PP (IF: 8.2) | 中国农业大学等: 番茄SlNAC3调控蛋白质稳态以响应早期低温胁迫
- 2026-07-17 05:47:52
PP (IF: 8.2) | 中国农业大学等: 番茄SlNAC3调控蛋白质稳态以响应早期低温胁迫
低温胁迫对番茄生产构成重大挑战。SlNAC3是番茄中的一个NAC转录因子,在低温胁迫的早期响应中发挥着关键作用。然而,此前仅有少数参与乙烯生物合成的基因被鉴定为负调控耐冷性的NAC3靶基因。本研究探究了SlNAC3影响耐冷性的机制。对经过2小时低温胁迫处理的SlNAC3瞬时沉默株系和过表达株系进行RNA测序分析,结果显示SlNAC3下调了核糖体途径,同时上调了泛素-蛋白酶体系统。进一步分析表明,SlNAC3直接靶向参与泛素-蛋白酶体系统的基因,导致泛素化蛋白和游离氨基酸的积累。此外,SlNAC3负调控核糖体蛋白的表达,减少了可溶性蛋白的积累,并损害了低温胁迫下CBF蛋白的水平。我们的研究结果表明,SlNAC3的早期诱导可能激活CBF的转录,而SlNAC3持续的高水平表达会激活泛素-蛋白酶体系统,从而损害CBF蛋白的积累和番茄的耐冷性。 篇名: Tomato SlNAC3 regulates proteostasis in response to early cold stress 期刊: Plant Physiology 影响因子: 8.2(1区top) 发表时间: 2026.7 通讯单位: 中国农业大学、海南省农业科学院、斯特拉斯堡大学 DOI: 10.1093/plphys/kiag501 
(1) SlNAC3的瞬时沉默导致耐冷性增强 番茄NAC3在低温胁迫的早期响应中发挥着关键作用,然而此前仅有少数参与乙烯生物合成的基因被鉴定为负调控耐冷性的NAC3靶基因(Wang等,2024)。为了全面理解SlNAC3在早期低温胁迫响应中所诱导的转录组重编程,我们在2小时低温胁迫后,对三个不同的株系:野生型、SlNAC3沉默株系和SlNAC3过表达株系(OE-SlNAC3 #5)进行了RNA测序分析(表S2)。 SlNAC3的组成型缺失在非胁迫条件下也会导致明显的发育表型,表明该基因具有强烈的多效性效应。这些多效性效应使低温胁迫下下游转录响应的解释变得复杂,并可能掩盖早期、直接的调控事件(Mohanta等,2020;Xiong等,2025)。重要的是,SlNAC3是一个在低温处理2小时后快速诱导的转录因子,表明它主要在低温胁迫的极早期阶段发挥作用(Olsen等,2005;Nakashima等,2012;Yuan等,2022;Wang等,2024)。因此,为了捕捉清晰且即时的SlNAC3依赖性转录响应,同时最大限度地减少次级效应或补偿效应,我们采用了瞬时沉默株系。与SlNAC3敲除株系相似(Wang等,2024),瞬时沉默SlNAC3导致耐冷性增强,这通过表型、相对电解质渗透率、丙二醛含量和光合效率得到了证实(图1A-E)。因此,该沉默株系被用于RNA测序分析。在正常条件下,主成分分析显示对照株系和SlNAC3沉默株系之间具有相对相似的转录组谱。然而,随后进行的2小时低温处理凸显了对照株系和TRV:SlNAC3株系之间的差异(图1F)。这一结果有力地表明SlNAC3在早期低温响应中具有关键作用,这与经过2小时低温处理后,其转录本水平相比于其他NAC转录因子显著升高的情况一致(Wang等,2024)。此外,三个TRV:SlNAC3株系中有一个在低温胁迫下表现出与对照相似的特征,表明瞬时沉默效应的影响较小。 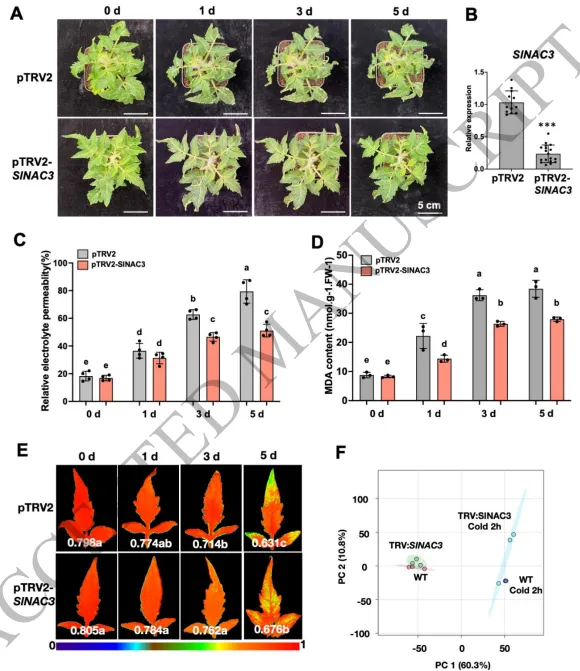
