2026 年 3 月 19 日,来自吉林农业大学的Hang Yang等人在Journal of Agricultural and Food Chemistry上发表了一篇题为Rational Design of Flexible Regions in a GH10 Xylanase from Paecilomyces aerugineus to Improve Its Thermostability, Catalytic Efficiency, and XOS Production from Corn Stover的研究性论文。Abstract
为了提升来源于Paecilomyces aerugineus的GH家族10木聚糖酶(PaXyn10A)的热稳定性和催化效率,本研究开展了理性设计。通过分子动力学模拟和序列比对来识别柔性区域并规避功能冲突位点,获得了突变体PaXyn10A D68F/S95A。与PaXyn10A相比,PaXyn10A D68F/S95A的最适温度提高了5°C,在50°C下的热失活半衰期延长了11.5倍。同时,PaXyn10A D68F/S95A的比活力提高了2.1倍,催化效率提高了3.9倍。在最优条件下,通过PaXyn10A D68F/S95A水解与蒸汽爆破相结合,从玉米秸秆中获得的低聚木糖(XOS)得率达到64.27%。这些结果表明,通过计算机模拟指导、利用理性点突变对柔性区域进行刚性化改造,是提高GH10木聚糖酶热稳定性和催化效率的有效策略。
Introduction
不可再生资源的过度使用和不可持续的工业实践已导致全球污染危机日益恶化,凸显了开发环境友好型替代品的迫切需求。木质纤维素作为地球上最丰富的可再生资源之一,主要由纤维素、半纤维素和木质素组成,其高效转化与利用是生物炼制领域的核心挑战。传统的化学方法,如酸处理和碱处理,会产生较多副产物并造成严重的设备腐蚀,从而推高工艺成本。此外,酸碱的使用还可能增加木质素的缩合程度,不利于后续的酶水解和木质素转化。与化学法相比,木质纤维素的酶法降解具有特异性更高、反应条件更温和、副产物更少等优势。木聚糖酶能随机断裂木聚糖的β-1,4-糖苷键,生成木糖和低聚木糖(XOS),这在生物能源和生物基材料的开发中具有重要意义。XOS具有显著的益生元特性,能促进有益肠道菌群的增殖,有助于维持肠道健康。因此,XOS已广泛应用于食品、饲料和医药行业。据估计,全球每年产生约3.1亿吨玉米秸秆。鉴于其木聚糖含量约为20%(w/w),玉米秸秆是生产XOS极具潜力的原料。木质纤维素的降解在高温和酸性条件下更为高效,这能加快反应速率、降低成本并防止微生物污染。这对木聚糖酶的耐酸性和热稳定性提出了更高要求。然而,大多数酸性木聚糖酶的热稳定性较差。因此,通过酶工程提高热稳定性对于工业应用至关重要。
