EST (IF:12.2)| 南京农业大学:钼酸盐对水稻土微生物砷甲基化产生靶向与非靶向抑制效应
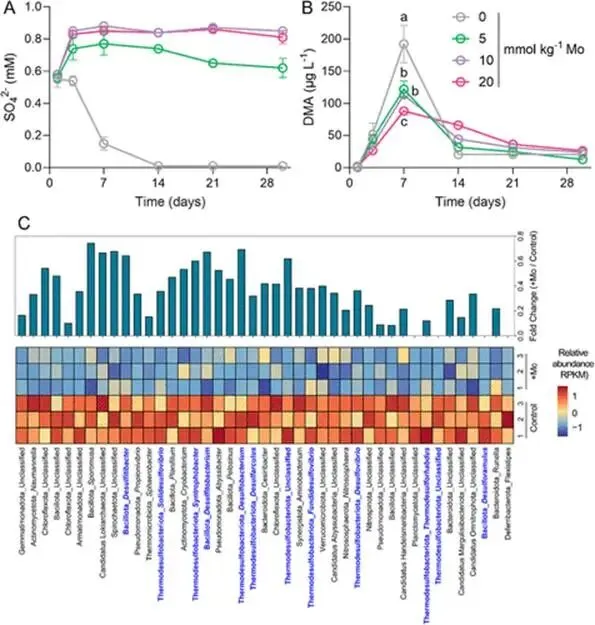
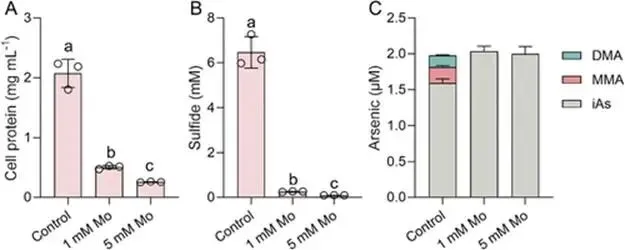
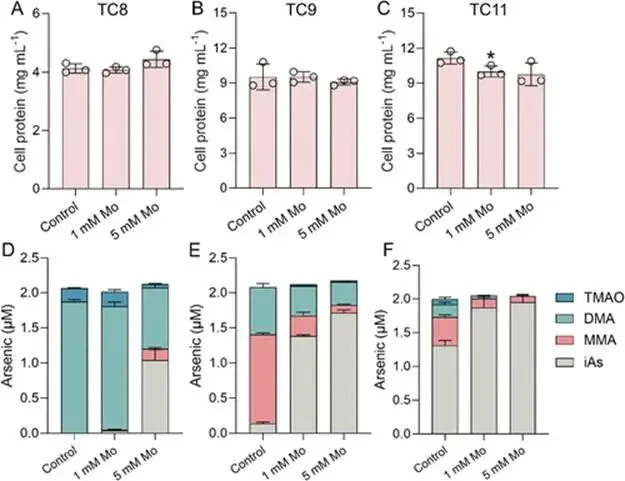
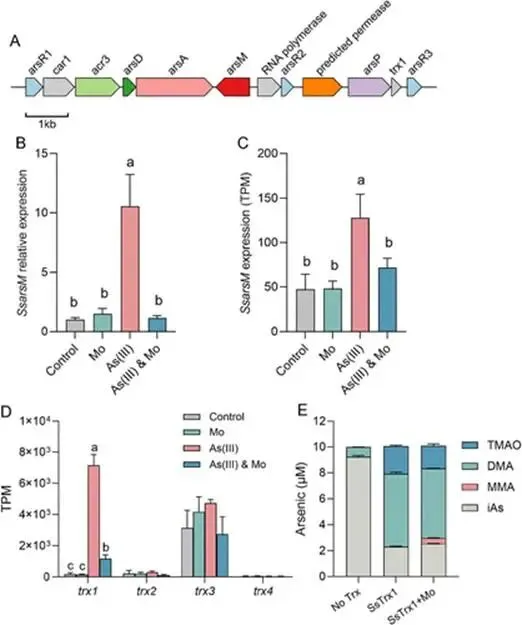
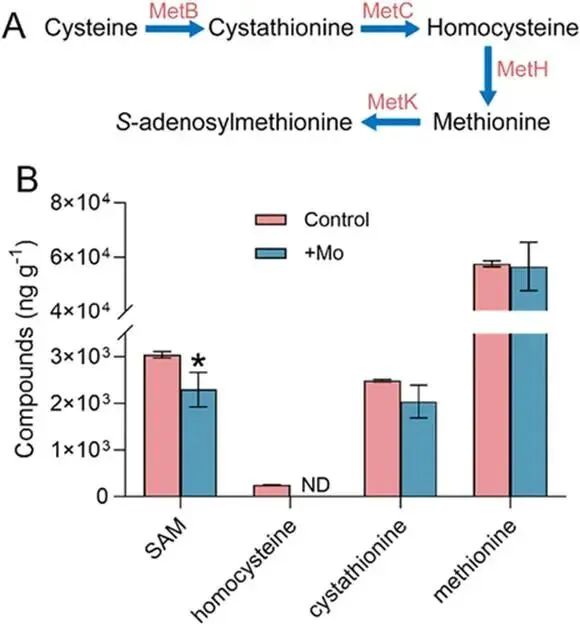
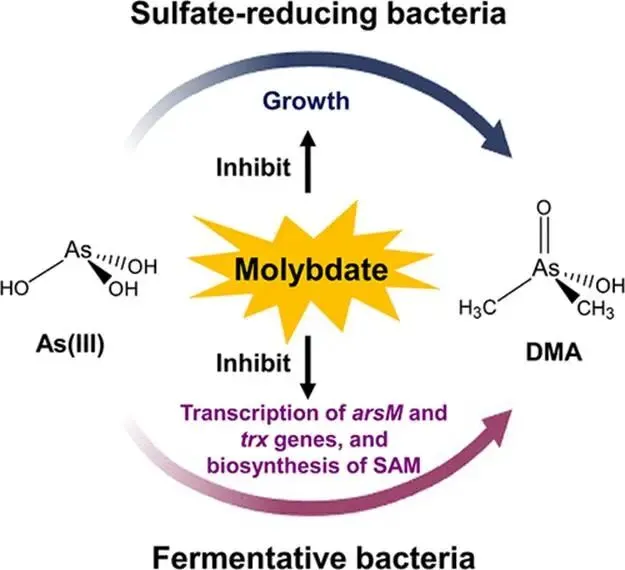
淹水水稻土中微生物砷(As)甲基化作用增强,主要产生二甲基砷(DMA),可导致水稻直穗病并造成严重产量损失。硫酸盐还原菌(SRB)被认为是主要的砷甲基化微生物,这主要基于使用钼酸盐作为SRB选择性抑制剂的实验。然而,钼酸盐抑制对SRB和非SRB介导的砷甲基化的特异性尚未得到充分评估。本研究表明,添加10 mmol kg⁻¹钼酸盐使淹水水稻土中DMA产量降低了41%,同时伴随着异化亚硫酸盐还原酶编码基因(降低78%)和亚砷酸S-腺苷甲硫氨酸甲基转移酶编码基因(arsM,降低28%)转录水平的下降。宏转录组学分析显示,添加钼酸盐显著抑制了由SRB和非SRB分别携带的12个和31个arsMs的表达。为解析钼酸盐对砷甲基化的影响,我们以从水稻土中分离的代表性SRB和非SRB发酵菌进行了纯培养实验。钼酸盐特异性抑制了SRB的生长并完全抑制了其砷甲基化活性。钼酸盐还对发酵菌的砷甲基化造成部分抑制,但不影响其生长。整合转录组学与靶向代谢组学分析揭示,钼酸盐抑制了As(III)诱导的arsM和trx(编码砷甲基化所需还原剂硫氧还蛋白)的转录,以及甲基供体S-腺苷甲硫氨酸(SAM)的生物合成,从而导致了非SRB中砷甲基化的部分抑制。这些发现表明,尽管钼酸盐特异性抑制SRB生长及其相关的砷甲基化,但它也会对发酵菌介导的砷甲基化产生非靶标效应,在解释淹水水稻土中微生物对砷甲基化的贡献时应考虑这一点。篇名: Molybdate Causes Target and Nontarget Inhibitory Effects on Microbial Arsenic Methylation in Paddy Soil期刊: Environmental Science & TechnologyDOI: 10.1021/acs.est.5c17204本研究以具高砷甲基化能力的水稻土进行淹水厌氧培养实验,设置对照组及5、10、20 mmol kg⁻¹钼酸盐添加处理,于28 °C暗处培养30天,定期监测Eh、pH,并测定硫酸盐及砷形态。一周后采集对照与10 mmol kg⁻¹处理土壤进行宏基因组与宏转录组测序,分析arsM和dsr基因丰度与表达。选取一株SRB(S. magneticus TC2)和三株发酵菌(P. benzoelyticum TC8、Romboutsia sp. TC9、Sporacetigenium sp. TC11),分别在含As(III)和不同浓度钼酸盐的培养基中培养,评估钼酸盐对生长及砷甲基化的影响。