PBJ|安徽农业大学揭示串联MADS-box基因FUL2与MADS1形成调控模块,通过直接激活ASMT5抑制番茄果实中的血清素生物合成
尽管血清素在植物防御和人类营养方面具有潜在作用,但其在果实发育和成熟过程中的代谢调控仍知之甚少。本文证明,MADS-box转录因子FUL2通过与MADS1形成功能模块,成为番茄中血清素积累的关键抑制因子。CRISPR-Cas9产生的ful2-cr突变体表现出成熟延迟、果实尺寸减小以及血清素水平惊人的10倍增加,这揭示了FUL2与次级代谢之间先前未被认识的联系。免疫沉淀-质谱分析揭示FUL2与MADS1存在物理相互作用,遗传分析显示mads1-cr突变体在发育和血清素超积累表型上都复制了ful2-cr突变体的表型。此外,ChIP-seq和转录组分析证明,FUL2-MADS1复合体直接结合ASMT5启动子中的CATG-box基序,激活其表达,同时抑制TDC1。电泳迁移率变动实验和双荧光素酶报告基因实验证实了它们的协同DNA结合和协同转录调控。我们的工作建立了一个MADS-box转录模块,通过协调调控生物合成和代谢基因来控制血清素通量。这些发现为工程化改造作物中的血清素含量提供了框架,并加深了对发育转录因子如何在成熟过程中控制特化代谢的理解。图 1:FUL2功能丧失突变体改变果实成熟并增强血清素生物合成
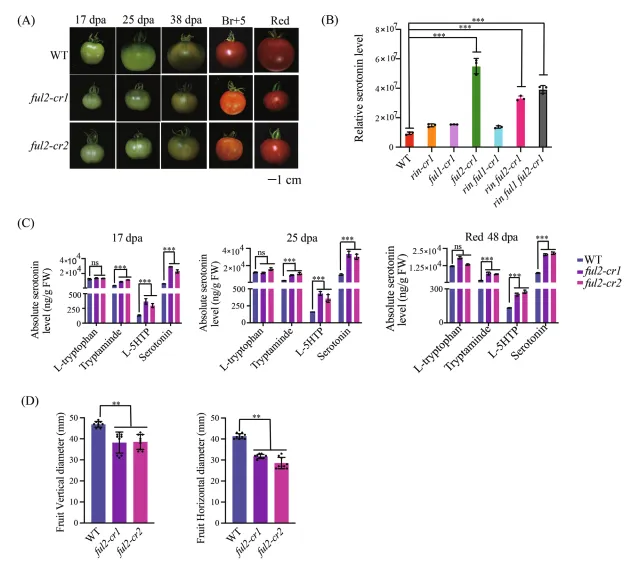
(A) 在破色期后5天,野生型和两个独立ful2-cr突变体(ful2-cr1和ful2-cr2)果实的表型。ful2-cr突变体果实呈现独特的橙色着色。比例尺=1厘米。
(B) 通过广靶向代谢组学检测的破色期果实血清素含量。数据以平均±标准差表示(n=3)。星号表示与野生型相比的显著性差异(学生t检验:**p<0.01)。
(C) 在多个基因型中通过靶向代谢组学验证的血清素含量。数据以平均±标准差表示(n=3)。不同字母表示显著性差异(Tukey检验,p<0.05)。
(D) 成熟过程中野生型和ful2-cr1突变体果实的代表性图像。图 2:FUL2相互作用调节因子在血清素代谢中的功能表征
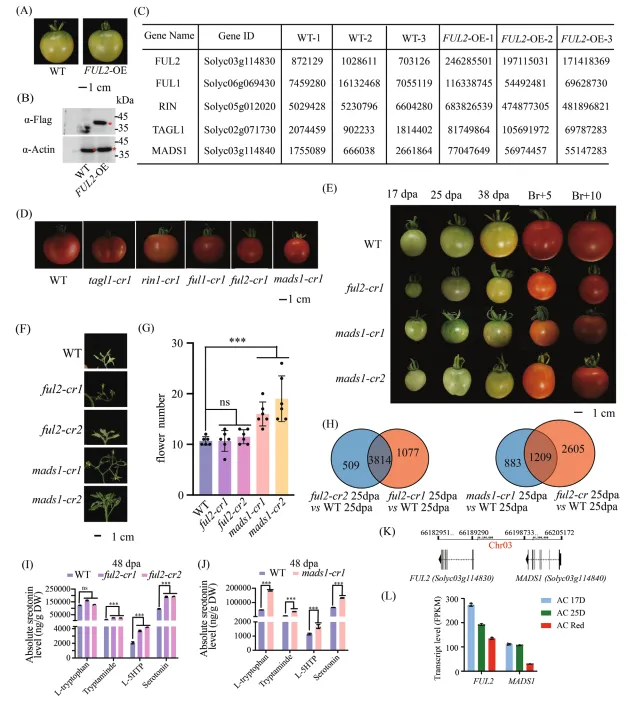
(A) 破色期野生型和FUL2过表达(FUL2-OE)果实的表型比较。比例尺=1厘米。
(B) 使用抗FLAG抗体对破色期FUL2-OE和野生型果实中FUL2蛋白水平的免疫印迹分析,以Actin作为上样对照。星号表示FUL2-FLAG融合蛋白。