目前,提高木聚糖酶热稳定性主要通过定向进化、理性设计或半理性设计方法实现。计算机辅助技术的进步有助于更好地理解蛋白质的结构-功能关系。分子对接、分子动力学模拟和虚拟饱和突变等技术显著提高了酶工程的效率。增强蛋白质的结构刚性可提高其热稳定性。这通常涉及两个关键环节:预测柔性区域和实现结构刚性化。分子动力学(MD)模拟能够快速准确地预测蛋白质中的柔性区域。通过MD模拟,定位了木聚糖酶XynA N端的五个柔性区域。针对这些柔性区域进行定点饱和突变,获得了最适温度比野生型高20°C的突变体。基于吉布斯自由能变化(ΔΔG)的虚拟饱和突变有助于高通量筛选潜在的稳定化突变位点。通过结合保守序列分析和虚拟饱和突变,获得了比活力为野生型酶9.3倍、半衰期为130.9倍的突变体。通过序列比对定位耐热木聚糖酶中的保守残基,有助于计算机引导的柔性区域理性设计。因此,快速准确地识别高柔性区域中的潜在刚性化位点,并基于序列保守性分析构建双突变体,是提高GH10木聚糖酶热稳定性的可行策略。
在我们此前的研究中,来自Paecilomyces aerugineus的GH10木聚糖酶PaXyn10A被克隆和表达。PaXyn10A表现出优异的耐酸性和水解活性。然而,作为一种中温酶,PaXyn10A的热稳定性较差,难以满足工业要求。在本研究中,首先通过分子动力学(MD)模拟预测了PaXyn10A的柔性区域,随后针对这些柔性区域内的残基进行了计算机虚拟饱和突变。基于突变诱导的去折叠自由能变化(ΔΔG)合理构建了突变体文库,并系统研究了突变体的热稳定性和催化活性。随后,通过与耐热GH10木聚糖酶进行多序列比对来定位保守区域,以尽量减少突变位点之间可能存在的上位效应。系统表征表明,与野生型酶相比,组合突变体表现出增强的热稳定性和更优的催化活性。
1. 分子动力学模拟与虚拟饱和突变
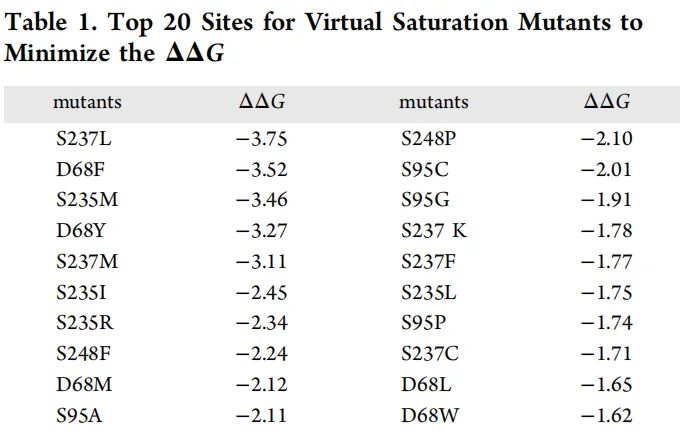
通过分子动力学模拟观察到了三个柔性区域(图1b)。基于PaXyn10A的三维结构,对柔性区域内的氨基酸进行了饱和突变。最终,获得了20个具有最低去折叠自由能(ΔΔG)的突变位点(表1)。
2. 野生型PaXyn10A及其突变体的比较
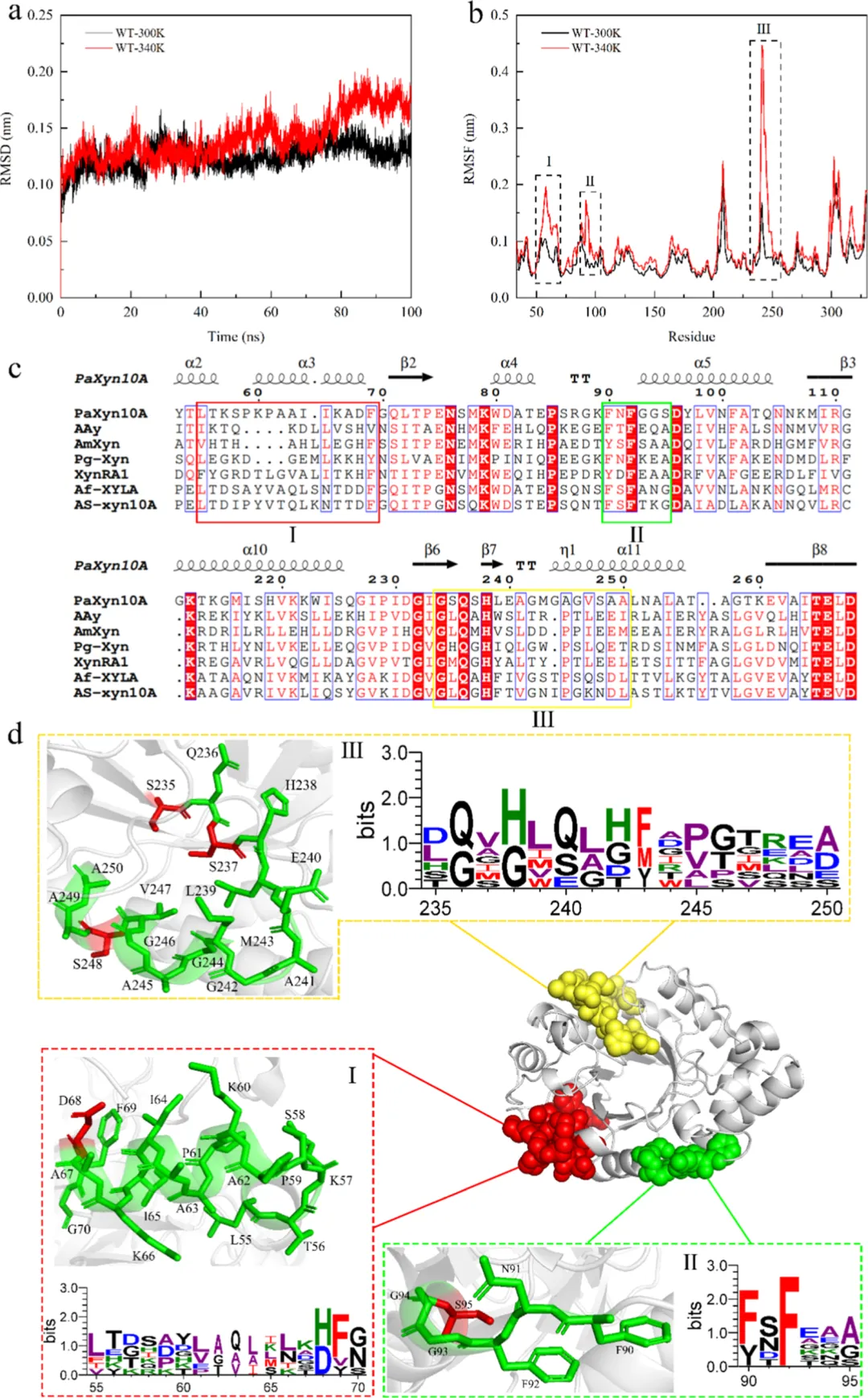
