浙南山地新石器时代晚期农业结构及相关问题研究
邓振华1 李艳江1 戴 伟1 仲召兵2
(1.北京大学中国考古学研究中心,北京大学考古文博学院 2.浙江省文物考古研究所)
摘 要 作为长江下游文化区的重要组成部分,浙南山地缺乏系统植物考古研究材料的现状,限制了对当地早期农业经济特征和区域内部格局的全面认识。本文通过在江山山崖尾遗址开展系统的大植物遗存分析和农作物直接测年,首次证明当地至少在距今5300~4900 年已经形成了稻粟黍混作的农业结构,而且粟黍在整个农业体系中所占的比重较高。同时,稻米粒形的测量和对比分析显示,当地新石器时代晚期至商代种植的稻米均为“短圆形”的小粒型,作物结构和水稻品种特征均表现出与邻近的江西、福建地区较为密切的联系,明显不同于同属长江下游的环太湖地区。这一结果证明了农业经济影响因素的复杂性,以及今后相关工作中重视各大文化区内部差异,在各个小区域内系统开展基础研究的必要性。
关键词:浙南山地;山崖尾遗址;混作农业;小粒型稻
一、引言
近年来国内植物考古研究的迅速发展和基础资料的不断积累,大大推进了对我国新石器时代农业发展状况的总体认识,不仅南北方农业起源和早期发展的历程日益清晰[1],不同时期农业结构的空间格局也渐趋明朗[2]。不过,已有研究的时空分布表现出明显的不平衡,成为植物考古研究需要重视和解决的关键问题之一。这种不平衡在各区域之间和区域内部都有所表现,特别是因为对区域内部差别的相对忽视,当前农业结构时空格局的研究,往往将考古学文化区与农业经济结构两个不同层面的内容进行简单对应,造成明显的模式化倾向,影响了对早期农业经济状况的准确、全面的认识。
长江下游是我国农业起源与早期发展研究的关键地区,也是植物考古工作开展较早且相对充分的区域之一。即便如此,当地植物考古研究资料的区域分布仍然很不平衡,长江下游早期植物资源利用模式从林地和湿地野生资源与水稻共同利用,到稻作农业占绝对优势并长期延续的单线发展序列,实际上是通过在金衢盆地、宁绍平原和环太湖平原等几个相对独立的小区域之间横跳,接续而成的[3]。不同区域之间是否存在稻作农业发展序列的直接承继,金衢盆地、宁绍平原和浙南山地等地的早期农业后续如何发展,是像环太湖地区一样长期保持了单纯的稻作农业传统,还是如华南其他地区那样在新石器时代晚期形成了稻粟混作的农业结构[4],都缺乏充分的研究和论证。
因此,重视小区域之间的差异,在环太湖地区之外,系统开展有针对性的基础工作,完善小区域发展序列,进而开展对比分析和综合研究,是当前推进长江下游地区早期农业研究的突破口之一。有鉴于此,本研究在江山市山崖尾遗址开展了较为系统的植物考古采样和分析,试图为了解浙南山地地区新石器时代晚期农业经济结构提供一定的基础资料,同时尝试从农业模式的角度探讨当地与周边地区之间的联系。
二、材料与方法
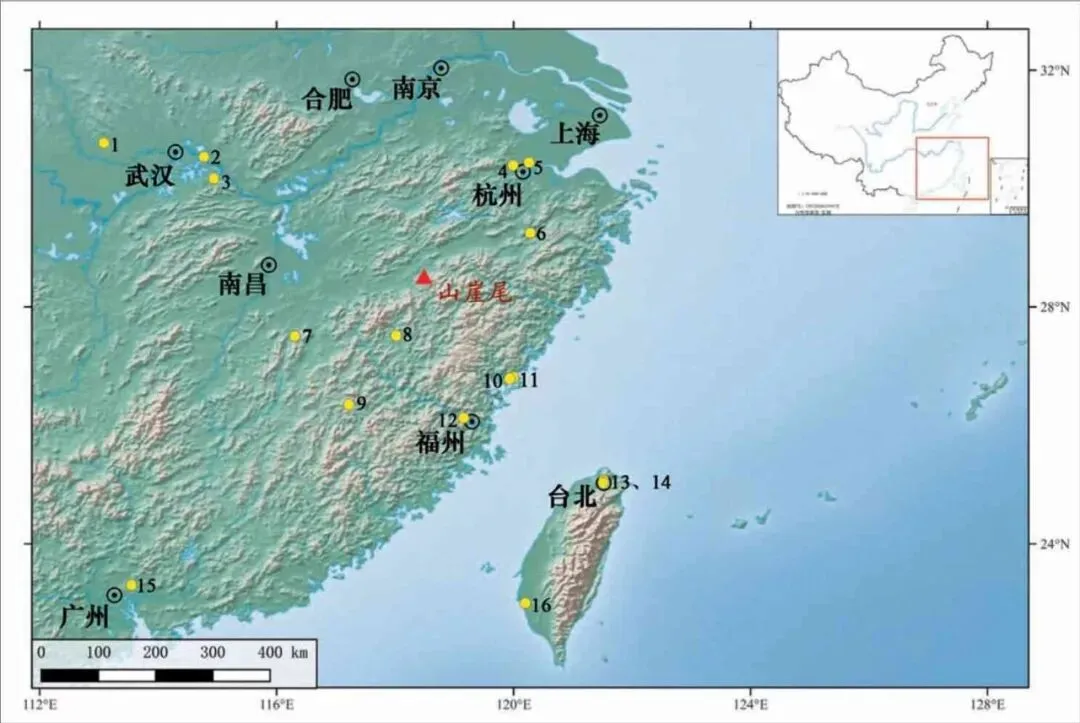
山崖尾遗址位于浙、闽、赣三省交界处,行政区划上属浙江省衢州市江山市峡口镇合新村(图一)。遗址所在地为仙霞山脉北麓一个南北向的山前岗地,海拔约179 米,总面积约1500 平方米。1978~1979 年江山地区的专题考古调查中,研究者首次发现了该遗址并进行试掘,开设探沟两条,发现了一座灰坑[5],出土的盉、豆、罐、杯等与1997 年发掘的好川墓地特征相似[6]。为了进一步明确遗址的年代、范围和文化内涵,推动好川文化的研究,2018 年浙江省文物考古研究所对遗址进行了全面勘探和再次发掘,发掘面积600 平方米,新石器时代晚期的遗迹主要有墓葬59 座,灰坑45 座。根据陶器特征和碳十四测年结果,山崖尾遗址的新石器时代晚期遗存可分为两个阶段,早期遗迹主要为灰坑,绝对年代范围大致为距今5100~4500 年(暂称之为前好川时期,年代上限或许可早至距今5300 年),晚期的主体遗存是好川文化时期的一片墓地,年代上限约为距今4500 年,年代下限不晚于距今3900 年[7]。除此之外,还发现有少量的商代遗迹。
图一 山崖尾遗址及本文涉及主要遗址位置示意图
1. 三房湾 2.城子山 3.蟹子地 4.莫角山 5.玉架山 6.尖山湾 7.锅底山 8.葫芦山 9.南山 10.黄瓜山 11.屏风山 12.白头山13.植物园 14.大龙峒 15.甘草岭 16.南关里东
