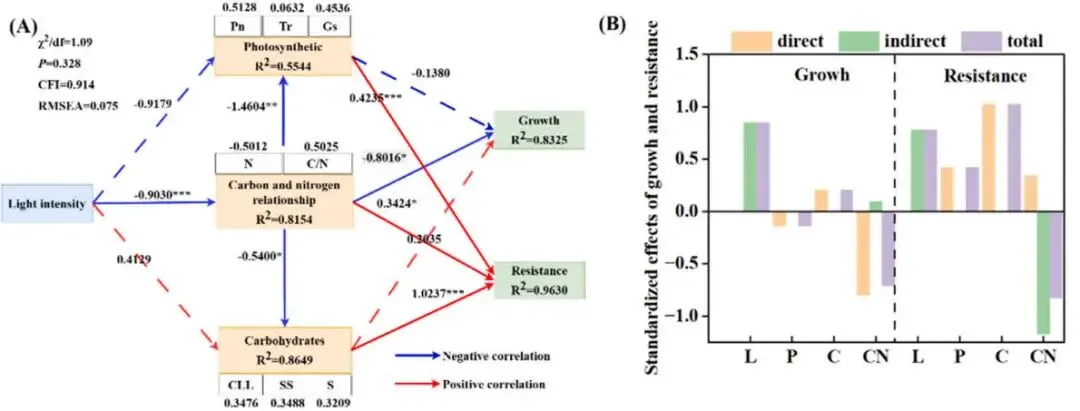
路径模型分析:SEM模型显示,碳氮关系对生长有直接负效应,碳水化合物对康性有最高的直接正效应。
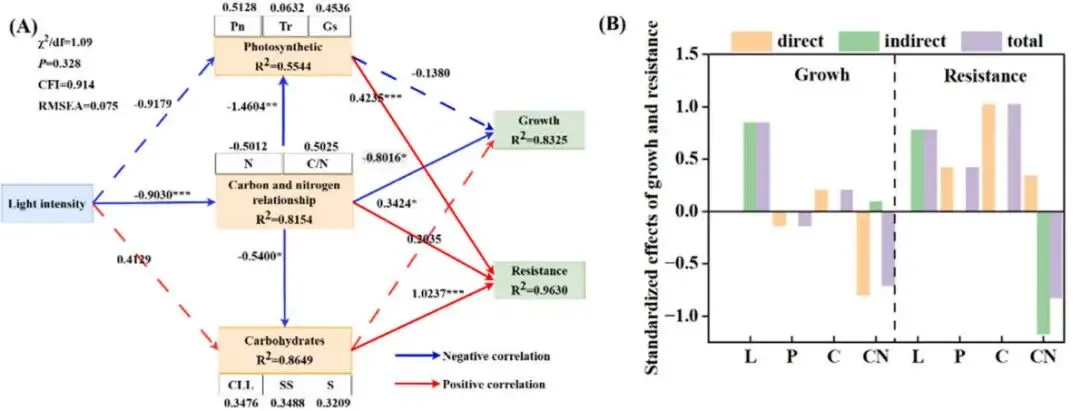
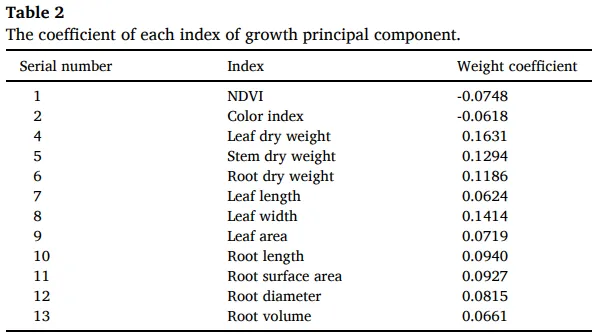
通过主成分分析(PCA),将多个指标整合为‘生长’(由NDVI、颜色指数、干重、茎粗和形态性状构成)和‘抗性’(由APX, CAT, POD, MDA, SOD、维管束厚度、叶片厚度和木质部厚度构成)两个主成分。综合评分显示,‘生长’的最优处理为L700,而‘抗性’的最优处理为L500,这表明从L100到L500,生长与抗性协同增强,但在更高光强下二者发生了解耦与权衡。结构方程模型(SEM)分析进一步揭示了调控路径,模型拟合良好。结果显示,光强通过‘光合’、‘碳水化合物’和‘碳氮关系’三个路径影响生长和抗性。其中,‘碳氮关系’对‘生长’有显著的直接负向影响(-0.8016),而‘碳水化合物’对‘抗性’有最大的直接正向影响(1.0236),证实了碳氮平衡与碳水化合物代谢在介导光强诱导的生长-抗性权衡中的核心作用 (Table 2, Table 3, Table 4, Fig. 9)。
表2. 生长主成分各指标的系数。
表3. 抗性主成分各指标的系数。 注:表中APX(抗坏血酸过氧化物酶)、CAT(过氧化氢酶)、POD(过氧化物酶)、MDA(丙二醛)、SOD(超氧化物歧化酶)。
表4. 各处理生长与抗性的综合排名。
图9. 不同光照强度下海滨雀稗的结构方程模型分析 (A) 和效应 (B)。白色方框代表构成路径(黄色方框)的指标,上方数字表示各指标对应的权重系数。实线箭头表示显著路径,虚线箭头表示不显著路径,各路径显示了标准化路径系数。* 表示显著性 (* P < 0.05, ** P < 0.01, *** P < 0.001)。χ²/df (卡方自由度比), P (模型拟合卡方检验的p值), CFI (比较拟合指数), RMSEA (近似误差均方根), Pn (净光合速率), Tr (蒸腾速率), Gs (气孔导度), N (N浓度), C/N (碳氮比), CLL (纤维素), SS (可溶性糖), S (淀粉), L (光强), P (光合), C (碳水化合物), CN (碳氮关系)。