基于上述20个突变体,构建了用于酶表达的转化子。热稳定性测定结果显示,PaXyn10A在55°C下孵育30分钟后,其残余活性几乎完全丧失(6.3%)。具有木聚糖酶活性的突变体主要集中在D68、S95和S248位点。在这三个位点中,248位点的所有突变体比活力均低于PaXyn10A(248.7 ± 0.5 U/mg)。D68F突变体在比活力和残余酶活力方面均显著优于其他突变体。WebLogo分析显示,丙氨酸是95位点的主要保守氨基酸(图1d)。因此,构建了双突变体D68F/S95A以进一步改善酶学性质。
3. PaXyn10A及其突变体的表达与纯化
PaXyn10A及其突变体在大肠杆菌BL21(DH3)中成功表达,重组木聚糖酶(PaXyn10A、PaXyn10A D68F、PaXyn10A S95A和PaXyn10A D68F/S95A)通过Ni-HP柱纯化至表观均一,纯化倍数分别为3.0、5.6、5.2和5.3,回收率分别为60.9%、71.4%、83.9%和74.6%。四种木聚糖酶的分子量均约为32.0 kDa。与PaXyn10A相比,突变体的比活力也有所提高。其中,PaXyn10A D68F/S95A的比活力最高(524.5 U/mg),是野生型酶(248.7 U/mg)的2.11倍。
4. PaXyn10A及其突变体的生化表征
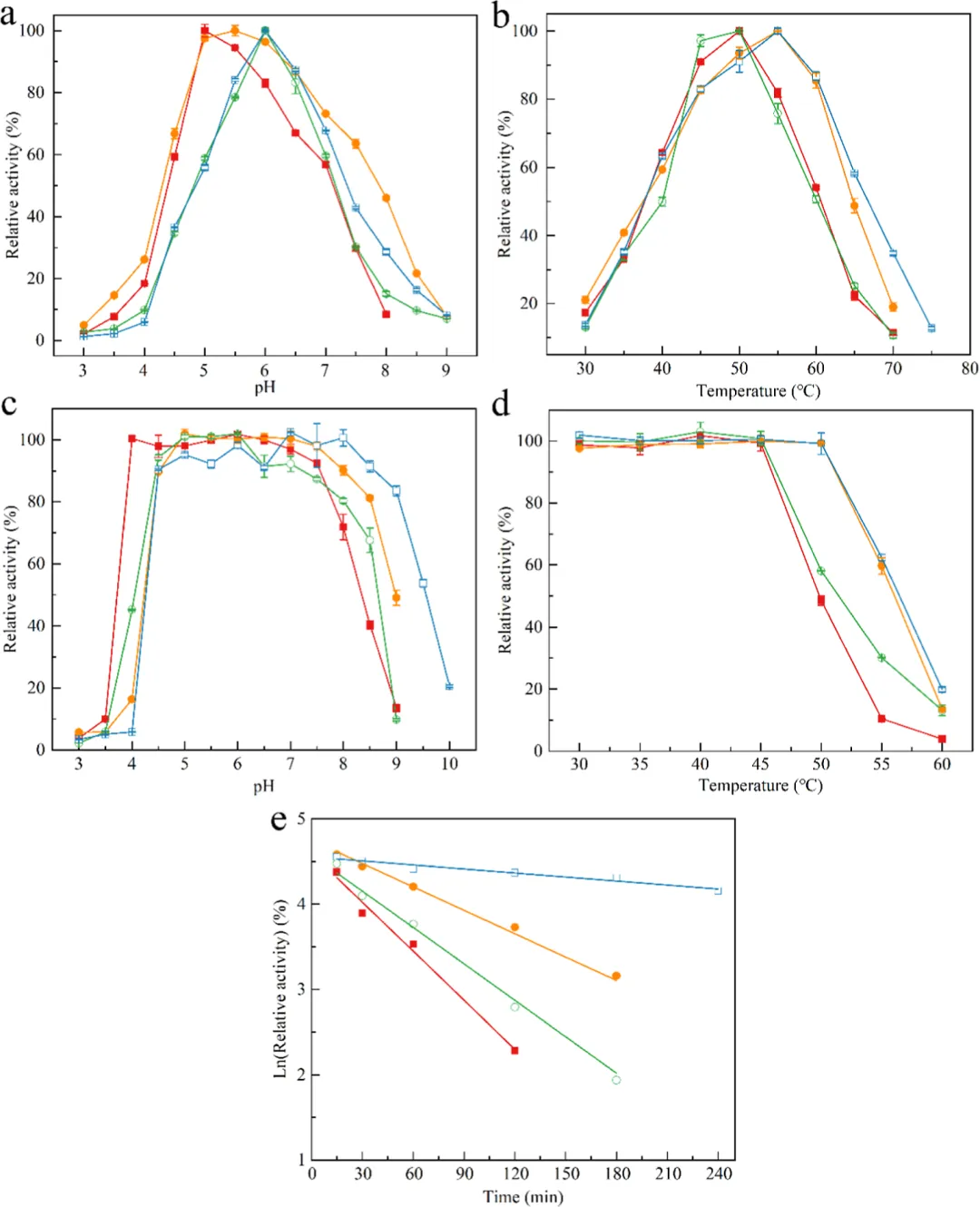
PaXyn10A、D68F、S95A和D68F/S95A的最适pH分别为5.0、5.5、6.0和6.0(图2a)。PaXyn10A在pH 4.0−7.5范围内保持80%以上的相对活性(图2c)。D68F、S95A和D68F/S95A分别在pH 4.5−8.5、4.5−8.0和4.5−9.0范围内稳定(图2c)。PaXyn10A和S95A均在50°C时表现出最大活性(图2b),且在45°C以下稳定(图2d)。而D68F和D68F/S95A均在55°C时表现出最大活性(图2b),且在50°C以下稳定(图2d)。PaXyn10A、S95A、D68F和D68F/S95A的半衰期分别为35、48、94和401分钟(图2e)。PaXyn10A D68F/S95A在55°C和60°C下的半衰期分别是PaXyn10A的8.5倍和5.9倍。
5. PaXyn10A与PaXyn10A D68F/S95A的底物特异性和动力学参数
