原名:A pathogen lncRNA secreted into rice sequesters a host miRNA for virulence
中文名:一种分泌到水稻中的病原体长链非编码RNA劫持宿主miRNA以实现毒力
发表时间:2026年5月20日
DOI:https://doi.org/10.1038/s41586-026-10572-x
第一作者:Min He, Jia Su, Xiaogang Zhou, Tuo Qi, Jiazheng Wang, Tianxin Zhang, Jinhua Chen
通讯作者:贺闽(四川农业大学水稻研究所)、陈学伟(四川农业大学水稻研究所)
主要研究结果
稻瘟病是水稻生产中的重要病害,传统研究多关注病原体分泌蛋白效应子如何干扰植物免疫。该研究提出了一个新的致病模式:稻瘟病菌Magnaporthe oryzae可以将长链非编码RNA(lncRNA)转运到水稻Oryza sativa细胞中,并通过RNA与RNA之间的互补配对,劫持水稻自身的微小RNA(miRNA),从而削弱宿主免疫。
研究首先从稻瘟病菌不同发育和侵染阶段筛选出高度表达的lnc117761。敲除lnc117761后,病菌生长和附着胞形成基本正常,但侵入菌丝在水稻细胞中的扩展明显受阻,叶片病斑和菌量显著下降。相反,在水稻中异位表达lnc117761会增加感病性,说明lnc117761本身就是影响稻瘟病菌致病力的重要非蛋白效应因子。进一步分析发现,lnc117761能够与水稻miR5827配对结合。miR5827是水稻抗病的正调控因子,它通过抑制PKR1表达来增强免疫反应,而PKR1编码一个负调控免疫的受体样激酶。稻瘟病菌分泌的lnc117761进入水稻细胞后,会像分子海绵一样结合并隔离miR5827,使PKR1表达被释放,最终降低水稻抗病能力并促进病菌侵染。