本研究首次将植物生态学中的“响应策略转变”假说系统地应用于海滨雀稗这一重要的草坪植物,填补了以往研究多集中于模式植物或园艺作物而忽略草坪植物光响应机制的理论空白。
研究在方法学上实现了突破,通过主成分分析(PCA)整合了形态、生理、解剖等数十个指标,首次构建并量化了“生长”与“抗性”两大综合主成分,为科学评估植物复杂的生长-抗性权衡提供了全新的定量框架。
本研究开创性地运用结构方程模型(SEM),构建并验证了一个包含“光合”、“碳水化合物”和“碳氮关系”三个潜在中介路径的理论模型,成功揭示了光照强度调控海滨雀稗生长与抗性权衡的具体作用途径及其相对重要性。
研究明确指出了碳氮关系是调控光胁迫下生长与抗性权衡的核心枢纽,发现其对生长具有最强的直接负向效应,为理解植物在胁迫下的资源分配策略提供了关键的实验证据。
本研究精准地确定了海滨雀稗生长与抗性协同增强的最佳光照强度阈值(500 μmol·m⁻²·s⁻¹),并揭示了在更高光强下二者发生解耦与权衡的具体生理与代谢变化,为海滨雀稗的精准光环境管理提供了直接的理论依据。
- 随着光照强度增加,海滨雀稗从主动适应转变为防御响应。
- 结构方程模型(SEM)揭示碳氮关系主导了生长与抗性调控。
- 在500 μmol·m⁻²·s⁻¹光照强度下,淀粉、纤维素、半纤维素含量及叶片厚度、叶干重、抗氧化酶活性达到最高,实现了生长与抗性的协同增强。
- 在700 μmol·m⁻²·s⁻¹光照强度下,可溶性糖、MDA含量达到峰值,但淀粉含量显著下降,叶片氮浓度降低,碳氮比显著升高,导致抗性下降。
- 光照强度通过光合作用能力、碳水化合物代谢和碳氮关系三条途径调节生长与抗性权衡。
- 适度的光照强度(500 μmol·m⁻²·s⁻¹)通过维持碳氮平衡优化碳分配,促进地上生长和抗氧化系统。
- 高光照强度(700 μmol·m⁻²·s⁻¹)打破代谢平衡,增加碳资源分配至可溶性糖和损伤修复,抑制抗氧化酶活性,增加地下干重,牺牲地上生长和草坪质量以应对光胁迫。
- 研究结果为优化光管理和抗逆性栽培提供了理论依据,同时提出了在不同光照强度下的氮肥管理建议。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