图1. 病毒诱导的SlNAC3基因沉默增强了番茄植株的耐寒性。(A) pTRV2和pTRV-SlNAC3番茄植株在低温胁迫(4°C)处理5天后的表型。比例尺 = 5厘米。处理中使用的是Micro-Tom品种。(B) pTRV诱导的SlNAC3沉默株系和pTRV2对照中SlNAC3的相对表达水平。图中分析了显著差异性(n=12-18,平均值±标准差,单因素方差分析后采用Tukey事后检验,*** p < 0.001)。(C-D) Micro-Tom品种的pTRV2和pTRV-SlNAC3番茄植株在不同低温胁迫(4°C)时间下的相对电解质渗透率(REP)(C)、丙二醛(MDA)含量(D)和Fv/Fm(光系统II的最大量子产额)(E)。图上的小写字母或星号表示显著差异(n=4,平均值±标准差,单因素方差分析后采用Tukey事后检验,p < 0.05)。(E) pTRV2和pTRV-SlNAC3植株在不同低温胁迫(4°C)时间下的Fv/Fm(光系统II的最大量子产额)。图像底部描绘的颜色变化条代表叶片的损伤程度。小写字母表示叶片损伤更严重。(F) 对Micro-Tom品种的pTRV2和pTRV-SlNAC3番茄植株在2小时低温胁迫(4°C)后的RNA-seq数据进行的主成分分析(PCA)。 热图分析揭示了一个引人注目的基因簇,命名为“Group 1”(图S1A)。该基因集合包含了一系列在野生型株系中响应低温的基因。这些基因在常温条件下的SlNAC3沉默株系中表现出轻微诱导,而在低温胁迫处理后表现出更为显著的诱导。Group 1基因的功能特性很可能对在SlNAC3沉默株系中观察到的耐冷性增强表型有显著贡献。因此,我们对该基因集中富集的KEGG通路进行了细致的分析(图S1B)。分析揭示了“植物-病原体互作”通路的优势富集,该通路包含多种与钙信号通路有关的基因,包括钙依赖性蛋白激酶和钙结合蛋白(图S1C)。此外,在鉴定出的多种通路中,参与活性氧稳态的谷胱甘肽还原酶,以及与细胞壁修饰和可溶性糖积累相关的β-半乳糖苷酶(TBG13)反复出现(图S1D,E)。所有上述通路已被证实是耐冷性的正向调节因子。 (2) SlNAC3对冷诱导的转录组重编程表现出微调效应 除了分析SlNAC3瞬时沉默株系,我们还对先前构建的SlNAC3过表达株系(OE-SlNAC3 #5)在2小时低温胁迫后进行了RNA测序。在所有三个受检株系中,响应低温胁迫而差异表达的基因主要表现出上调模式(图2A),且富集的功能类别相似(图S2)。有趣的是,包含上调基因和下调基因的三个不同基因集显示出显著的重叠(图2B-E)。这一观察结果表明,SlNAC3在早期低温胁迫响应的转录组重编程过程中可能具有有限的意义。此外,核糖体基因仅在低温胁迫下的TRV:SlNAC3株系的上调基因中富集(图S2),这暗示SlNAC3在核糖体生物发生和翻译中具有负向作用。在经历低温处理的OE-SlNAC3株系中,观察到的上调基因数量显著减少。这一现象很可能归因于SlNAC3过表达所诱导的预处理效应。推测该基因簇在预设生理状态中发挥作用,而这种生理状态最终导致了OE-SlNAC3株系表现出的冷敏感表型。 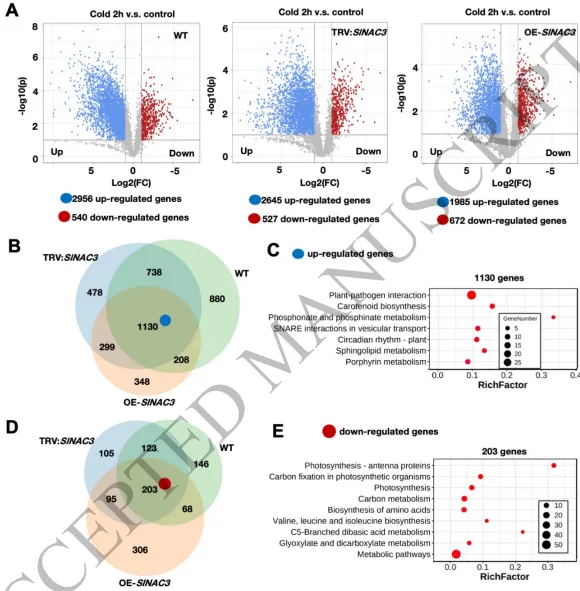