PaXyn10A和PaXyn10A D68F/S95A对不同木聚糖底物表现出相似的底物特异性。PaXyn10A D68F/S95A对小麦阿拉伯木聚糖的底物特异性最高(2,041.7 U/mg),其次为山毛榉木木聚糖(506.4 U/mg)、燕麦木聚糖(156.4 U/mg)和桦木木聚糖(76.5 U/mg)。
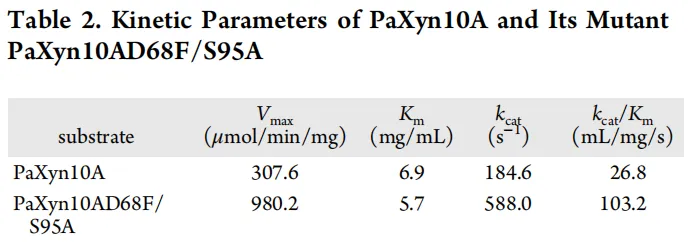
PaXyn10A的Km值(6.9 mg/mL)大于PaXyn10A D68F/S95A(5.7 mg/mL)(表2),表明突变体与底物的结合力略有增强。同时,与PaXyn10A相比,PaXyn10A D68F/S95A的Vmax和kcat均有提高,使其催化效率(kcat/Km)提高了3.9倍(表2)。
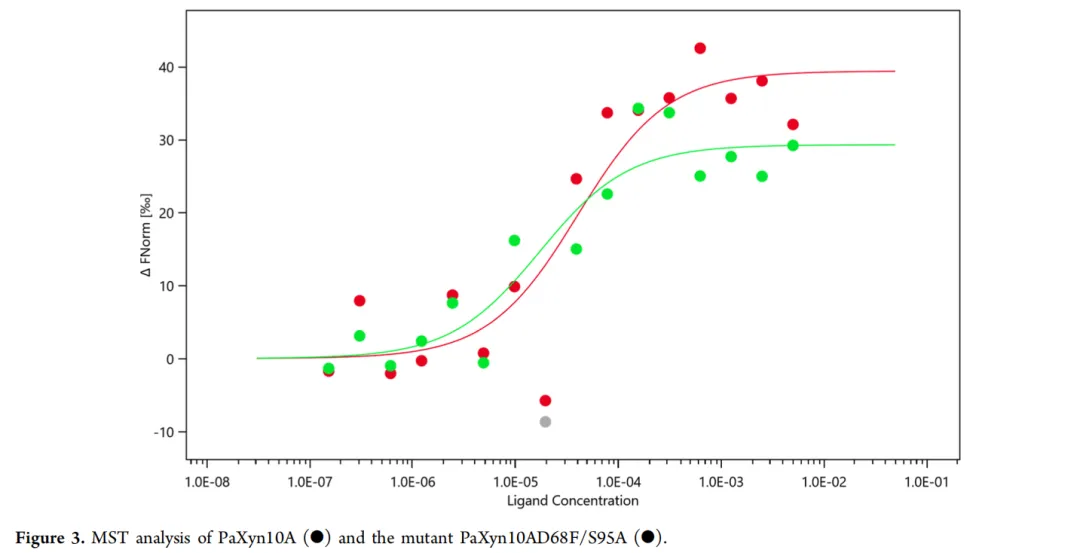
6. PaXyn10A与PaXyn10A D68F/S95A的MST分析
PaXyn10A和PaXyn10A D68F/S95A与底物的亲和力(kd)分别测定为50.0 μM和14.7 μM(图3)。结果表明,PaXyn10A D68F/S95A对底物表现出更强的亲和力,这与Km观察到的趋势一致。
7. PaXyn10A与PaXyn10A D68F/S95A的结构分析
利用SWISS-MODEL服务器,以来自P. simplicissimum的木聚糖酶(PDB ID: 1BG4;序列相似性:78.74%)为模板,对PaXyn10A和PaXyn10A D68F/S95A的三维结构进行了同源建模。通过Ramachandran图评估了建模质量。预测的三维模型显示,PaXyn10A和PaXyn10A D68F/S95A中分别有97.44%和97.00%的氨基酸残基位于Ramachandran图的允许区。结果表明,预测模型质量较高,适用于分子对接和动力学模拟。
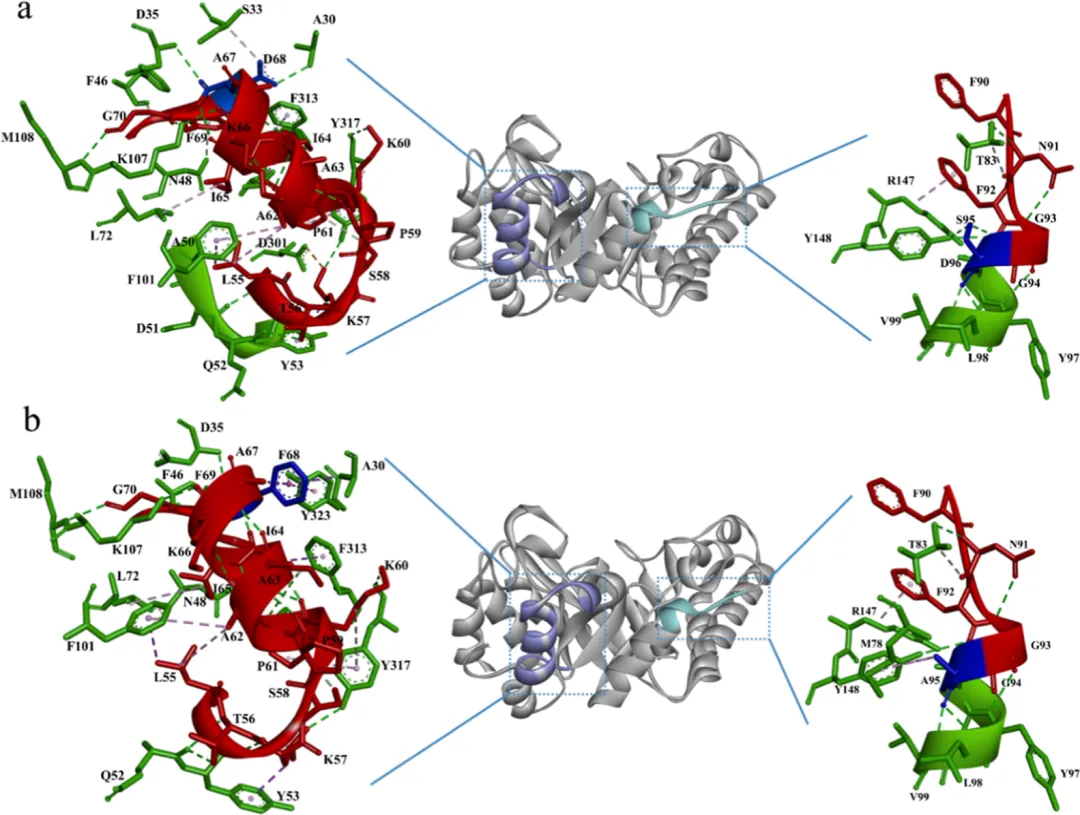
与PaXyn10A相比,PaXyn10A D68F/S95A的柔性区域之间观察到显著的相互作用变化(图4)。在N端近端区域,与PaXyn10A相比,两个氢键(A30−D68和S33−D68)丢失。PaXyn10A D68F/S95A不仅形成了一个新的氢键(D68−Y323),还建立了三个疏水相互作用(A30−D68、D68−F69和D68−Y323)(图4)。另一个突变位点(S95A)位于靠近催化沟槽的柔性区域内。与构象A相比,构象B丢失了一个氢键(S95−Y148),但获得了两个疏水相互作用(A95−Y148和M78−A95)(图4)。