对TC11菌株进行转录组分析及RT-qPCR验证,并体外纯化其ArsM蛋白测定酶活性,同时采用LC-MS/MS靶向代谢组学分析硫代谢物变化。(1) 钼酸盐添加抑制水稻土中硫酸盐还原与砷甲基化钼酸盐的添加以剂量依赖的方式提高了土壤溶液中钼酸盐浓度,该浓度在30天培养期间下降了32–50%(支持信息,图S2A)。培养期间,对照组的土壤Eh迅速降至约−100 mV,并从第7天起保持稳定。钼酸盐添加未改变土壤Eh的动态变化(支持信息,图S2B)。土壤溶液pH在7天内从6.0升至约6.8。尽管钼酸盐因可能的聚合反应导致初始瞬时酸化,但所有处理的pH均稳定在接近中性(支持信息,图S2C)。在未添加钼酸盐的对照组中,土壤溶液中硫酸盐浓度在14天后从0.55 mM降至<0.006 mM,表明硫酸盐已被还原。相比之下,所有添加钼酸盐的处理在前3天土壤溶液硫酸盐浓度从0.55 mM初始上升至0.74–0.85 mM,这可能是由有机硫矿化所致。添加10和20 mmol kg⁻¹钼酸盐后,硫酸盐浓度从第3天起保持稳定,表明微生物硫酸盐还原被有效抑制(图1A)。无机砷(iAs)和DMA是土壤溶液中的主要砷形态(图1B,支持信息,图S2D)。对照组中DMA浓度呈短暂上升,在第7天达到峰值192 μg L⁻¹,随后在第30天降至20.6 μg L⁻¹(图1B)。钼酸盐添加显著降低了土壤溶液中DMA浓度,添加5、10和20 mmol kg⁻¹钼酸盐使其峰值浓度分别降低36%、41%和54%(图1B)。添加5或10 mmol kg⁻¹钼酸盐对土壤溶液中iAs浓度影响甚微,而较高剂量(20 mmol kg⁻¹)导致iAs显著下降(支持信息,图S2D),表明缺氧条件下iAs的迁移释放可能受到抑制。图1. 钼酸盐对淹水条件下TC水稻土中土壤溶液硫酸盐浓度指示的硫酸盐还原(A)及DMA生成(B)的影响。(C)宏转录组学分析揭示添加10 mmol kg⁻¹钼酸盐后下调的arsM基因及其宿主。arsM的SRB宿主以蓝色高亮显示。单个arsM的相对丰度基于z-score。(B)中不同字母表示在P < 0.05水平存在显著差异(Tukey HSD检验)。数据为均值±标准差(n = 3)。(2) 钼酸盐添加改变水稻土中arsM基因的丰度、转录及多样性为探究钼酸盐对arsM基因丰度、多样性及转录的影响,培养1周后采集对照组和10 mmol kg⁻¹钼酸盐处理的土壤样品,用于宏基因组学和宏转录组学分析。宏基因组分析共鉴定出490个arsM基因和1769个dsr基因,其中分别有342个arsM基因和1252个dsr基因发生转录。钼酸盐添加未显著影响宏基因组中arsM和dsr基因的丰度,但使其转录水平分别显著降低28%和78%(支持信息,图S3)。鉴定到的arsM基因的分类学归属显示其具有多样性,分布于细菌(93%)和古菌(7%)中。假单胞菌门、绿弯菌门、芽孢杆菌门、拟杆菌门、浮霉菌门、放线菌门、酸杆菌门、广古菌门、螺旋体门、热脱硫杆菌门、疣微菌门和惰杆菌门是携带arsM基因的12个优势门,占arsM宿主总量的73%(支持信息,图S4)。