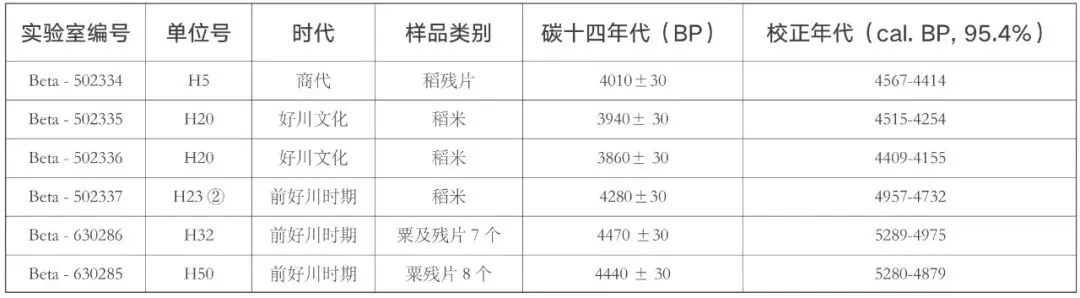
为了探索浙南山区新石器时代晚期的植物利用状况和生业经济模式,山崖尾遗址在2018年发掘过程中,按照堆积单位对大部分灰坑和少量地层进行了浮选土样的采集,共计29 个堆积单位,其中前好川时期19 个,好川文化时期4 个,商代6 个,土样量总计315.5 升。所有浮选土样均在遗址所在地使用水桶法进行了浮选,收取轻浮物的筛网孔径为0.3 毫米。所获样品在当地阴干后,送至北京大学植物考古实验室进行鉴定分析。所有轻浮物在实验室按照粒径进行了再次分筛,大于0.3 毫米的样品,在体视显微镜下放大10~15 倍进行拣选、分类、鉴定和统计,各类植物遗存的鉴定参考现代标本、已发表的鉴定标准和植物种子鉴定图谱等进行[8]。所有完整的成熟稻米,在尼康显微镜下使用NIS-Elements 软件进行了拍照测量。此外,为了进一步确定各类农作物遗存的绝对年代,本研究还对来自5 个单位的6 个农作物样品进行了碳十四测年。
三、出土植物遗存概况
山崖尾遗址三个时期的29 份样品共出土各类植物遗存2233 个,平均密度为7.1 个/升,其中密度最低的单位0.4 个/升,最高的27.4 个/升, 植物遗存含量总体偏低,且存在一定的样品间差别。这些植物遗存中可鉴定的有30 种,大部分可精确到种或属,部分因保存状况、植物自身特性等仅可鉴定到亚科或科,同时还有约2%的植物遗存(49 个)未能鉴定。
按照其生物特性及与人类的关系,可将所有的植物遗存分为农作物、果实类、禾本科杂草和其他杂草四大类。农作物包括稻(Oryza sativa)、粟(Setaria italica)、黍(Panicum milliaceum)三种(图二),共计1490 个,占所有植物遗存的66.73%。其中稻类遗存发现1366 个,占所有农作物的91.68%,是山崖尾遗址本次出土植物遗存中最主要的部分,占比达61.16%。此类遗存种类丰富,具体来讲,包括稻米、稻壳残片和小穗轴等不同部位,稻米又可根据保存状况分为完整米粒、残片(≥1/2)、碎片(<1/2,且>1 毫米)以及小碎片(<1 毫米)四类,完整米粒中大部分为成熟稻米,仅在前好川时期的两份样品中发现有3 粒不成熟稻米。小穗轴根据已建立的标准,可分为不自动落粒型(驯化型)、自动落粒型(野生型)、突出型(不成熟型)和因保存较差而无法鉴定的个体共四类。仅在好川文化时期的H7 中发现1 个稻壳顶部的小残片,稻壳其他部位残片因难以计数未进行统计。粟根据已有研究建立的分类标准可分为成熟粟、不成熟粟和极不成熟粟三类[9],此外还发现有一定量可确认为粟的种子残片,这些遗存共计107 个,占所有农作物的7.18%。黍可以根据种子形态分为成熟和不成熟两类,同时还有零星的黍残片,三类遗存共计17 个,只占所有农作物的1.14%。
图二 山崖尾遗址出土主要植物遗存
a.稻米(左:短圆形;右:长粒形) b.稻属穗轴(不自动落粒型) c.稻属穗轴(自动落粒型) d.粟 e.黍 f.狗尾草属 g.马唐属h.藨草属 i. 柿属 j.猕猴桃属 k.构树属 l.葡萄属
农作物的直接测年结果显示(表一),除商代的H5 出土稻米残片应为扰动的早期遗存外,前好川时期的3 个数据和好川文化时期的2 个数据与遗址各时期已有的绝对年代数据基本一致。特别重要的是,前好川时期H32 和H50 两个单位出土粟和粟残片的直接测年结果,确认了粟确实在前好川时期的最早阶段就已经在当地出现,是探讨粟黍南传、稻旱混作农业分布和扩散的重要证据。
表一 山崖尾遗址出土农作物遗存直接测年结果
果实类遗存总体上发现很少,可鉴定的有猕猴桃属(Actinidia sp.)、柿属(Diospyros sp.)、葡萄属(Vitis sp.)、构树(Broussonetia papyrifera)和接骨木属(Sambucus sp.),但均只有零星发现,数量最多的接骨木属种子5 粒,见于3 个单位,葡萄属种子3 粒,仅见于1 个单位,其余两种都只发现1 粒。除此之外,还有少量不可鉴定的果壳和果肉残块,果实类遗存的总量只占所有植物遗存的1.43%,可见在当时的食物资源中并不占有显著的地位。