图2. 对照及2小时低温胁迫条件下野生型、pTRV-SlNAC3和OE-SlNAC3株系的转录组学分析。(A) 在对照和2小时低温胁迫(4°C)条件下,野生型、pTRV-SlNAC3和OE-SlNAC3株系(OE#5)中的差异基因火山图。横坐标表示基因表达量的倍数变化,纵坐标表示差异基因的显著性水平。红点代表上调差异基因,蓝点代表下调差异基因,灰点代表非差异表达基因。韦恩图展示了不同的基因。(B, D) 韦恩图展示了在对照和2小时低温胁迫(4°C)条件下,野生型、pTRV-SlNAC3和OE-SlNAC3株系(OE#5)之间上调(B)和下调(D)差异表达基因(DEG)的比较。(C, E) KEGG富集分析展示了基于上调(C)和下调(E)差异表达基因数量富集的通路,按富集因子排序。 通过比较野生型和OE-SlNAC3(OE#5)株系,主成分分析清晰地将四个样本区分开来。具体而言,无论低温处理状态如何,PC2都呈现出基于基因型的明显聚类(图3A)。重点关注PC2的正载荷值,这些值代表因SlNAC3过表达而被抑制的基因,我们选取了贡献最大的前500个基因。该基因集在碳/硫代谢和核糖体过程中表现出显著富集(图3B)。对与PC2负载荷值相关的、代表被SlNAC3过表达诱导的前500个基因进行更细致的分析,揭示了三个不同通路的显著覆盖:蛋白质加工、蛋白质降解和蛋白质输出(图3C)。 为了解析驱动OE-SlNAC3株系冷敏感表型的潜在机制,我们对这些株系在低温胁迫下异常表达的基因进行了细致的探究。通过热图聚类分析,鉴定出两个不同的基因群组(图3D-H)。Group 1基因包含那些在野生型中由低温处理诱导,但相对于两个SlNAC3过表达株系的平均表达水平却受到抑制的基因。相反,Group 2基因则由那些在野生型中受低温处理抑制,但在OE-SlNAC3株系中却异常诱导的基因组成。对在野生型中低温诱导但被SlNAC3过表达抑制的Group 1基因的进一步研究,揭示了其在碳和硫代谢途径以及核糖体过程中的富集(图3E)。诸如大亚基核糖体蛋白1(RPL1)、亚硫酸盐还原酶(SiR)和乙烯响应因子1(ERF1)等显著例子在OE-SlNAC3株系中受到显著抑制(图3G)。相反,Group 2基因代表了在野生型中受低温处理抑制,但在OE-SlNAC3株系中异常诱导的基因,这些基因主要与蛋白质加工途径相关(图3F)。该基因子集包括热休克70kDa蛋白1(HSPA1)和微管蛋白β(TUBB)等元件,这些基因在OE-SlNAC3株系中表现出异常诱导(图3H)。 