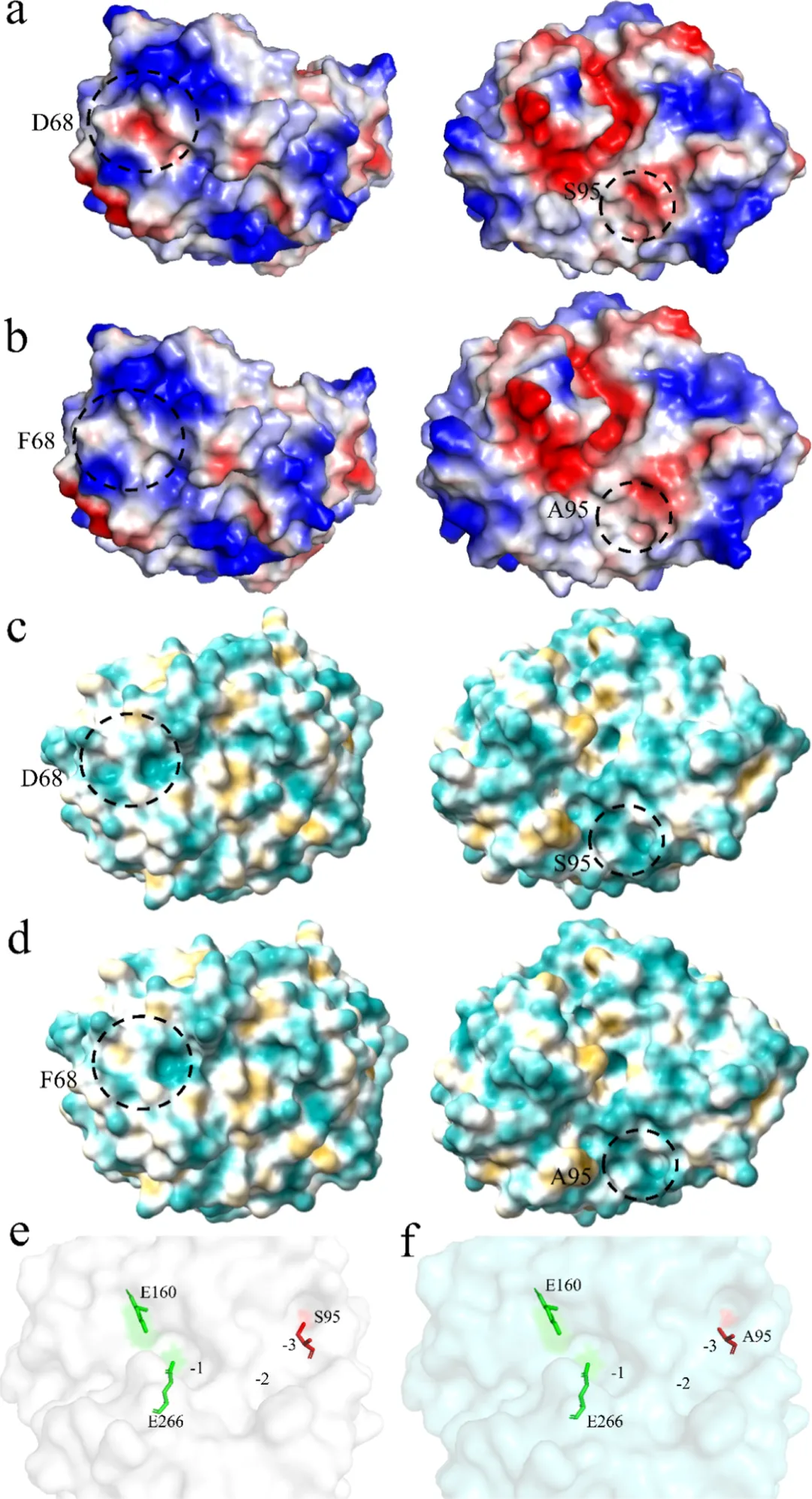
木聚糖酶的表面静电势被认为对其热稳定性和pH适应性具有重要影响。突变D68F在68位氨基酸周围引入了增强的正静电势,而S95A突变位点周围的静电势与野生型PaXyn10A非常相似(图5a,b)。此前的研究表明,增加蛋白质的净表面电荷有利于提高木聚糖酶的热稳定性。此外,68位点的突变诱导了蛋白质基底表面从亲水性向疏水性的转变(图5c,d)。
分子对接结果显示,PaXyn10A和PaXyn10A D68F/S95A的结合自由能分别为−4.16和−4.25 kcal/mol。与PaXyn10A相比,突变体D68F/S95A的底物与残基Asn91和Tyr53之间分别建立了新的氢键网络,这改善了糖苷键与催化残基的对齐,并降低了断键所需的活化能。值得注意的是,Ala95与相邻残基如Phe92和Gly93协同作用,增强了底物非极性骨架周围的疏水堆积,使其稳定在更有利于催化的构象中。这可能归因于Ala95的小非极性侧链消除了野生型酶中与Ser95存在的直接相互作用,从而增加了底物进入和产物释放的可用空间。总体而言,这些修饰优化了底物结合并加速了转换,从而提升了整体催化性能。
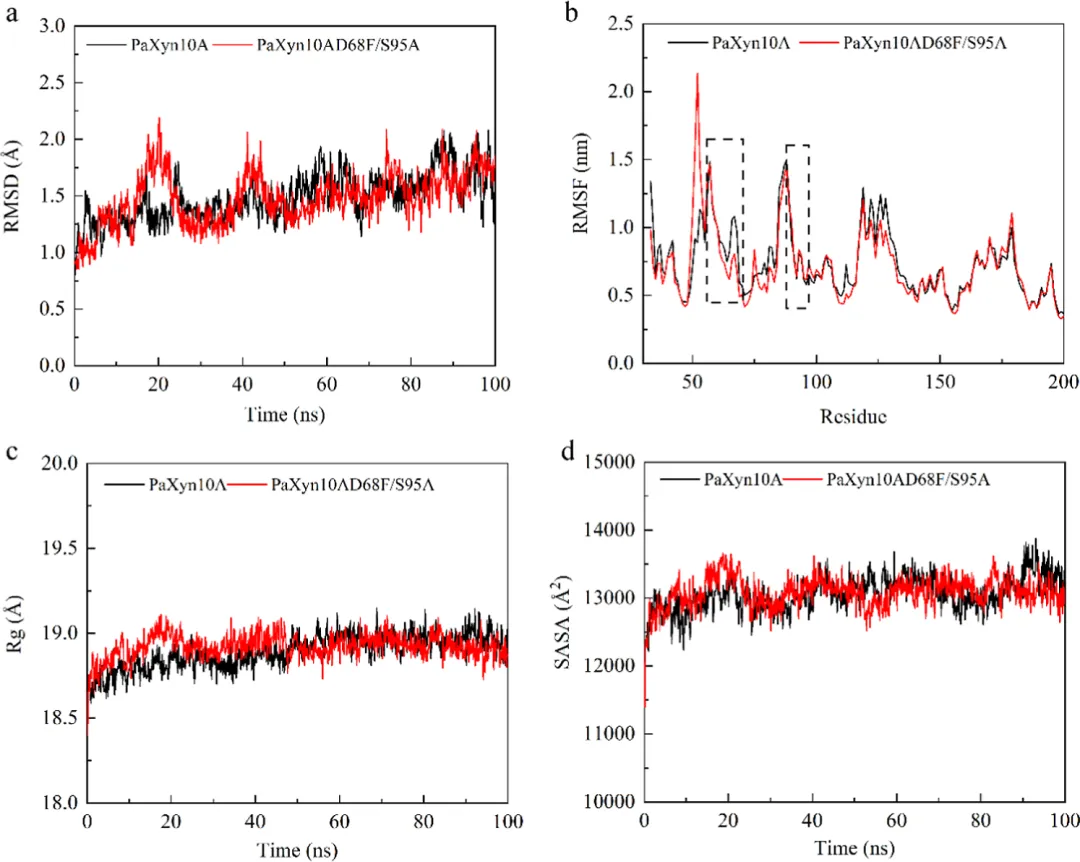
RMSD能够清晰地指示MD模拟过程中蛋白质的结构稳定性和构象变化。在40−100 ns期间,PaXyn10A D68F/S95A的RMSD值低于PaXyn10A,表明PaXyn10A D68F/S95A相对于其初始构象发生了较小的构象变化,并且具有更高的热稳定性(图6a)。RMSF结果显示,D68F突变的引入导致柔性区域I(残基55−70)的RMSF显著降低,证实了该突变有效增强了该区域的刚性(图6b)。Rg反映了蛋白质结构的整体大小和致密性。Rg值与结构刚性呈负相关。PaXyn10A D68F/S95A的Rg低于PaXyn10A,表明PaXyn10A D68F/S95A的结构更加紧凑,这与蛋白质的稳定性直接相关(图6b)。更紧凑和稳定的结构限制了不必要的构象波动,防止了催化残基的空间偏移,从而维持了高效催化所必需的底物结合裂缝的几何构型。其次,它减少了底物结合和催化重排过程中的不利熵罚,从而直接提高了催化效率(kcat/Km)。最后,它稳定了多亚位点结合裂缝两侧的柔性环区,维持了结合亚基的保守微环境,以促进木聚糖底物的特异性和持续性结合,并进一步增强催化转换效率。