禾本科杂草共计出土244 粒,占所有植物遗存的10.93%,但其中马唐属(Digitaria sp.)115 个,牛筋草(Eleusine indica)104 个,狗尾草属(Setaria sp.)和稗属(Echinochloa sp.)分别只发现4 粒和5 粒,此外还有少量不能确认具体种属的黍亚科和早熟禾亚科杂草。需要指出的是,出土数量最多的马唐属主要见于前好川时期的H16 ①和商代的H14 两个单位,分别发现29 个和50 个。同样的现象也见于牛筋草,前好川时期的H16 ①、好川文化时期的H7 和商代的H17分别出土27 个、49 个和21 个,其余单位发现很少。从保存状况看,部分牛筋草未完全炭化,可能源自晚期扰动。综合这些因素,山崖尾遗址本次发现的禾本科杂草总体而言种类有限且数量偏少。
其他杂草的情况与禾本科类似,虽然共计发现有467 个,占所有植物遗存的20.91%,但其中338 个为个体较小、原址性相对较差的十字花科(Brassicaceae),此外还有31 个粟米草科(Molluginaceae)和49 个未鉴定的杂草,其余杂草的总量仅49 个,包括藨草属(Scirpus sp.)、飘拂草属(Fimbristylis sp.)、春蓼属(Persicaria sp.)、青葙属(Celosia sp.)、铁苋菜(Acalypha australis)和酢浆草科(Oxalidaceae)等,各类杂草的数量均不超过10 个。
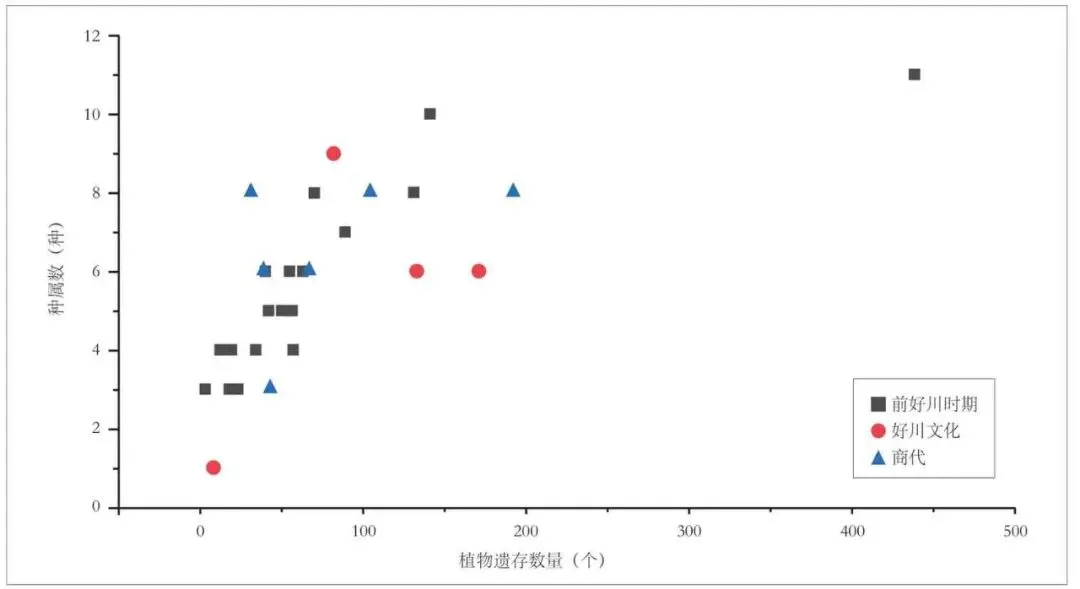
这种以农作物为主,果实类和杂草数量较少的现象,在各时期样品的植物种属多样性分布方面也有明显的体现(图三)。山崖尾遗址本次采样的29个单位中,出土植物遗存数量最多的是前好川时期的H16 ①,发现各类植物遗存438 个,其植物类别也最为丰富,共计11 种。相比之下,其他单位的植物遗存种属数量很少,大多不超过8 种,考虑到大部分单位都发现有2~3 种农作物,其他类别植物遗存的出现频率实际上非常低。总体而言,山崖尾遗址出土植物遗存的构成特征,表现出与农作物加工利用活动之间较强的相关性,伴生杂草较少的现象说明农作物的收割方式可能以掐穗为主或者遗址上进行的活动处于整个加工流程的偏后阶段。
图三 山崖尾遗址各样品出土植物遗存数量与种属数分布图
四、山崖尾遗址早期农业结构:再论稻旱混作在我国南方的分布
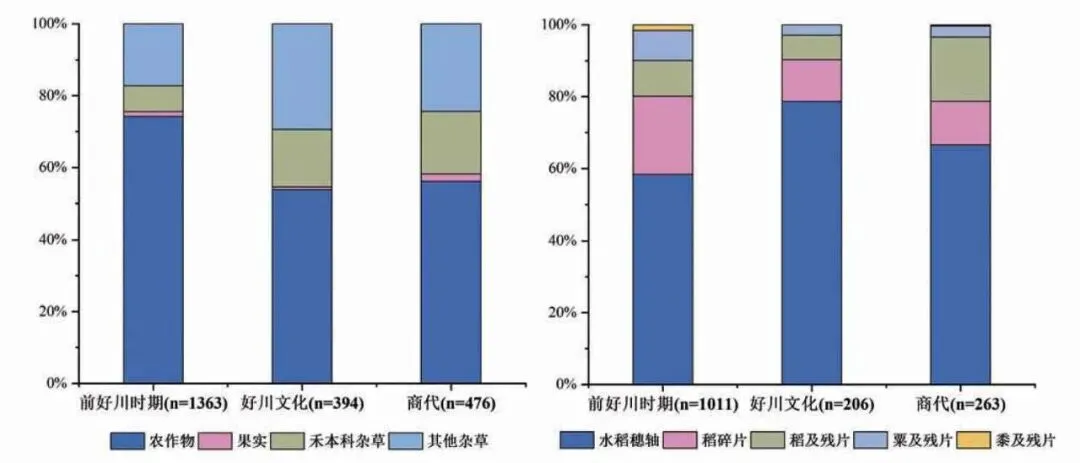
如前所述,山崖尾遗址本次发掘所获的植物遗存中农作物占绝对优势,可食用果实仅有零星发现,且均为野生浆果类,未发现宁绍平原和环太湖地区常见的橡子、菱角、芡实等富含淀粉的野生资源[10],各时期的植物遗存组合也都与此一致(图四)。因此,农业在当地新石器时代晚期和商代生业经济中的主体地位是可以肯定的。更值得关注的是,山崖尾遗址除了水稻外,还发现了数量可观的粟和黍两种旱地作物,明显不同于同时期的环太湖地区。因此,要全面认识当地早期农业结构的特点,需要对不同农作物在农业经济体系中所占的比重进行更为细致的分析。
图四 山崖尾遗址各时期不同类型植物遗存(左)与农作物(右)绝对数量百分比图
不同类型的植物因人类的使用方式、遗址埋藏条件和植物本身特性等的差异,在考古遗址中的保存状况往往存在很大的差别,因此从遗址中获取的植物遗存组合不能直接等同于古人利用的植物组合。谷物、果实和块根块茎类等几个大的植物类别之间食用方式和保存概率的差别,已经得到研究者的普遍注意,因此相关数据分析通常结合现代实验和民族学观察的结果,使用分等级的方式进行不同类别植物之间的比较[11],进而推测古代植物资源组合,而不是直接比较其数量的多寡或出现的概率。但是对谷物类内部不同作物之间的比较,目前仍以绝对数量百分比和出土概率等统计方法为主,对保存差异等因素可能导致的偏差考虑不足。