图3. 对照及2小时低温胁迫条件下野生型、pTRV-SlNAC3和OE-SlNAC3株系的转录组学分析。(A) 在对照和2小时低温胁迫(4°C)条件下,野生型和OE-SlNAC3株系(OE#5)之间转录组谱的主成分分析(PCA)。(B, C) 对PC2水平上正载荷(B)和负载荷(C)的差异表达基因进行KEGG富集分析。(D) 在对照和2小时低温胁迫(4°C)条件下,野生型和OE-SlNAC3#5株系的热图分析。(E, F) 对(D)中的组1 (E) 基因和组2 (F) 基因进行KEGG富集分析。(G, H) 代表性基因的相对转录水平,包括在OE-SlNAC3株系(OE#4, OE#5)中被抑制的组1基因(G),以及在野生型中被低温处理抑制但在OE-SlNAC3株系(OE#4, OE#5)中被诱导的组2基因(H)。EF-1α用作内参对照。数据显示为三个独立生物学重复的平均值,每个重复包含5-6株单株。 (3) SlNAC3协调蛋白质稳态和氨基酸代谢重编程以适应胁迫 鉴于观察到的核糖体途径下调和蛋白水解途径的同步上调,我们有理由推测OE-SlNAC3株系可能调控蛋白质稳态。我们对泛素介导的蛋白水解途径中的关键基因进行了仔细分析,包括F-box蛋白S期激酶相关蛋白2(SKP2)、泛素激活酶E1(UBE1)、泛素C(UBC)以及泛素结合酶E2 D/J2(UBE2D/J2)。这些基因的启动子中含有多个NAC结合域,并在OE-SlNAC3株系中表现出上调,而在敲除突变株系(CR#2和CR#13)中表现出下调(图4A,B;图S3C)。此外,低温胁迫抵消了NAC3靶向的参与蛋白水解的基因的表达(图4A,B)。由于存在多个NAC结合域,我们采用双荧光素酶实验来研究SlNAC3对这些基因的调控(图5A,B)。事实上,SlNAC3与由靶基因启动子驱动的LUC的共浸润实验表明,SlNAC3激活了UBC、UBE和UBE2J2的转录(图5C),这表明SlNAC3广泛地调控泛素-蛋白酶体系统。此外,ChIP-qPCR分析表明,SlNAC3在体内结合到UBC、UBE和UBE2J2的启动子上(图5D;图S3D)。 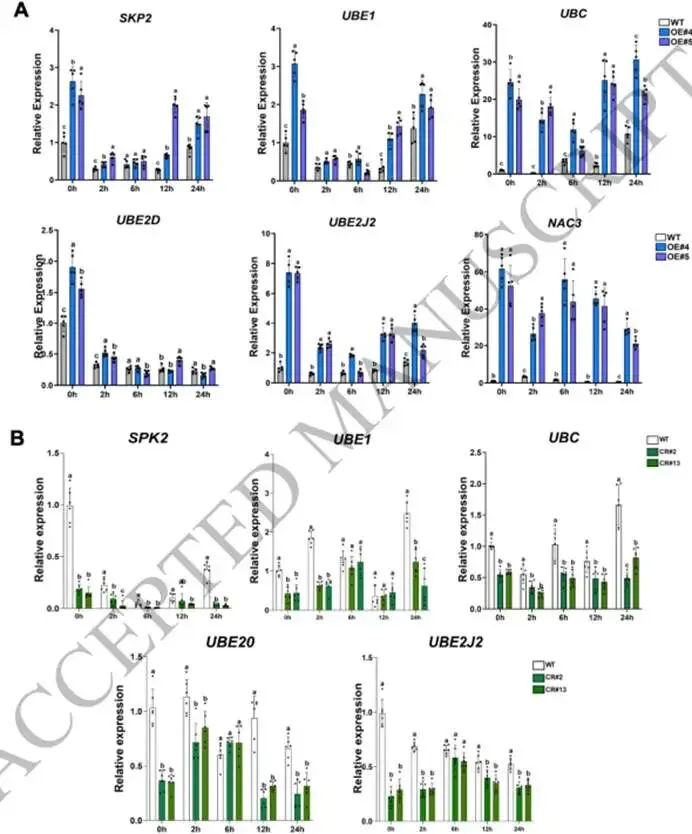
图4. SlNAC3调控参与蛋白水解过程的基因的转录。(A) 在不同低温胁迫时间下,Micro-Tom野生型和OE-SlNAC3株系(OE#4, OE#5)中SKP2、UBE1、UBC、UBE2J2和UBE2D的相对表达水平。EF-1α用作内参对照。(B) 在不同低温胁迫时间下,Micro-Tom野生型和CRISPR/Cas9敲除SlNAC3株系(CR#2, CR#13)(Wang et al., 2024)中SKP2、UBE1、UBC、UBE2J2和UBE2D的相对表达水平。EF-1α用作内参对照。数据显示为五个独立生物学重复的平均值,每个重复包含3-5株单株。图上的小写字母或星号表示显著差异(平均值±标准差,单因素方差分析后采用Duncan事后检验,p < 0.05)。 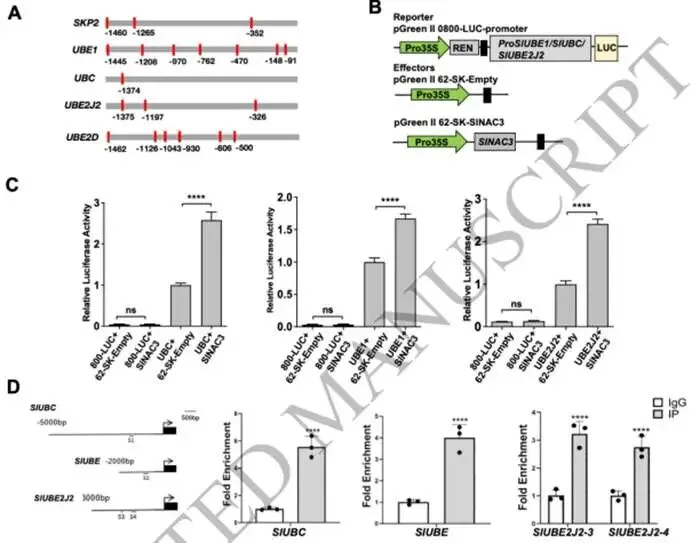