(C) 通过对破色期果实的FUL2免疫沉淀物进行质谱分析,鉴定出的主要FUL2相互作用候选蛋白(n=3个生物学重复)。
(D) 在破色期后10天,tagl1-cr1、rin-cr1、ful1-cr1、ful2-cr1、mads1-cr1突变体与野生型果实的表型比较。比例尺=1厘米。
(E) ful2-cr1、mads1-cr1、mads1-cr2突变体和野生型果实在成熟阶段过程中的发育进程。比例尺=1厘米。
(F, G) 花序结构分析:(F) 花表型的代表性图像;(G) 每个花序花数的定量评估(平均±标准差,n=6)。
(H) 差异表达基因的维恩图:(左)花后25天ful2-cr1 vs. 野生型和ful2-cr2 vs. 野生型之间差异表达基因的重叠;(右)花后25天mads1-cr1 vs. 野生型和ful2-cr vs. 野生型之间差异表达基因的重叠。
(I, J) 在花后48天,(I) ful2突变体和(J) mads1突变体与野生型相比,色氨酸途径中间产物的绝对定量(平均±标准差,n=3)。统计学显著性通过单因素方差分析(Tukey检验)或学生t检验确定:***p<0.001;ns,不显著。图 3:FUL2与MADS1在血清素调控中的分子相互作用
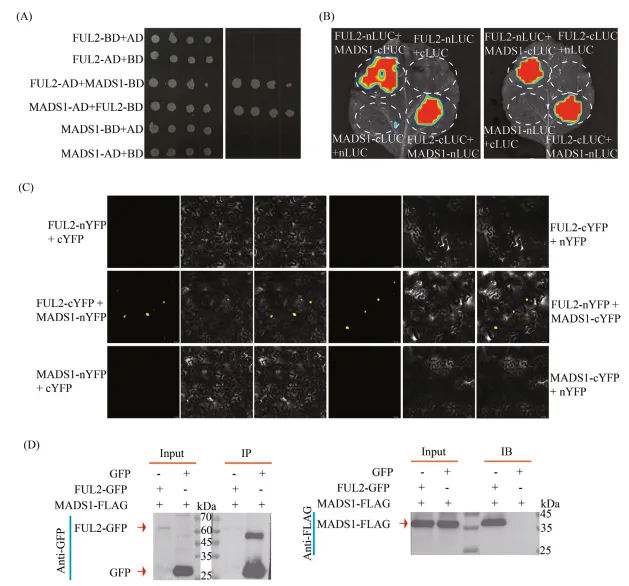
(A) 酵母双杂交实验证明FUL2与MADS1的直接相互作用。
(B) 在本氏烟草叶片中进行的Split-luciferase互补实验。
(C) 在本氏烟草表皮细胞中的双分子荧光互补可视化。
(D) 在本氏烟草中间时表达FUL2-GFP和MADS1-FLAG。使用GFP-Trap磁珠对FUL2-GFP和GFP对照进行免疫沉淀,并使用抗FLAG抗体通过免疫印迹检测共沉淀的MADS1-FLAG。此结果证实了FUL2与MADS1在番茄中的特异性相互作用。图 4:FUL2和MADS1对血清素生物合成中转录网络的协同调控
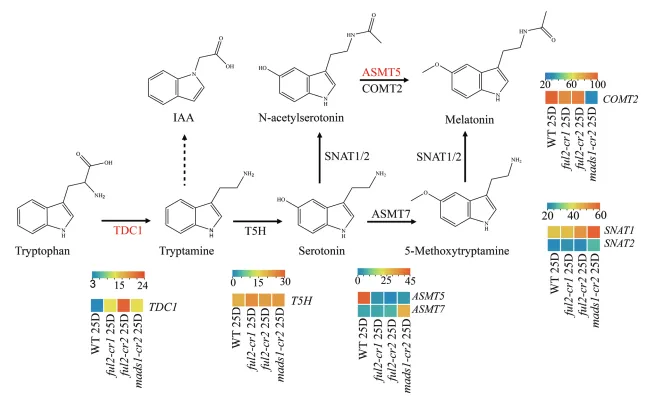
在番茄中,从色氨酸到血清素及其衍生物褪黑素的生物合成途径示意图。FUL2和MADS1通过调控关键酶基因(TDC1、TSH、SNAT、ASMT5、COMT)来协调血清素的积累和向褪黑素的转化。色块表示基因表达水平(绿色:表达较低;红色:表达较高)。连接线代表调控关系(箭头:激活;T型线:抑制)。图 5:血清素生物合成中FUL2靶基因的全基因组鉴定