山崖尾遗址前好川时期出土各类水稻遗存911 个,粟类遗存84 个,黍类遗存仅16 个,直接对比数据的话,水稻在作物遗存组合中的绝对优势显而易见。但是,对这些植物遗存构成进行更为细致的分析就会发现,水稻遗存中590 个为小穗轴,占比高达64.76%。作为明确的作物加工过程中的废弃物,小穗轴与人类食用的米粒部位得以进入考古记录的原因截然不同,保存概率远高于米粒。然而,粟黍因其个体较小,相应的部位在遗址中保存下来的概率非常低,被发现的可能性微乎其微。若将这两类不同性质的遗存同等对待,必然会因为统计方式选择不当,人为地夸大水稻相较于粟黍的重要性。
与之类似,不同破碎程度的稻米是另一个需要关注的因素。谷物的残片如何计数和统计分析是一个比较复杂的问题,之前已有一些讨论[12],但目前并未达成系统的共识,此处不做赘述。就水稻与粟黍的比较而言,可以明确的是,稻米因其个体远大于粟黍,破碎度很高的碎片有更高的概率能够在考古遗址中得到保存。山崖尾遗址前好川时期出土稻米碎片和小碎片合计220 个,占所有稻类遗存的24.15%。本研究中区分出的大部分碎片和所有小碎片,若全部计入总量,无疑同样会拉升水稻的相对优势。
因此,在进行水稻与粟黍的数量对比时,如果考虑到小穗轴和较小的稻米碎片可能带来的偏差,不将其计算在内,则前好川时期的完整稻米(包括不成熟稻)和残片仅101 个,相比于不同成熟度的较完整粟粒及其残片的数量总和(84 个)并没有明显的优势(图四)。黍、不成熟黍和黍残片共计发现16 个,相比于水稻和粟,其在当时作物结构中所占比重很低应当是可以肯定的。即便如此,考虑到黍比粟更难在炭化过程中得到保存的情况[13],其重要性也在一定程度上被低估了。因此,综合前述影响因素和分析结果,山崖尾遗址前好川时期的作物结构应当是水稻略占优势,粟所占比重相对较高,黍可能只是作为二者的补充得到小规模种植和利用。
好川文化时期仅采集了4 个单位的45.5 升土样,共获得各类植物遗存394 个,其中农作物212 个,包括稻和粟两种,未发现黍。从农作物的构成来看,仍然是水稻穗轴最多,共计162 个,占所有稻类遗存的80.6%。稻米及其残片的数量略多于粟类遗存,但二者总数都十分有限(图四)。商代6 份样品出土的植物遗存也大体表现出同样的特征,稻类遗存最多,且68.9%为水稻穗轴。粟类遗存仅有6 粒粟、2 粒极不成熟粟和4 个粟残片,黍则仅发现1 粒不成熟黍。虽然本研究中采集的好川文化和商代样品量有限,确切的作物结构还需今后在当地开展更多有针对性的研究,但现有的证据无疑表明至少距今5000 年前后就已经形成了稻粟黍混作的作物结构,而且在浙南山地地区得到了长期延续。
此前,浦江上山遗址的新石器晚期和商周时期单位中也曾分别发现过2 粒和3 粒粟,其中新石器晚期的年代估计为距今6500 年前后的河姆渡文化时期[14]。不过,参考对粟黍南传过程的认识[15],这一年代似乎偏早。因此,山崖尾遗址的发现,首次通过系统的证据明确了浙南地区新石器时代晚期稻旱混作农业体系的存在,而且旱地作物在整个农业结构中所占的比重并不低,这与环太湖地区长期保持的单纯的稻作农业体系表现出明显的差异。整个环太湖地区,目前比较明确的粟仅见于无锡杨家遗址,马家浜中晚期的几个单位共出土9 粒粟,根据同时期稻米的测年结果,推测其年代约为距今6400 年[16]。安吉安乐遗址出土的少量粟黍,从发表的农作物照片来看,应当存在鉴定错误[17]。因此,杨家遗址的结果与环太湖地区已有的众多植物考古证据明显不同,特别是其后的崧泽良渚时期未见有明确的粟黍利用现象,当地新石器时代是否存在粟黍的种植和利用,其传入路线如何,都需要进一步的系统工作。综合现有的证据,浙南山地地区这种稻旱混作的农业模式更可能是通过邻近的江西地区传入的,这一点也与陶器特征等所反映的新石器时代晚期闽浙赣交界地带密切的文化交流状况相一致[18]。
需要注意的另一个现象是,与山崖尾遗址类似,目前在福建、台湾地区都发现有新石器时代晚期水稻与粟黍共存的证据,但在江汉平原、澧阳平原、江西和岭南地区都只见粟而未见黍,这使得目前推测的粟黍从长江中游经江西分别向岭南和福建、台湾等地的传播路线[19]是否成立存在一定的不确定性。造成这一现象的原因,一方面可能与已有研究不足有关,特别是位置至为关键的江西,仅宜黄锅底山一处新石器时代遗址开展过较为系统的植物考古研究。另一方面作物种类的选择,应当也受到了环境因素的影响,这一点在湖北的已有材料中表现得非常明显。湖北北部的郧县大寺[20]、房县计家湾[21]、保康穆林头[22]等遗址都发现有一定量的黍,在作物结构中所占的比重甚至超过了水稻,但进入江汉平原之后,所有遗址均表现出以水稻为主,兼有少量粟的作物结构。因此,如何理解粟黍传播过程中的环境和文化因素影响,以及不同区域对作物种类的选择,需要今后在各个地区进一步积累基础研究数据,特别是不同环境背景下的遗址材料才有望解决。