图5. SlNAC3结合到蛋白水解相关基因的启动子上。(A) SKP2、UBE1、UBC、UBE2J2和UBE2D启动子上游约1.5 kb序列内NAC结合序列(CACG 或 CGT(A/G))位置的示意图。数字代表距翻译起始位点的距离。CACG或CGT(A/G)的位置在启动子示意图上用红色条标记。(B) 双荧光素酶检测实验中报告基因和效应基因构建的示意图。(C) UBE1、UBC和UBE2J2各组的双荧光素酶检测实验的相对LUC/REN(萤火虫荧光素酶/海肾荧光素酶)比值。数据显示为三个独立生物学重复的平均值,每个重复包含3-5株单株。星号表示显著差异(单因素方差分析后采用Tukey事后检验,****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.05)。(D) 针对UBE1、UBC和UBE2J2启动子指定片段(S1-P4)的ChIP-qPCR分析。来自p35S:SlNAC3-GFP转基因OE-SlNAC3株系(OE#4, OE#5)的染色质用抗GFP抗体进行免疫沉淀。Micro-Tom野生型植株用作阴性对照。通过qPCR测定指定DNA片段的量,并归一化至野生型对照。数据显示为三个独立生物学重复的平均值,每个重复包含3-5株单株。 研究发现五个SlNAC3靶基因(SKP2、UBE1、UBC、UBE2J2和UBE2D)在整个24小时低温处理期间表现出显著的时序表达动态。然而,在大多数时间点,它们在OE-SlNAC3株系中的转录本水平显著高于野生型,表明在低温胁迫期间,SlNAC3的表达与这些泛素化相关基因的激活之间存在正相关关系(图4A,B)。基于这一观察,我们随后检测了低温胁迫下OE-SlNAC3株系中泛素化蛋白的水平(图6A,B;图S3A)。在野生型植株中,泛素化蛋白水平在低温处理2小时和6小时后逐渐降低,而在OE-SlNAC3株系中观察到相反的趋势,即泛素化蛋白在同一时期内积累(图6A,B)。相比之下,SlNAC3敲除株系中的泛素化水平在低温胁迫6小时后急剧下降(图6C,D)。这些结果表明,SlNAC3在低温胁迫的早期阶段促进持续的泛素介导的蛋白水解。 为了描绘由增强的蛋白水解作用释放的氨基酸的命运,我们量化了氮同化和次生代谢中的中间产物。在OE-SlNAC3株系中,参与谷氨酸代谢的酶,如谷氨酸脱氢酶(GDH)和谷氨酰胺合成酶(GS),表现出表达改变(转录组数据,未显示),这与谷氨酸和谷氨酰胺的显著积累相关(图6E-G)。此外,作为胁迫相关代谢物前体的氨基酸,包括脯氨酸(Pro)和γ-氨基丁酸(GABA),在OE-SlNAC3株系中升高(图6F,G)。脯氨酸是一种关键的渗透调节物质,GABA是胁迫响应中的信号分子,这提示SlNAC3介导的蛋白水解不仅支持氮的再循环,也支持胁迫适应。这表明蛋白水解产生的氨基酸碳骨架进入了三羧酸循环,证据是OE-SlNAC3株系中三羧酸循环中间产物(如α-酮戊二酸和草酰乙酸)的水平增加,从而将增强的蛋白水解与中心碳代谢联系起来(Fuentes等,2001;Nagaya等,2005;Roberts等,2014;Beernink等,2025)。 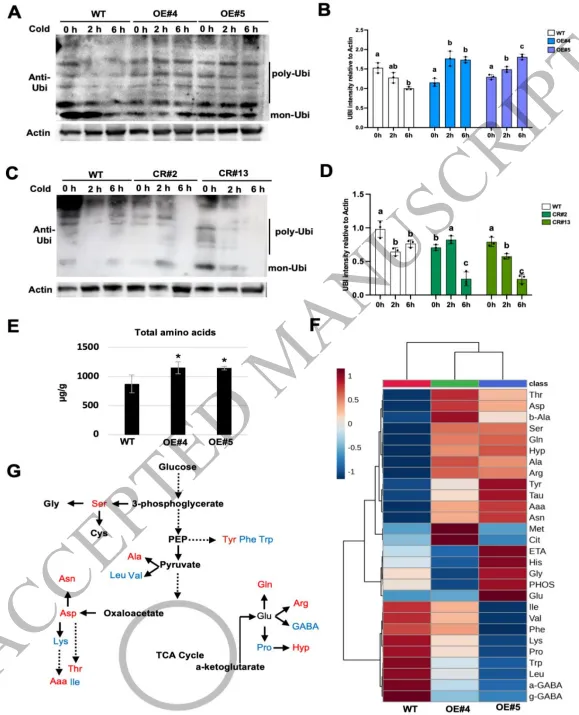