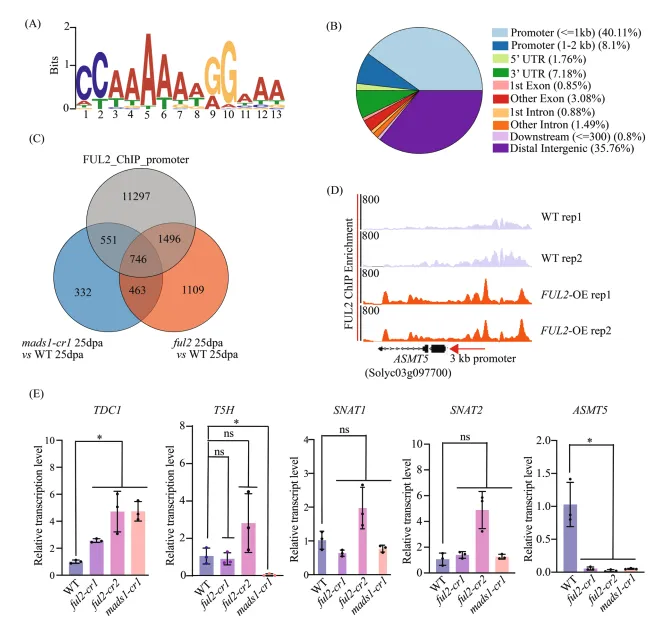
(A) FUL2 ChIP-seq峰值的从头基序分析揭示了富含的CATG-box共有序列。
(B) FUL2结合位点的基因组分布。饼图百分比表示在启动子、基因区和基因间区的峰值。
(C) FUL2靶基因的整合分析:维恩图显示FUL2结合(ChIP-seq)与ful2-cr和mads1-cr1突变体中差异表达基因之间的重叠。
(D) FUL2在ASMT5启动子区域占据情况的代表性ChIP-seq轨迹图。y轴表示归一化的读长覆盖度。
(E) 在花后25天,通过定量RT-PCR验证ful2-cr1、ful2-cr2和mads1-cr1突变体与野生型相比血清素途径基因的表达。数据以平均±标准差表示(n=3个生物学重复)。统计学显著性通过单因素方差分析(Tukey检验)确定:*p<0.05;ns,不显著。图 6:FUL2-MADS1复合体对ASMT5的直接结合和转录激活
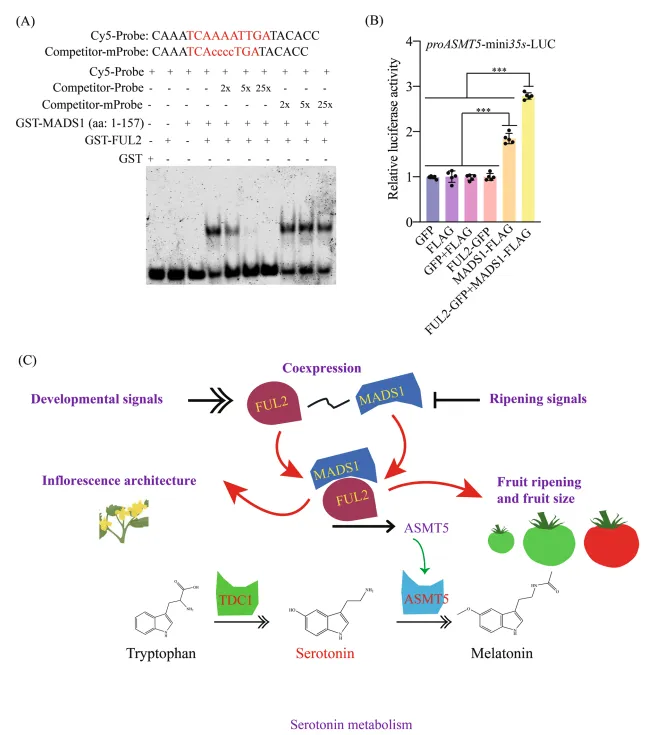
(A) 电泳迁移率变动实验证明重组FUL2-MADS1复合体与标记的ASMT5启动子中CATG-box基序的序列特异性结合。使用2倍、5倍和25倍摩尔过量的未标记探针进行的竞争实验显示出剂量依赖性的结合减弱。
(B) 在本氏烟草叶片中进行的双荧光素酶报告基因实验显示,共表达FUL2和MADS1对ASMT5启动子具有协同激活作用。数据以平均±标准差表示(n=5)。星号表示显著性差异(单因素方差分析,Tukey检验,***p<0.001)。
(C) 简化模型示意图。番茄果实发育和成熟受复杂网络调控,转录因子FUL2和MADS1在其中起关键作用。FUL2和MADS1形成一个功能模块,调控ASMT5和TDC1的表达,从而控制血清素的通量。研究发现,FUL2和MADS1的突变体表现出成熟延迟、果实尺寸减小以及血清素水平显著增加。这些发现不仅凸显了FUL2和MADS1在果实发育和代谢中的作用,也为工程化改造作物中的血清素含量提供了框架,这对植物防御机制和人类营养具有重要意义。 聚焦植物科学前沿动态,追踪领域最新研究进展。从基础机制到应用突破,从分子层面到生态系统,这里有权威解读、成果速递与趋势洞察,陪你一起探索植物世界的奥秘与创新——关注「植物科学研究进展」,读懂植物科学的每一步成长。

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