此外,钼酸盐添加显著下调了43个arsM基因的转录,包括来自SRB和非SRB的12个和31个arsMs(图1C)。这些差异表达的arsM基因分布于6个门,包括热脱硫杆菌门、芽孢杆菌门、拟杆菌门、绿弯菌门、假单胞菌门和放线菌门,合计占所有差异表达arsM基因的65%。基于宏基因组数据共构建了101个宏基因组组装基因组(MAGs),其中93个(92%)携带arsM基因。在含arsM的MAGs中,86个和48个分别含有trx和gshA基因,分别占arsM宿主的92%和52%,表明trx基因比gshA更为普遍。值得注意的是,48个MAG同时含有gshA和trx,且所有携带gshA的MAG均含有trx(支持信息,图S5)。(3) 钼酸盐抑制S. magneticus TC2的生长与砷甲基化利用先前从TC水稻土中分离的As(III)甲基化SRB菌株S. magneticus TC2(23),测试钼酸盐的抑制作用。培养一周后,以细胞蛋白含量测定生长(避免钼酸盐对OD600测量的干扰),发现钼酸盐添加显著抑制了S. magneticus TC2的生长和硫化物产生,且5 mM钼酸盐的抑制效果强于1 mM(图2A、B)。对照组中,S. magneticus TC2将21%的2 μM As(III)甲基化为MMA和DMA。相比之下,添加钼酸盐的培养液中未检测到甲基化砷,表明砷甲基化被完全抑制(图2C)。图2. 钼酸盐对SRB S. magneticus TC2砷甲基化的影响。(A)以细胞蛋白含量测量的细菌生长;(B)硫化物产生;(C)As(III)甲基化。(A)和(B)中不同字母表示在P < 0.05水平存在显著差异(Tukey HSD检验)。数据为均值±标准差(n = 3)。(4) 钼酸盐抑制厌氧发酵细菌的砷甲基化但不抑制其生长为探究钼酸盐对非SRB砷甲基化的影响,我们选取了从TC水稻土中分离的、具有砷甲基化能力的3株非SRB厌氧发酵细菌(P. benzoelyticum TC8、Romboutsia sp. TC9和Sporacetigenium sp. TC11)(23)。钼酸盐添加对3株非SRB菌株的生长无显著影响,仅1 mM钼酸盐对Sporacetigenium sp. TC11有轻微抑制(约10%)(图3A–C)。与S. magneticus TC2相比,这3株非SRB菌株表现出更高的砷甲基化速率,可能是由于它们在培养条件下生长更快。24小时后,菌株TC8完全甲基化了2 μM As(III),产生1.9 μM DMA和微量TMAO。添加1 mM钼酸盐对菌株TC8的As(III)甲基化影响甚微,但5 mM钼酸盐显著抑制As(III)甲基化达49%(图3D)。值得注意的是,仅在5 mM钼酸盐处理中检测到MMA。菌株TC9和TC11分别将所添加2 μM As(III)的93%和34%甲基化,主要产物为MMA。钼酸盐添加使培养液中MMA和DMA浓度分别显著降低15–56%和7–17%。