lnc117761可以通过感染相关胞外囊泡从病菌转运进入水稻细胞,而且不仅存在于受侵染细胞附近,也能在相邻未见菌丝的水稻细胞中检测到。这说明稻瘟病菌并不是只在侵染点局部发挥作用,而是可以通过跨界RNA转运,提前改变周围宿主细胞的免疫状态,为侵入和扩展创造条件。miR5827相关序列在多种植物和微生物中具有较广泛存在。研究进一步把这一机制延伸到水稻纹枯病菌Rhizoctonia solani和小麦赤霉病菌Fusarium graminearum,并发现外源施用TamiR5827模拟物可以提高小麦对赤霉病的抗性。
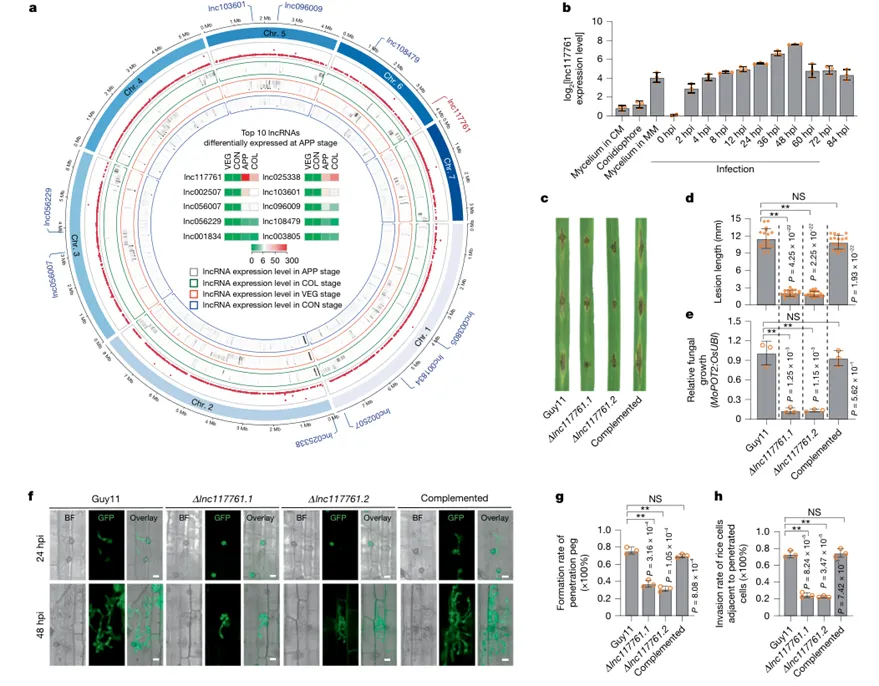
Figure 1|稻瘟病菌lnc117761参与致病并可结合水稻miR5827
该图首先展示稻瘟病菌不同发育阶段的lncRNA表达谱,其中lnc117761在附着胞和侵入菌丝阶段高度表达。敲除lnc117761后,稻瘟病菌引起的水稻叶片病斑明显缩短,菌量降低,侵入邻近细胞的能力下降,但附着胞形成并未明显受损,说明它主要影响侵染扩展过程。图中还显示,在水稻中表达lnc117761会提高感病性,并降低miR5827水平;电泳迁移率变动分析(EMSA)进一步证明lnc117761中的结合位点能够直接结合水稻miR5827。
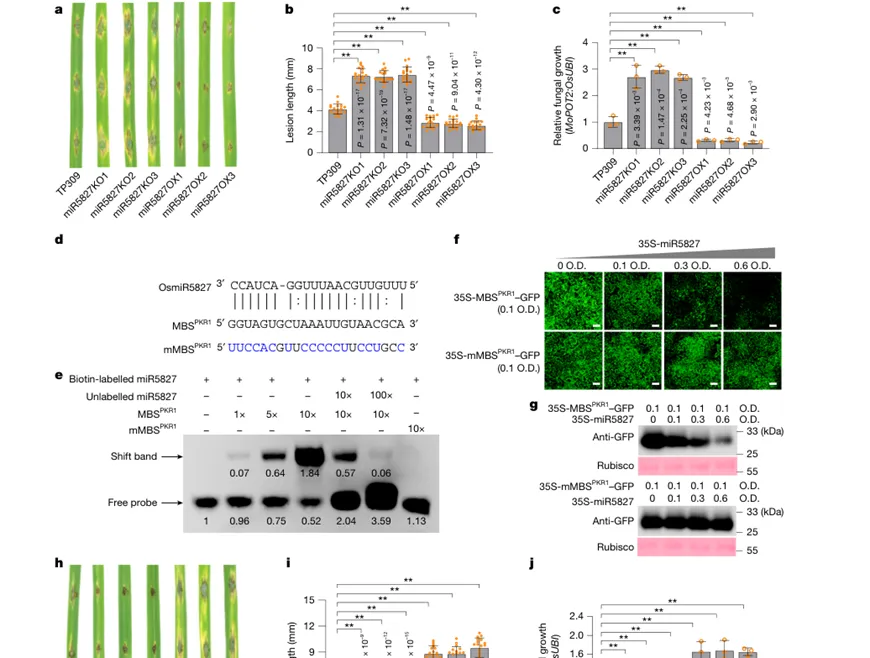
Figure 2|水稻miR5827通过抑制PKR1增强抗稻瘟病能力