8. PaXyn10A与PaXyn10A D68F/S95A的水解特性及玉米秸秆中XOS的生产
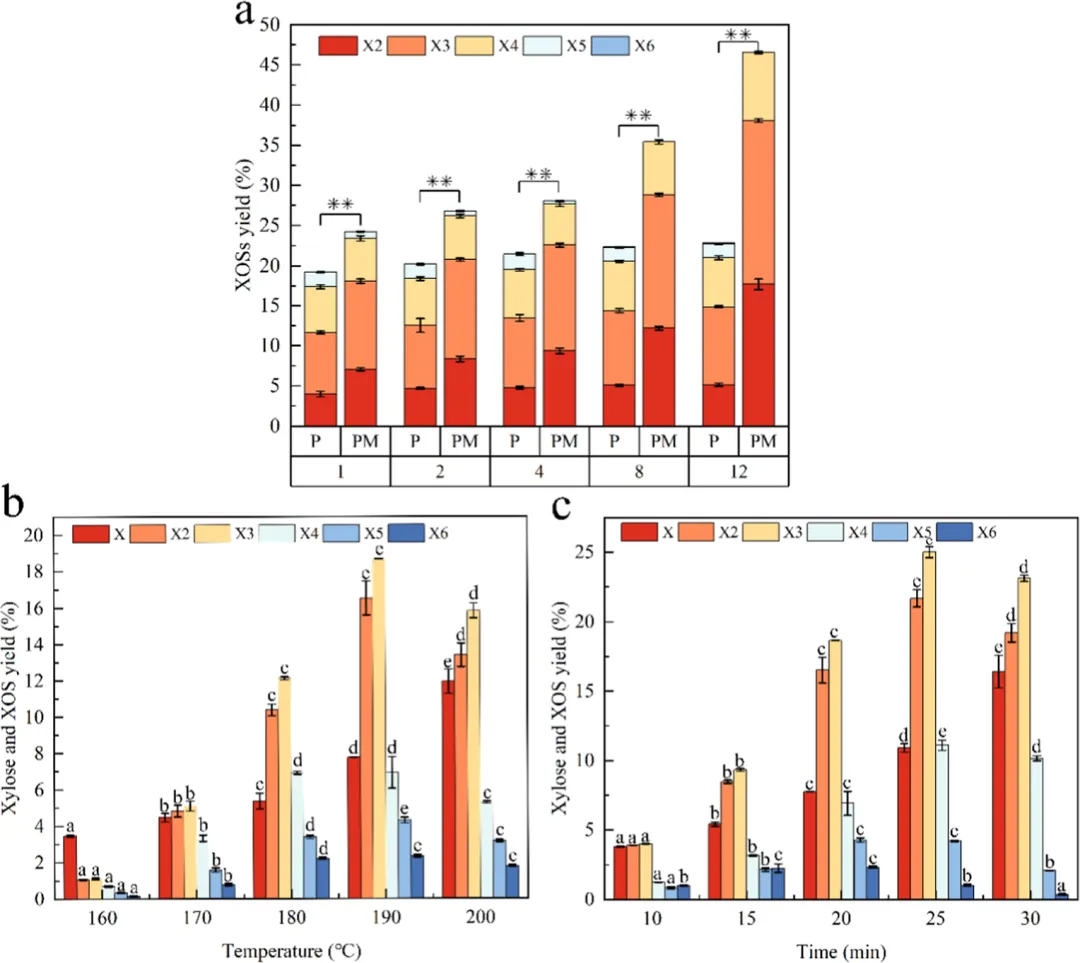
PaXyn10A D68F/S95A的水解特性与PaXyn10A相似,主要产物均为X2−X4和X2−X4。12小时后,PaXyn10A主要生成X2−X5,而PaXyn10A D68F/S95A主要生成X2−X4(图7a)。同时,PaXyn10A D68F/S95A水解产生的低聚木糖(XOS)得率是PaXyn10A的2.04倍(图7a)。此前研究发现,X2−X4具有最显著的生物活性,这使得PaXyn10A D68F/S95A成为XOS生产的理想催化剂。
大多数木聚糖酶无法同时具备高热稳定性和耐酸性,这严重限制了其工业应用。平衡稳定性和催化活性仍然是酶工程中的关键挑战。PaXyn10A相对较低的热稳定性限制了其工业应用价值。在本研究中,采用理性设计对PaXyn10A的柔性区域进行改造,以提高其热稳定性、比活力和催化效率,同时保持其耐酸性。
尽管PaXyn10A D68F/S95A的热稳定性(≤50°C)仍低于一些耐热GH10木聚糖酶,如Lc-Xyn81(≤75°C)、XYN329(≤65°C)和AmXyn(≤65°C),但其催化效率(kcat/Km = 103.2 mL/mg/s)高于这些对照酶。Lc-Xyn81、XYN329和AmXyn的kcat/Km值分别为52.48、23.61和91.89 mL/mg/s。值得注意的是,其他几种GH10木聚糖酶相对于PaXyn10A D68F/S95A表现出更优的热稳定性和催化效率。例如,来自Aspergillus fumigatus Af293的Af-XYLA的最适温度为80°C,kcat/Km值为15.456 mL/mg/s。
RMSD分析显示,PaXyn10A在340 K下的结构波动较300 K时更大,表明在高温下热稳定性降低(图1a)。通过RMSF分析进一步表征发现,在340 K下,三个区域(残基55−70、90−95和235−250)的柔性相对于300 K时显著增加(图1b)。此前的研究表明,在这些柔性区域进行靶向突变可有效提高木聚糖酶的热稳定性。通常,较低的ΔΔG值与改善的热稳定性相关。因此,在这些柔性区域内理性设计了20个替换突变,并随后评估了它们对酶活性和热稳定性的影响。