五、山崖尾遗址出土稻米粒形特征与新石器时代的小粒型稻
稻米是考古遗址中最为常见的一类水稻遗存,在系统的浮选法得到推广之前,大部分遗址偶然发现的水稻遗存基本均为稻米(谷),因此对其形态特征的分析也成为早期研究中探讨水稻生物性状,如野生稻还是驯化稻、粳稻还是籼稻的主要依据[23]。此后,随着相关研究的不断深化,有研究者指出稻米粒形受到遗传性状、自然环境乃至成熟度等多个因素的影响,而且现代驯化稻和野生稻的稻米粒形分布范围存在很大的重合,因此不能作为判断考古遗址出土的水稻是否被驯化的“硬”指标[24]。尽管如此,不可否认的是,人为选种、自然环境等因素综合影响下的稻米粒形,仍然有可能为我们检视同一区域不同时段乃至不同区域之间的稻种选择及其所反映的区域间联系等问题提供有益的线索。
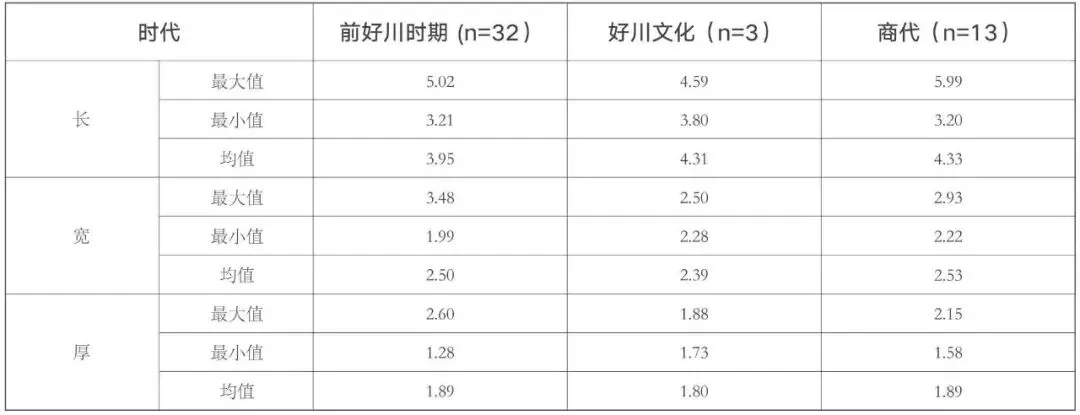
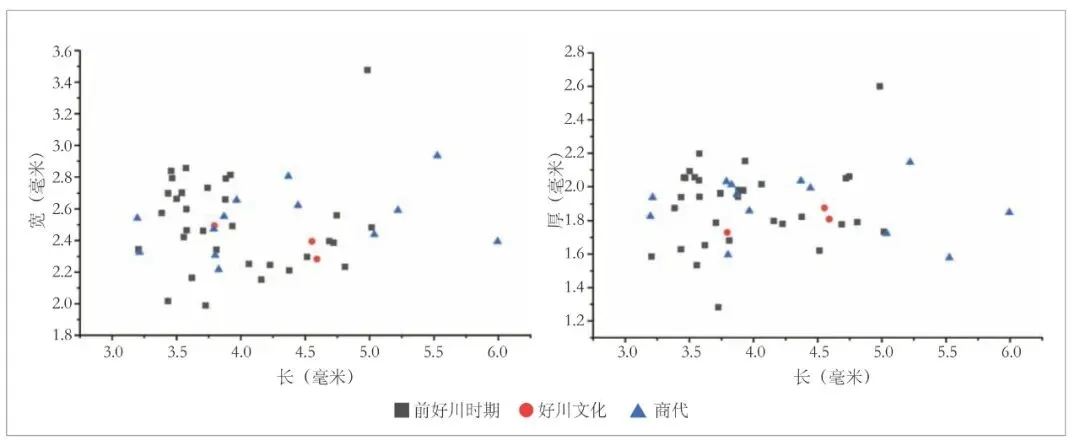
山崖尾遗址本次出土的66 粒较为完整的炭化稻米中,有48 粒能够满足测量要求,其中前好川时期32 粒,好川文化3 粒,商代13 粒。从各项指标的测量结果来看(表二,图五),山崖尾遗址各时期的稻米长度分布范围较大,介于3.20~6.00 毫米之间,但总体上长度小于4.00 毫米的稻米偏多,分别占前好川时期和商代可测量总数的65.63%和53.85%,5.00 毫米以上的前好川时期仅1 粒,商代则发现4 粒。相比之下,宽度和厚度的分布都更为集中,除前好川时期1 粒稻米的宽度达3.48 毫米外,其余稻米的宽度基本分布在2.00~3.00 毫米之间。厚度则基本位于1.50~2.60毫米之间,只有前好川时期T11 ②∶1的1 粒稻米厚度仅1.28 毫米,其长度和宽度分别为3.73 和1.99 毫米,应当是不成熟稻米。总体来讲,山崖尾遗址三个时期的稻米粒形特征基本一致,内部差异主要体现在长度方面,在宽度和厚度分化不大的情况下,稻米粒形呈现出“短圆形”和“长粒形”两种不同的趋势(图二,a),不过二者之间没有明显的分布界限,且整体上以粒形偏小的“短圆形”为主(图五)。
表二 山崖尾遗址各时期稻米测量数据分布单位:毫米
图五 山崖尾遗址各时期稻米粒形分布图
研究者很早就已经注意到这类“短圆形”稻米在新石器时代遗址中的存在,游修龄和郑云飞两位先生曾专门撰文探讨了尖山湾、石峡、绰墩和龙虬庄等遗址出土稻米中存在部分小粒型稻米的现象,并结合历史文献提出此类稻米应为密穗型,且很可能具有耐瘠、耐旱、耐寒、晚熟的生育特性[25]。需要指出的是,这几处遗址中,尖山湾遗址的稻米粒形虽然最小,但与山崖尾的相比仍然偏大,其长度分布在4.20~4.56 毫米之间,平均长度为4.38 毫米。此外,绰墩遗址的稻米长度分布在4.10~5.40 毫米之间,平均值为4.78 毫米,应当是存在两种不同粒形的稻米,且总体上偏长的粒形更多。龙虬庄和石峡遗址最短的稻米分别达4.55 毫米和4.90 毫米,与尖山湾和山崖尾相比,已经属于长粒形了。研究者对澧县城头山遗址出土的大溪文化时期稻米粒形分析也发现,所有稻米按照长度可分为七类,其中第七类属于小粒型,平均粒长4.27 毫米,占测量稻米总数的30%[26]。
此后,随着植物考古研究的逐渐展开,各个区域的新石器时代遗址都开始公布稻米测量数据,并出现了相应的分析和讨论。汉水中下游及邻近地区十余处遗址稻米的粒形分析结果显示,当地新石器时代晚期至末期的稻米粒形表现出较强的一致性,各遗址出土稻米的平均长度基本集中在4.00~4.50 毫米之间[27],表现出明显的小粒型特征。与之明显不同的是,环太湖地区新石器时代遗址出土稻米粒形比较复杂,但总体上表现出偏长的特征,大部分遗址的稻米平均长度大于4.50 毫米,田螺山偏早阶段、玉架山等遗址出土稻米的平均长度甚至超过5.00 毫米。不过,东山村和茅山遗址崧泽文化时期的稻米平均长度低于4.50 毫米,也表现出偏“短圆形”的特征[28]。由此可见,在稻作农业成熟之后,长江中下游地区的稻米粒形特征表现出一定的区域差异,无论是出于人为的选种习惯和区域传统,还是自然环境的限制,都是研究新石器时代晚期以来稻作农业发展需要关注的一个重要问题。