该图围绕水稻miR5827的功能展开。miR5827敲除材料更感病,病斑和菌量增加;miR5827过表达材料则表现出更强稻瘟病抗性。研究进一步发现,miR5827能够直接结合PKR1 mRNA中的靶位点,并抑制其表达。PKR1敲除增强水稻抗病性,PKR1过表达则降低抗病性。也就是说,miR5827是抗病正调控因子,而PKR1是免疫负调控因子,稻瘟病菌劫持miR5827的核心后果就是解除PKR1受到的抑制。
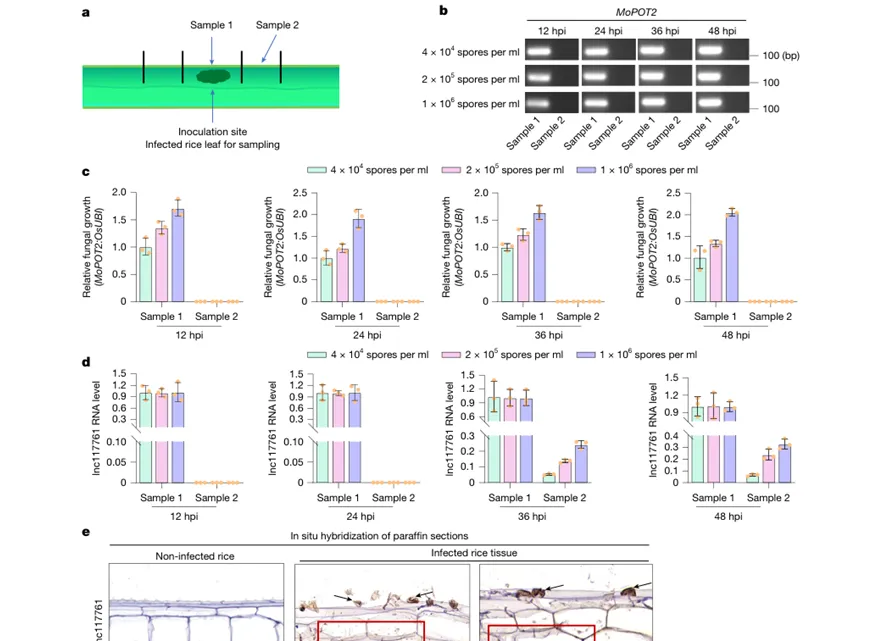
Figure 3|lnc117761可从稻瘟病菌转运进入水稻细胞
该图验证lnc117761是否真的进入水稻细胞。研究将侵染点及其周围组织分成含菌丝的样品1和无明显菌丝的样品2。结果显示,样品2中几乎检测不到病菌DNA和普通真菌mRNA,却能检测到lnc117761,说明它可以离开病菌并进入水稻组织。原位杂交进一步显示,lnc117761信号出现在侵染点、受侵染水稻细胞以及相邻未受侵染细胞中,而非转运对照mRNA只出现在病菌细胞中。这证明lnc117761是可跨界转运的RNA效应子。
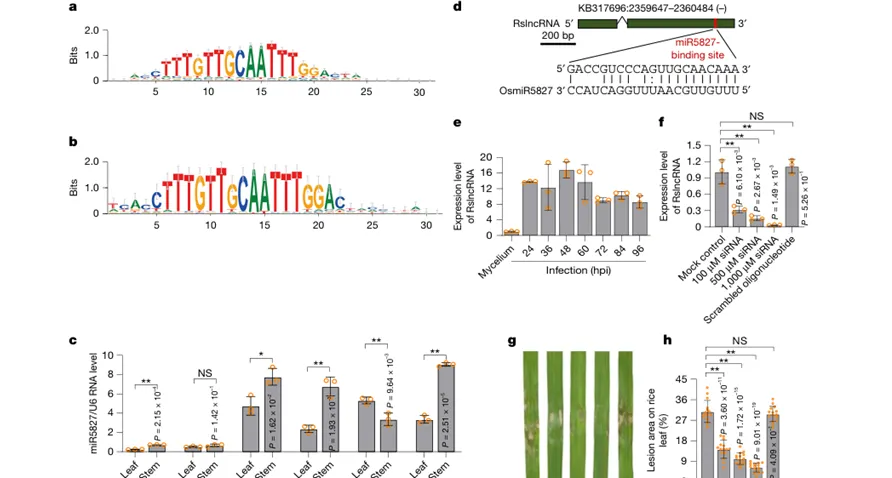
Figure 4|miR5827相关结合位点在植物和微生物中具有较广泛存在
该图把视角从水稻稻瘟病扩展到更广泛的植物和微生物。序列分析显示,与miR5827和lnc117761互补结合相关的DNA序列在704种微生物和76种植物中具有相似性。RT-qPCR也检测到大麦、拟南芥、马铃薯、二穗短柄草、小麦和水稻中存在类似miRNA表达。研究还在水稻纹枯病菌R. solani中找到可与水稻miR5827配对的RslncRNA,外源小干扰RNA降低该RNA水平后,纹枯病致病力下降,说明类似机制可能并非稻瘟病菌特有。
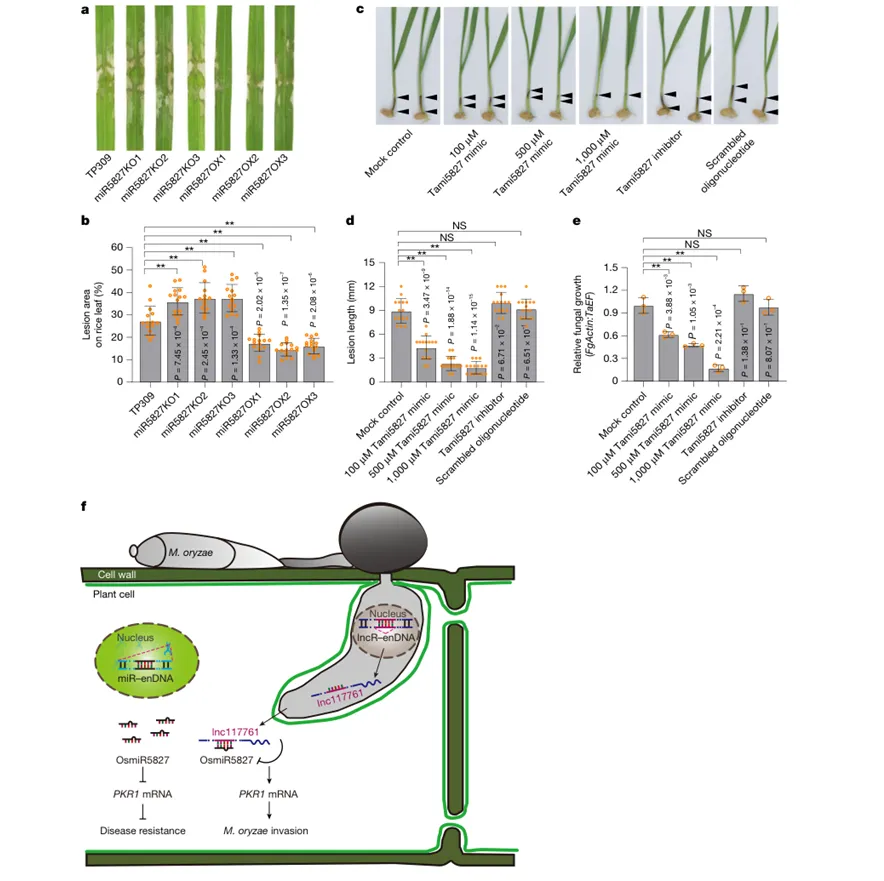