图6. OE-SlNAC3株系中的总蛋白泛素化分析。(A, C) 在不同低温胁迫时间下,野生型和OE-SlNAC3株系(OE#4, OE#5)(A),以及CRISPR/Cas9敲除株系(CR#2和CR#13)(C)中泛素蛋白的蛋白质印迹分析。(B, D) 通过Image J软件测定的相对于肌动蛋白的泛素蛋白信号强度。数据显示为三个独立生物学重复的平均值,每个重复包含5-6株单株。图上的小写字母或星号表示显著差异(n=4,平均值±标准差,单因素方差分析后采用Duncan事后检验,p < 0.05)。(E-G) Micro-Tom野生型和SlNAC3过表达株系(OE#4, OE#5)中的氨基酸谱。总氨基酸水平显示于(E)。单个氨基酸的变化以热图形式展示(F)。两个过表达株系(OE#4, OE#5)中一致的变化在氨基酸代谢通路图中描绘,红色表示上调,蓝色表示下调(G)。数据来源于三个独立的生物学重复,每个重复包含5-6株单株。星号表示显著差异(单因素方差分析后采用Tukey事后检验,***p < 0.001, **p < 0.01, *p < 0.05)。 鉴于核糖体途径的下调(图2B;图3E-H),我们研究了减少的蛋白质合成与增强的蛋白水解之间的协调性。总蛋白泛素化分析显示,在OE-SlNAC3株系中,单泛素化蛋白的比例较低,而多泛素化蛋白的比例较高,这与翻译的减弱相一致(图5A-D)。翻译和蛋白水解之间的平衡可能确保了高效的氮再利用,即过剩或受损的蛋白质通过泛素-蛋白酶体系统(UPS)降解,释放的氨基酸被重新导向至响应胁迫的代谢。此外,在OE-SlNAC3株系中,负责陪伴错误折叠蛋白的热休克蛋白(HSP)的转录本水平降低(图3H),这暗示SlNAC3将平衡从分子伴侣介导的重折叠转向蛋白酶体降解,进一步强化了蛋白质稳态的重塑。 (4) SlNAC3过表达在早期低温胁迫期间减弱核糖体基因表达并损害CBF蛋白的翻译 为了进一步阐明SlNAC3在低温胁迫响应中的调控机制,我们探索了核糖体功能和蛋白质动态分析的更多层面。关键核糖体蛋白,例如大亚基核糖体蛋白L10/13/18/21(RPL10/13/18/21)(图7A)和小亚基核糖体蛋白S6(RPS6)(图S3B),在OE-SlNAC3株系中表现出一致的下调,在KO株系中表现出上调。核糖体基因表达的广泛抑制意味着翻译机器的整体减弱。可溶性蛋白水平的明显降低是植物响应低温胁迫的一个关键特征,并最终影响其冷敏感性(Dong等,2006;Ding等,2018;Chen等,2023)。实际上,当我们使用考马斯亮蓝染色测定相同量新鲜组织中的总可溶性蛋白时,野生型在低温处理后表现出蛋白质含量的逐渐增加,而这种增加在OE-SlNAC3株系中减弱了(图7B)。在敲除株系中,蛋白质积累未受影响(图7C)。尽管SlNAC3的敲除导致了核糖体基因的上调,但蛋白质合成是一个耗能过程,并受到严格的细胞调控。因此,单一敲除SlNAC3不足以改变整体的蛋白质稳态,这可能是由于维持翻译平衡的补偿机制。相反,SlNAC3的过表达直接抑制核糖体基因表达并促进蛋白质周转,导致在低温胁迫下总可溶性蛋白出现可测量的减少。这些观察结果强调了功能获得与功能缺失研究方法的独特逻辑,即过表达放大了调控效应,而单一敲除则可以被冗余途径和能量限制所缓冲。 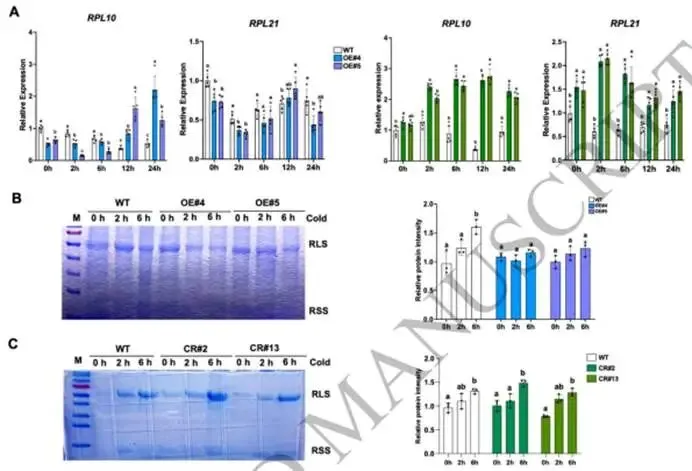