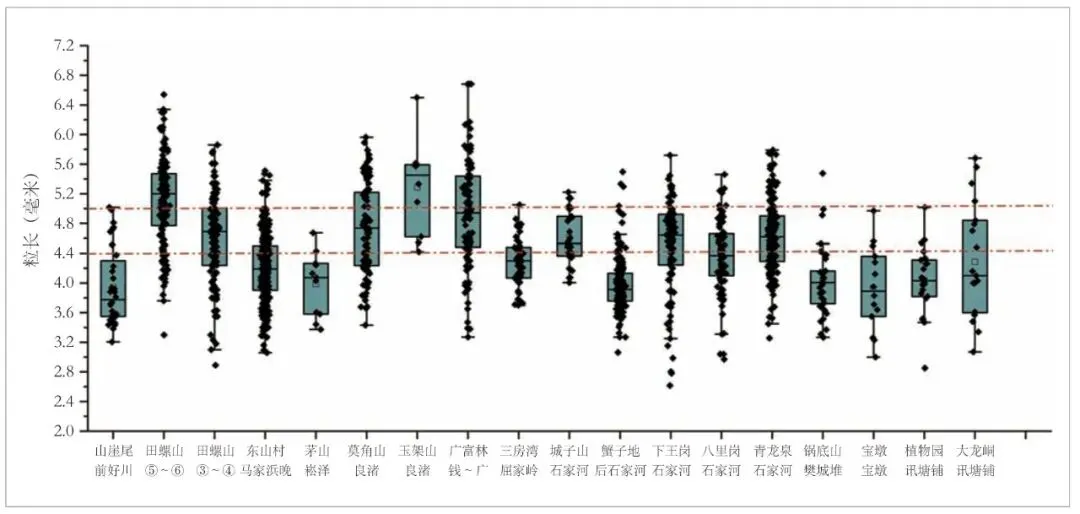
根据上述已有认识,山崖尾遗址三个时期出土的炭化稻米显然整体上属于“短圆形”的小粒型稻,为了更为细致地认识这些稻米的粒形特征以及和其他区域新石器晚期稻米的异同,本文选取了我国南方公布炭化稻米粒形数据相对较多的新石器晚期遗址进行对比分析[29]。与以往的认识基本一致,平均长度4.50 毫米是一个比较明显的分界线,长江下游地区的遗址以此为界可以被分成两类,其他地区除个别遗址略高外,稻米平均长度基本都处于这一界限以下(图六)。
图六 我国南方新石器时代晚期遗址稻米长度分布图
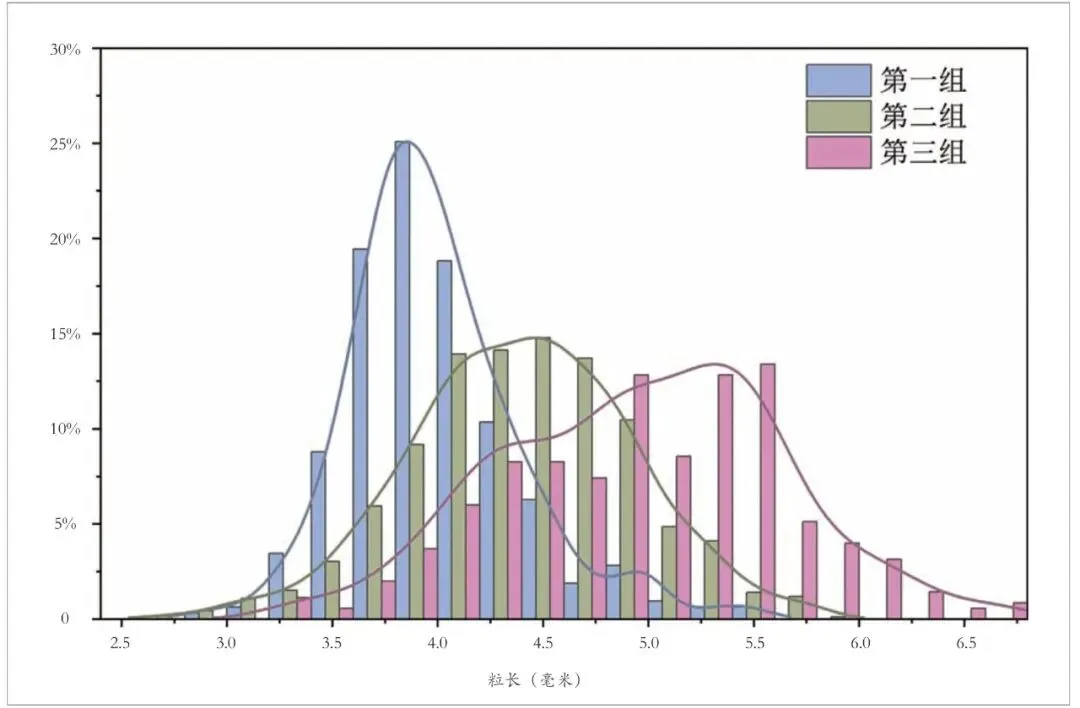
不过,平均值并不能完全反映这些遗址稻米粒形的分布状况,根据各遗址的稻米长度数据分布重心,可将其进一步分为三组:第一组粒形整体偏小,极少数粒长大于4.40 毫米,均值仅4.00 毫米上下,包括山崖尾、茅山(崧泽时期)、锅底山、蟹子地、宝墩和植物园;第二类粒形稍大,但长度大于5.00 毫米的稻米比例较低,均值在4.50 毫米上下,包括田螺山③~④层(约距今6300~6000 年)、东山村、三房湾、城子山、下王岗、八里岗、青龙泉和大龙峒;第三类粒形最大,稻米长度小于4.00 毫米的比例非常低,且有相当比例的稻米长度超过5.00 毫米,均值接近4.80毫米甚至超过5.00 毫米,包括田螺山遗址⑤~⑥层(约距今6700~6300 年)、莫角山、玉架山和广富林(钱山漾和广富林两期)。按照这一分组,对各组遗址所有稻米长度分布状况的分析也显示存在明显的组间差别,而且第一组与第二、三组的差异更为显著(图七)。
图七 新石器时代晚期三组遗址出土稻米长度分布图
综合上述分析,小粒型稻米在新石器时代晚期并非零星存在,反而普遍见于各个区域,分布范围和所占比重明显大于和高于偏长的粒形。新石器时代晚期较为典型的长粒形稻米目前主要见于环太湖地区,其他地区的遗址中占比较低。不过,根据城头山遗址稻米的粒形分布情况,澧阳平原可能也存在较高比例的长粒形稻[30],但整个区域的情况如何,尚不清楚。山崖尾遗址这种典型的“短圆形”小粒型稻,与同时期的环太湖地区大部分遗址存在明显的区别,反而与邻近的江西宜黄锅底山遗址表现出较强的相似性,这与稻旱混作的作物结构所体现的区域间联系是一致的,值得今后进一步关注。
粒长作为水稻粒形的三大构成因素之一,对水稻粒重的贡献最大,直接影响着水稻的产量,是评估稻种品质的重要指标。同时,细长型的稻米通常表现出更好的外观品质,更多地受到消费者的偏爱,改良水稻粒长也就成为现代水稻育种重要的努力方向[31]。按照这一标准,这种小粒型稻米被长期广泛种植的现象就显得很不合理。限于目前的材料基础,对于造成这一现象的具体原因,尚难给出详尽的解释。不过可以想见的是,这些稻种的粒形性状并不是独立存在的,在选择小粒型的同时很可能涉及其他更被人类看重的生物性状,例如在水稻驯化过程中,选择密穗、小粒的稻种或许还与其更易于保持非落粒性有关,新石器时代晚期及之后的广泛种植,则很可能与其更强的环境适应性存在关联。
六、结 语
山崖尾遗址出土的大植物遗存,首次提供了浙南山地地区新石器时代晚期农业经济的系统证据,分析结果表明当地至少自距今5300~4900 年开始就已经形成了以水稻为主,兼有较高比例粟和少量黍的混作农业模式,而且至少延续到商代。稻米粒形的分析显示,当地种植的水稻为“短圆形”的小粒稻,稻米平均长度不足4 毫米。结合周边地区的已有发现,无论是稻旱混作的作物结构,还是小粒型稻的种植利用,都明显不同于环太湖地区,反而表现出与邻近的江西、福建等地更强的联系,这一点也与陶器特征等所表现出的新石器时代晚期闽浙赣交界地带密切的交流互动状况相一致。这一结果证明了农业经济影响因素的复杂性,以及重视各大文化区内部多样性,在各个小区域内系统开展基础研究的重要性。