钼酸盐的抑制效应呈剂量依赖性,分别使Romboutsia sp. TC9和Sporacetigenium sp. TC11的砷甲基化降低69–81%和26–29%(图3E、F)。钼酸盐添加还改变了Romboutsia sp. TC9的甲基化模式,使其主要产物由MMA转变为DMA。这些结果表明,钼酸盐对3株非SRB发酵细菌砷甲基化的抑制在很大程度上与其对生长的影响无关。图3. 钼酸盐对3株厌氧发酵菌砷甲基化的影响。(A–C)以细胞蛋白含量测量的细菌生长;(D–F)As(III)甲基化。(A、D)P. benzoelyticum TC8,(B、E)Romboutsia sp. TC9;(C、F)Sporacetigenium sp. TC11。数据为均值±标准差(n = 3)。(C)中星号表示对照组与钼酸盐处理组之间存在显著差异(P < 0.05,Tukey HSD检验)。(5) 钼酸盐抑制厌氧发酵细菌中arsM和trx基因的转录为阐明钼酸盐抑制厌氧发酵细菌砷甲基化的可能机制,我们首先研究了其对arsM表达的影响。从Sporacetigenium sp. TC11基因组中鉴定出一个典型的arsM同源物(命名为SsarsM),该基因位于一个砷抗性操纵子中。此操纵子包含As(III)响应阻遏蛋白基因arsR、As(V)还原酶基因car1、编码As(III)外排转运蛋白的acr3、编码As(III)伴侣蛋白的arsD、编码驱动As(III)外排的ATP酶的arsA、arsM、一个RNA聚合酶亚基sigma基因、一个预测的透性酶基因、编码MMA(III)外排转运蛋白的arsP,以及编码Trx的trx1基因(图4A)。RT-qPCR显示,暴露于2 μM As(III)使SsarsM转录显著上调10.5倍,而单独添加5 mM钼酸盐无显著影响。然而,当钼酸盐与As(III)共同添加时,As(III)对SsarsM转录的上调作用被完全消除(图4B)。进一步的全基因组转录组学分析证实了单独As(III)处理及As(III)与钼酸盐联合处理对SsarsM转录的影响(图4C)。在砷抗性操纵子内,SsarsP的表达同样受到As(III)增强,并在钼酸盐和As(III)同时添加时被抑制(支持信息,图S6)。这些结果表明,钼酸盐干扰了Sporacetigenium sp. TC11中SsarsM和SsarsP的As(III)依赖性转录激活。图4. 钼酸盐对发酵细菌Sporacetigenium sp. TC11中arsM和trx基因转录及ArsM体外酶活性的影响。(A)砷抗性操纵子内基因的遗传组织。(B、C)存在或不存在2 μM As(III)和5 mM钼酸盐时arsM基因的表达水平,通过(B)RT-qPCR和(C)RNA-seq检测。(D)存在或不存在2μM As(III)和5 mM钼酸盐时Sporacetigenium sp. TC11中trx基因的表达水平,通过RNA-seq检测。(E)存在或不存在5 mM钼酸盐时,利用Sporacetigenium sp. TC11的Trx系统,重组ArsM催化的砷甲基化。(B–D)中不同字母表示在P < 0.05水平存在显著差异(Tukey HSD检验)。数据为均值±标准差(n = 3)。