Figure 5|miR5827相关通路可用于水稻纹枯病和小麦赤霉病防控
该图进一步验证miR5827相关通路的病害防控潜力。水稻miR5827敲除材料对纹枯病更感病,过表达材料更抗病,说明miR5827不仅参与稻瘟病抗性,也能增强纹枯病抗性。研究还在小麦赤霉病菌F. graminearum中发现类似调控RNA,并证明外源施用TamiR5827模拟物可以降低小麦赤霉病病斑长度和病原菌量。模型图总结了这一机制:宿主miR5827抑制负调控免疫的PKR1,病原体lnc117761进入宿主细胞后吸附miR5827,释放PKR1表达,从而帮助病原体突破免疫防线。
总结与展望
研究将病原体非编码RNA、宿主miRNA和水稻免疫调控联系起来,揭示稻瘟病菌可以通过“RNA劫持”削弱宿主防御。病原体分泌的lnc117761不参与蛋白编码,而是进入水稻细胞后结合miR5827,使免疫负调控因子PKR1重新表达,最终促进病菌侵染。这一发现拓宽了我们对植物病原互作的认识,也提示非编码RNA可能是病原体效应体系中被低估的一类调控因子。
过去抗病育种更多关注抗病基因、免疫受体和蛋白效应子识别,而该研究表明,宿主miRNA也可以成为抗病资源。miR5827能够增强水稻对稻瘟病和纹枯病的抗性,且对主要农艺性状没有明显负面影响,具备进一步用于抗病育种或分子设计的潜力。外源TamiR5827模拟物还能提高小麦对赤霉病的抗性,说明基于RNA分子的绿色防控可能不局限于水稻体系。
未来研究可进一步在更多水稻品种和不同稻区中验证miR5827-PKR1模块的稳定性及其对产量、品质和抗病谱的影响;并评估外源miRNA模拟物在田间条件下的吸收效率、稳定性、施用方式和成本;同时继续挖掘病原体lncRNA与作物miRNA之间的互作关系,为病害预测、抗病设计和绿色植保提供新的RNA资源。