图7. SlNAC3在响应早期低温胁迫中调控蛋白质合成效率。(A) 在不同低温胁迫时间下,Micro-Tom野生型和SlNAC3过表达株系(OE#4, OE#5),以及野生型和敲除株系(CR#2和CR#13)中RPL10、RPL21的相对表达水平。EF-1α用作内参对照。数据显示为五个独立生物学重复的平均值,每个重复包含5-6株单株。不同字母表示显著差异(单因素方差分析后采用Duncan事后检验,p < 0.05)。(B) 在2小时和6小时低温胁迫下,通过考马斯亮蓝染色测定Micro-Tom野生型和SlNAC3过表达株系(OE#4, OE#5)的总蛋白水平(200毫克材料/200微升裂解液)。通过Image J软件分析在2小时和6小时低温胁迫下,Micro-Tom野生型和SlNAC3过表达株系(OE#4, OE#5)相对于野生型0小时的总蛋白水平信号强度。数据来源于三个重复,每个重复包含5-6株单株。(C) 在2小时和6小时低温胁迫下,通过考马斯亮蓝染色测定Micro-Tom野生型和敲除株系(CR#2和CR#13)的总蛋白水平(200毫克材料/200微升裂解液)。通过Image J软件分析在2小时和6小时低温胁迫下,Micro-Tom野生型和敲除株系(CR#2和CR#13)相对于野生型0小时的总蛋白水平信号强度。数据来源于三个重复,每个重复包含5-6株单株。不同字母表示显著差异(单因素方差分析后采用Duncan事后检验,p < 0.05)。 我们进一步检测了低温暴露下OE-SlNAC3株系和KO株系中CBF1/2/3的转录本和蛋白质水平。在OE-SlNAC3植株中,CBF蛋白的积累在低温处理期间发生了显著改变。尽管低温暴露诱导了野生型植株中CBF蛋白的积累,但在整个处理期间,所有三种CBF蛋白的水平在OE株系中始终维持在显著较低的水平(图8A-B)。为了确定这些蛋白质变化是否与转录改变相关,我们分析了CBF基因的转录本水平。一致地,OE-SlNAC3株系中的CBF转录本在0小时已经升高,并在低温处理后表现出动态变化(图8C),表明SlNAC3正调控CBF的转录。值得注意的是,尽管OE株系中CBF1转录本水平持续较高,但其相应的蛋白却未能积累。同样,尽管低温处理6小时后,CBF2/3转录本水平下降至野生型水平,但其蛋白水平在6小时时与野生型相比仍然受到强烈抑制(图8A-C)。相比之下,在KO株系中,CBF蛋白的积累与野生型相当,尽管在每个时间点其转录本和蛋白水平普遍低于野生型(图8D-F)。 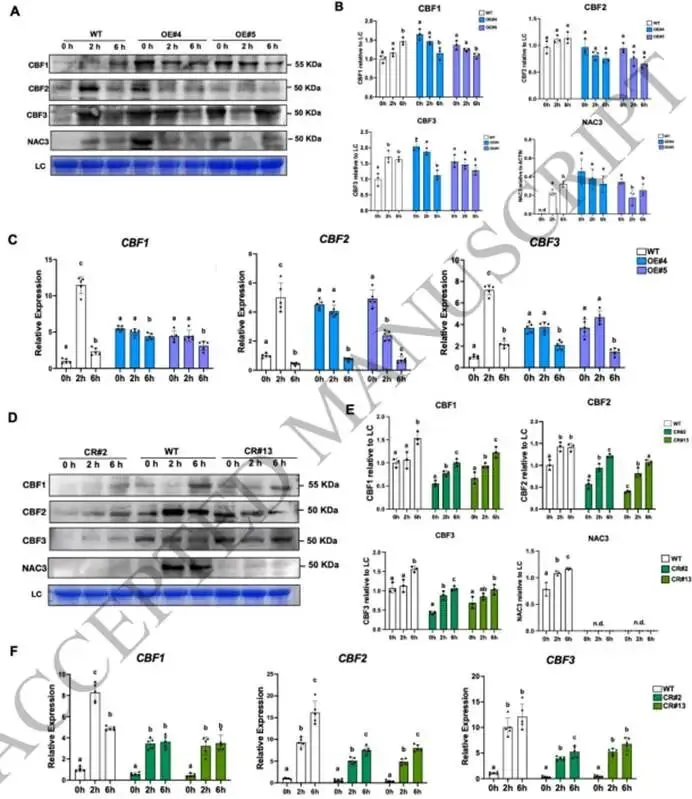
图8. SlNAC3过表达削弱了早期低温胁迫下CBF蛋白的翻译。(A) 在不同低温胁迫时间下,野生型和OE-SlNAC3株系(OE#4, OE#5)中CBF1/2/3蛋白、NAC3、肌动蛋白的蛋白质印迹分析以及考马斯亮蓝G-250染色的LC(大亚基)。(B) 通过Image J软件分析在2小时和6小时低温胁迫下,Micro-Tom野生型和OE-SlNAC3株系(OE#4, OE#5)中CBF1/2/3、SlNAC3蛋白相对于LC的信号强度。数据来源于三个重复,每个重复包含5-6株单株。不同字母表示显著差异(单因素方差分析后采用Duncan事后检验,p < 0.05)。(C) 在不同低温胁迫时间下,Micro-Tom野生型和SlNAC3过表达株系(OE#4, OE#5)中CBF1/2/3和SlNAC3的相对转录水平。EF-1α用作内参对照。