本研究得到国家重点研发计划(批准号:2020YFC1521606,2020YFC1521603)和国家自然科学基金(批准号:41872027)的共同资助。
注释
[1] 秦岭:《中国农业起源的植物考古研究与展望》,《考古学研究(九)》,文物出版社,2012 年;吕厚远:《中国史前农业起源演化研究新方法与新进展》,《中国科学:地球科学》2018 年第2 期;赵志军:《新石器时代植物考古与农业起源研究》,《中国农史》2020 年第3、4 期;邓振华:《粟黍的起源与早期传播》,《考古学研究(十三)》,科学出版社,2022 年。
[2] 袁靖主编:《中国新石器时代至青铜时代生业研究》,复旦大学出版社,2019 年。
[3] 高玉:《环太湖地区史前稻作农业研究》,北京大学博士学位论文,2017 年。
[4] Deng Z., Hung H. C., Fan X., Huang Y., Lu H.,“The Ancient Dispersal of Millets in Southern China:New Archaeological Evidence,”The Holocene, 2018,28(1): 34~43; Deng Z., Yan Z., Yu Z.,“Bridging the Gap on the Southward Dispersal Route of Agriculture in China: New Evidences from the Guodishan Site, Jiangxi Province,”Archaeological and Anthropological Sciences,2020, 12 (151): 1~10; Deng Z., Huang B., Zhang Q.,Zhang M.,“First Farmers in the South China Coast:New Evidence from the Gancaoling Site of Guangdong Province,”Frontiers in Earth Science, 2022, 10:858492.
[5] 牟永抗、毛兆廷:《江山南区古遗址墓葬调查试掘》,《浙江省文物考古研究所学刊》(第1 辑),文物出版社,1981 年。
[6] 浙江省文物考古研究所、遂昌县文物管理委员会编:《好川墓地》,文物出版社,2001 年。
[7] 仲召兵、裘佳欢:《好川文化进展与思考》,《东方博物》(第76 辑),中国书店,2020 年。
[8] Cappers R. T. J., Bekker R. M., A Manual for the Identification of Plant Seeds and Fruits, Groningen:Barkhuis & University of Groningen Library, 2013;Orton C., Sampling in Archaeology, Cambridge:Cambridge University Press, 2000;《中国农田杂草原色图谱》编委会编:《中国农田杂草原色图谱》,农业出版社,1990 年;中山至大等:《日本植物种子图鉴》,东北大学出版会,2004 年。
[9] 宋吉香、赵志军、傅稻镰:《不成熟粟、黍的植物考古学意义——粟的作物加工实验》,《南方文物》2014 年第3 期;邓振华、张海、李唯等:《河南漯河郝家台遗址早期农业结构研究》,《中国科学: 地球科学》2021 年第3 期。
[10] 傅稻镰、秦岭、赵志军等:《田螺山遗址的植物考古分析》,北京大学中国考古学研究中心、浙江省文物考古研究所编:《田螺山遗址自然遗存综合研究》,文物出版社,2011 年;高玉、秦岭:《小兜里遗址出土植物遗存分析》,浙江省文物考古研究所、海宁市博物馆编著:《小兜里》,文物出版社,2015 年。
[11] Pearsall D. M., Paleoethnobotany: A Handbook of Procedures, New York: Routledge, 2016.