D68F突变体在热稳定性方面显著优于同一位置的其他突变体,这与S95位点的观察结果形成对比。为确定95位点的最优替换,将PaXyn10A与六种耐热GH10木聚糖酶(即AAy、AmXyn、Pg−Xyn、XynRA1、Af−XYLA[29]和AS−xyn10A)进行了多序列比对(图1c)。选择95位点中保守性评分最高(排名第一)的残基作为最优突变靶点(图1d)。在第二轮工程改造中,所得双突变体在热稳定性、催化效率和比酶活力方面相较于野生型酶均有显著提升。
高温下柔性区域的去折叠和不可逆破坏通常是酶热变性主要决定因素。通过理性设计调控N端相互作用以实现刚性化,已被证明是增强结构刚性的有效策略。工程化变体引起了N端区域相互作用网络的重排。值得注意的是,与消失的相互作用相比,该突变体形成了新的氢键重排,且其位置更靠近N端,同时新增了三个疏水相互作用,进一步增强了N端刚性(图4b)。特别值得一提的是,F68与Y323残基之间形成了独特的π−π堆积相互作用,影响尤为显著。已知这种增强的疏水堆积能限制构象熵,同时促进三级结构压缩,这是热稳定性提升的一个明确机制。组合突变对疏水相互作用产生了协同效应(图5c,d)。比较分子动力学分析显示,PaXyn10A D68F/S95A相较于野生型PaXyn10A保持了更优的结构完整性,具体表现为RMSD波动减小、Rg更紧凑以及SASA值降低,表明引入的组合突变有助于形成更紧凑的木聚糖酶结构并增强酶的构象稳定性(图6)。此外,蛋白质表面电荷的改变可影响酶的催化特性。D68F替换中和了局部负电荷微环境(图5a,b),这为PaXyn10A D68F/S95A的最适pH向中性偏移1.0个单位提供了合理解释。
增强柔性区域的刚性可提高热稳定性,但可能降低比酶活力和催化效率。有趣的是,工程化变体PaXyn10A D68F/S95A在热稳定性、比活力和kcat/Km值方面相较于PaXyn10A均有显著提升(图2d、表2)。这种卓越的性能可通过催化装置的结构分析得到合理解释。GH10木聚糖酶通过双置换机制催化水解反应,其活性位点裂缝跨越C端结构域延伸。催化中心通常包含5−6个底物结合亚位点,其中−1和−2亚位点是底物识别和结合的关键锚点,而−1亚位点保持空间约束,排斥被取代的木糖残基,−3亚位点在容纳MeGlcA修饰方面表现出显著的可塑性。在PaXyn10A中,S95的侧链指向−3位点,产生空间位阻,影响了底物与酶的结合(图5e)。引入S95A突变减少了该区域的空间位阻(图5f),有利于底物进入结合沟槽,为观察到的比酶活力提升提供了结构基础。该工程化变体在改善底物可及性与保持催化几何构型方面的结合,为酶工程中经典稳定性-活性权衡问题提供了一个优雅的解决方案。