近期研究表明,Trx作为还原剂在微生物砷甲基化中发挥关键作用(32, 33)。trx1基因位于砷抗性操纵子中(图4A),暗示其在砷甲基化中的潜在作用,特别是考虑到Sporacetigenium sp. TC11基因组中缺少另一种砷甲基化还原剂——谷胱甘肽的生物合成基因。转录组学分析显示,As(III)暴露使Sstrx1转录水平显著上调43.2倍(图4D)。当钼酸盐与As(III)共同添加时,这种上调被显著抑制,与对SsarsM转录的影响相似。Sporacetigenium sp. TC11基因组中存在另外三个trx基因(Sstrx2、Sstrx3和Sstrx4),但不在砷抗性操纵子内(支持信息,图S7)。Sstrx3转录水平相对较高,而Sstrx2和Sstrx4几乎不表达。这三个trx基因的表达对As(III)或钼酸盐均无响应(图4D)。Sporacetigenium sp. TC11中的SsarsM基因编码一个由256个氨基酸组成的ArsM蛋白,与来自Methanosarcina acetivorans C2A的MaArsM具有48%的序列一致性(34)。SsArsM拥有四个已知对催化功能至关重要的保守半胱氨酸残基(支持信息,图S8)(35)。为测试钼酸盐是否影响SsArsM的酶活性,我们在大肠杆菌BL21(DE3)中表达了来自Sporacetigenium sp. TC11的SsarsM、Sstrx1和SstrxR基因,并纯化了重组蛋白。trxR基因编码NADPH依赖的Trx还原酶,是还原氧化态Trx中二硫键所必需的。SDS-PAGE分析证实,三种重组蛋白的分子量与其理论预测值一致(支持信息,图S9)。为保持SsArsM酶活性,在SsArsM洗脱缓冲液中加入了5 mM DTT。体外检测了SsArsM的砷甲基化活性。在缺少Trx1的情况下,残留的DTT仅支持8%的As(III)甲基化为DMA(图4E)。添加Trx1后,SsArsM在2小时内将85%的As(III)转化为甲基化砷形态,主要生成DMA,伴有少量TMAO和MMA(图4E)。添加5 mM钼酸盐对以Trx1为还原剂的SsArsM催化的砷甲基化仅有轻微影响(图4E)。(7) 钼酸盐抑制Sporacetigenium sp. TC11中SAM的生物合成SAM是As(III)甲基化的必需甲基供体,由甲硫氨酸通过硫同化途径合成(图5A)。靶向代谢组学分析显示,添加5 mM钼酸盐使Sporacetigenium sp. TC11细胞中SAM和高半胱氨酸浓度分别降低25%和100%(P < 0.05),但对胱硫醚和甲硫氨酸浓度无显著影响(图5B)。图5. 钼酸盐添加对硫同化途径中代谢物浓度的影响。(A)从半胱氨酸到SAM的同化途径。(B)经或不经过5 mM钼酸盐处理的Sporacetigenium sp. TC11中甲硫氨酸、SAM、高半胱氨酸和胱硫醚的浓度。(B)中星号表示存在显著差异(P < 0.05,Tukey HSD检验)。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