[12] 高玉、邓振华:《炭化植物遗存的提取与数据分析方法浅析——以八里岗遗址2007 年浮选结果为例》,《南方文物》2016 年第2 期。
[13] 王灿、吕厚远:《黍、粟炭化温度研究及其植物考古学意义》,《东南文化》2020 年第1 期。
[14] 赵志军、蒋乐平:《浙江浦江上山遗址浮选出土植物遗存分析》,《南方文物》2016 年第3 期。
[15] 邓振华:《粟黍的起源与早期传播》,《考古学研究(十三)》,科学出版社,2022 年。
[16] 郑晓蕖、邵栋、刘宝山等:《马家浜文化生业模式初探——来自杨家遗址和马家浜遗址的植物考古学证据》,《江汉考古》2021 年第5 期。
[17] Tang Y., Marston J. M., Fang X.,“Early Millet Cultivation, Subsistence Diversity, and Wild Plant Use at Neolithic Anle, Lower Yangtze, China, ”The Holocene,2022, 32(10): 1003~1014.
[18] 黄运明、和奇:《牛鼻山文化再认识——兼论闽浙赣交界地带新石器时代晚期考古学文化的交流》,《中国考古学会第十四次年会论文集》,文物出版社,2011 年;仲召兵、裘佳欢:《好川文化进展与思考》,《东方博物》(第76 辑),中国书店,2020 年。
[19] 同[15]。
[20] 唐丽雅、黄文新、郭长江、瞿磊:《湖北郧县大寺遗址出土植物遗存分析——兼谈鄂西北豫西南山区史前农业特点》,《西部考古》(第11 辑),科学出版社,2016 年。
[21] 田洁、唐丽雅、史德勇等:《湖北房县计家湾遗址出土炭化植物遗存研究》,《南方文物》2019年第5 期。
[22] 唐丽雅、田洁、刘嘉祺等:《屈家岭文化时期山地生业模式研究——以湖北保康穆林头遗址为例》,《南方文物》2019 年第5 期。
[23] 张文绪:《中国古栽培稻的研究》,《作物学报》1999 年第4 期;张文绪、裴安平:《澧县八十垱遗址古栽培稻的粒形多样性研究》,《作物学报》2002 年第1 期;张文绪、顾海滨:《湖南澧县城头山遗址古稻研究》,《作物学报》2005 年第6 期。
[24] 秦岭、傅稻镰、Emma Harvey:《河姆渡遗址的生计模式——兼谈稻作农业研究中的若干问题》,《东方考古》(第3 集),科学出版社,2006 年;秦岭:《中国农业起源的植物考古研究与展望》,《考古学研究(九)》,文物出版社,2012 年。
[25] 游修龄、郑云飞:《从历史文献看考古出土的小粒炭化稻米》,《中国农史》2006 年第1 期。
[26] 顾海滨:《湖南澧县城头山遗址出土的新石器时代水稻及其类型》,《考古》1996 年第8 期。
[27] 邓振华:《汉水中下游史前农业研究》,北京大学博士学位论文,2015 年。
[28] 同[3] 。
[29] 此处选取的对比遗址为公布有稻米原始测量数据且单一时期数据10 粒以上者,对于一些延续多个时期且每个时期稻米测量数据均十分丰富、粒形特征无明显差异的遗址,如八里岗、下王岗、青龙泉等,只选了一个时期的数据。田螺山遗址的稻米粒形存在从早到晚逐渐变短的现象,此处选取⑤、⑥层和③、④层两组数据,未使用时代偏早、粒形更大的⑦、⑧层。数据来源:傅稻镰、秦岭、赵志军等:《田螺山遗址的植物考古分析》,北京大学中国考古学研究中心、浙江省文物考古研究所编:《田螺山遗址自然遗存综合研究》,文物出版社,2011 年;姜铭等:《新津宝墩遗址2009 年度考古试掘浮选结果分析简报》,《成都考古发现(2009)》,科学出版社,2011 年;陈航:《上海广富林遗址出土植物遗存研究》,中国社会科学院研究生院硕士学位论文,2014 年;唐丽雅:《江汉地区新石器时代晚期至青铜时代农业生产动态的植物考古学观察》, 中国社会科学院研究生院博士学位论文, 2014 年;邓振华:《汉水中下游史前农业研究》,北京大学博士学位论文,2015 年;秦岭:《东山村遗址出土植物遗存分析》,南京博物院等编著:《东山村:新石器时代遗址发掘报告》,文物出版社,2016 年;高玉:《环太湖地区史前稻作农业研究》,北京大学博士学位论文,2017 年;Deng Z., Yan Z., Yu Z.,“Bridging the Gap on the Southward Dispersal Route of Agriculture in China: New Evidences from the Guodishan Site, Jiangxi Province,”Archaeological and Anthropological Sciences, 2020, 12 (151): 1-10; Deng Z., Kuo S., Carson M.T., Hung H.C.,“Early Austronesians Cultivated Rice and Millet Together: Tracing Taiwan's First Neolithic Crops,”Frontiers in Plant Science, 2022, 10:962073。
[30]同 [26]。
[31] 张静、李晨、潘大建等:《水稻粒长遗传及其功能基因研究进展》,《广东农业科学》2021 年第3 期。
Abstract:As an integral part of the Lower Yangtze valley, the mountainous region in southern Zhejiang has been understudied in previous archaeobotanical investigations, resulting in a limited understanding of the nuanced characteristics and internal diversity of local agricultural practices. To address this gap, we conducted systematic sampling, analyzed macro plant remains, and performed direct radiocarbon dating of crops at the Shanyawei site. This study, for the first time, substantiates the existence of mixed farming in this region no later than 5300-4900 cal. BP, with a notable presence of foxtail millet and broomcorn millet in the crop assemblage during the late Neolithic period and Shang dynasty. Moreover, a grain morphometric analysis indicates that the cultivated rice at Shanyawei exhibited characteristics of a shortgrained variety, with an average length of less than 4.0 mm. These findings suggest a closer agricultural affinity between southern Zhejiang and the neighboring Jiangxi and Fujian regions in comparison to the Taihu Lake area. The outcomes of this study underscore the intricate influencing factors contributing to the formation of agricultural systems and emphasize the necessity of fundamental archaeobotanical investigations within each sub-region.
Keywords: Mountainous Region of Southern Zhejiang; The Shanyawei Site; Mixed Farming;Small-grained Rice

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