关键挑战在于优化蛋白质的刚性-柔性平衡,以同时提高热稳定性和催化效率。计算机模拟能够高效快速地识别柔性区域和潜在的功能突变位点。通过在柔性区域引入额外相互作用力并修改蛋白质表面疏水性来策略性增强结构刚性是一种有效策略。然而,过高的结构刚性可能限制活性位点必要的构象动力学,从而对催化功能构成风险。本研究表明,多种蛋白质工程方法的协同整合可显著增强GH10家族木聚糖酶的功能优化。值得注意的是,本方法通过增强N端柔性区域的刚性,同时缓解催化口袋外围的空间位阻,实现了结构刚性与柔性的最佳平衡。
蒸汽爆破是一种有效的生物质预处理技术,通过快速膨胀破坏玉米秸秆的内部结构,以增强木聚糖酶水解效果。温度和保留时间是关键的预处理参数。过高的温度会生成糠醛等副产物,阻碍后续酶水解,在190°C下通过木聚糖酶水解获得的最大XOS得率为48.72%(图7b)。延长保留时间不利于最终产物组成的调控。在最优条件(190°C,25分钟)处理后,玉米秸秆的XOS得率达到64.27%,其中X2和X4组分占总XOS的89.9%(w/w)(图7c)。早期研究主要采用酸/碱处理从玉米秸秆生产XOS。然而,酸碱的使用引发了设备腐蚀和环境安全方面的担忧。随后,Zhang等人采用球磨辅助碱性过氧化氢预处理玉米秸秆,通过纤维素酶-木聚糖酶两步水解工艺获得了69.95%(w/w)的XOS得率。因此,复杂的耦合预处理-酶水解工艺通常被认为是实现高XOS得率的先决条件。在本研究中,通过简单的“蒸汽爆破+一步木聚糖酶水解”工艺获得了相当的XOS得率,结合PaXyn10A成功的理性设计,最终实现了从玉米秸秆高效便捷生产XOS的技术目标。
总之,本研究采用分子动力学模拟、虚拟饱和突变、多序列比对和保守序列分析,靶向GH10木聚糖酶的三个柔性区域。成功构建的双突变体PaXyn10A D68F/S95A在热稳定性和催化性能方面表现出协同提升。热稳定性的增强可部分归因于优化的分子间相互作用赋予了更紧凑的结构。同时,PaXyn10A D68F/S95A催化效率的提高源于双重优化机制:通过缓解活性位点微环境中的空间约束来简化底物容纳,以及有利的疏水微环境重塑。这一理性再设计的生物催化剂展现出优越的工业应用性。评估了该突变体在玉米秸秆生产XOS中的应用潜力,通过PaXyn10A D68F/S95A与蒸汽爆破相结合,获得了64.27%的最高XOS得率。








 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