数据显示为五个独立生物学重复的平均值,每个重复包含5-6株单株。(D) 在不同低温胁迫时间下,野生型和敲除株系(CR#2和CR#13)中CBF1/2/3蛋白、NAC3、肌动蛋白的蛋白质印迹分析以及考马斯亮蓝G-250染色的LC(大亚基)。(E) 通过Image J软件分析在2小时和6小时低温胁迫下,Micro-Tom野生型和敲除株系(CR#2和CR#13)中CBF1/2/3、SlNAC3蛋白相对于LC的信号强度。数据来源于三个重复,每个重复包含5-6株单株。不同字母表示显著差异(单因素方差分析后采用Duncan事后检验,p < 0.05)。(F) 在不同低温胁迫时间下,Micro-Tom野生型和敲除株系(CR#2和CR#13)中CBF1/2/3和SlNAC3的相对转录水平。EF-1α用作内参对照。数据显示为五个独立生物学重复的平均值,每个重复包含5-6株单株。 这些结果揭示了OE-SlNAC3株系中CBF转录本和蛋白质丰度之间存在明显的脱节,表明SlNAC3可能抑制CBF的翻译或促进CBF蛋白的降解。值得注意的是,尽管在低温处理6小时后,SlNAC3蛋白水平在野生型和OE株系之间已变得相当,但OE株系在此阶段仍表现出升高的泛素化蛋白水平(图6A)。这一观察结果表明,OE植株中较高的SlNAC3基础水平可能在低温暴露后快速激活泛素-蛋白酶体系统,从而在早期低温胁迫期间维持较高的泛素化能力。这种升高的泛素化活性可能导致了在OE株系中观察到的CBF蛋白积累减少。 总体而言,来自转录组学和蛋白质分析的结果共同提示了一个合理的机制:可溶性蛋白,尤其是CBF蛋白积累的减少,可能是OE-SlNAC3株系中观察到的冷敏感表型的基础。在响应早期低温胁迫时,SlNAC3被诱导并结合到多个促进蛋白水解的靶基因上,从而通过蛋白质稳态微调低温响应。SlNAC3的早期诱导(在低温胁迫的2小时和6小时时)对于CBF基因的转录激活可能是必要的。随着低温胁迫的持续,SlNAC3表达的随后下降(Wang等,2024)可能解除其对CBF蛋白积累的负向影响,从而增强植株在低温条件下的存活。 点击下方“阅读原文”即可阅读全文 




1
导读
2
文章详情
3
结果
精彩文章
欢迎投稿
生物组学研究
获取联系方式
关注公众号
本文来自网友投稿或网络内容,如有侵犯您的权益请联系我们删除,联系邮箱:wyl860211@qq.com 。
随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 农业农村部关于设立绿色食品产业发展专项资金的建议
- 原始的农业革命
- 工农业“剪刀差”重现?
- 自治区农业农村厅调研组莅临察雅县开展撂荒地排查整治工作
- 山西农业大学2026年山西省公费农科生录取分数!
- 天津市农业农村委关于2026年第一批主要农作物引种备案的公告
- 赵波率队赴省交通运输厅、省农业农村厅、省应急管理厅汇报对接工作
- 【银川招聘】【灵武/银川周边·农业科创】宁夏自然之星农业科技有限公司诚聘:配送中心班组长1名(4500-5000元)、保安/保洁1名(3800元)
- 2026年智慧农业创新大赛暨雄安智慧农业技术应用大赛拟参赛单位名单公示
- 奋战三季度 拼搏下半场|市农业农村局召开局系统半年度工作会议
