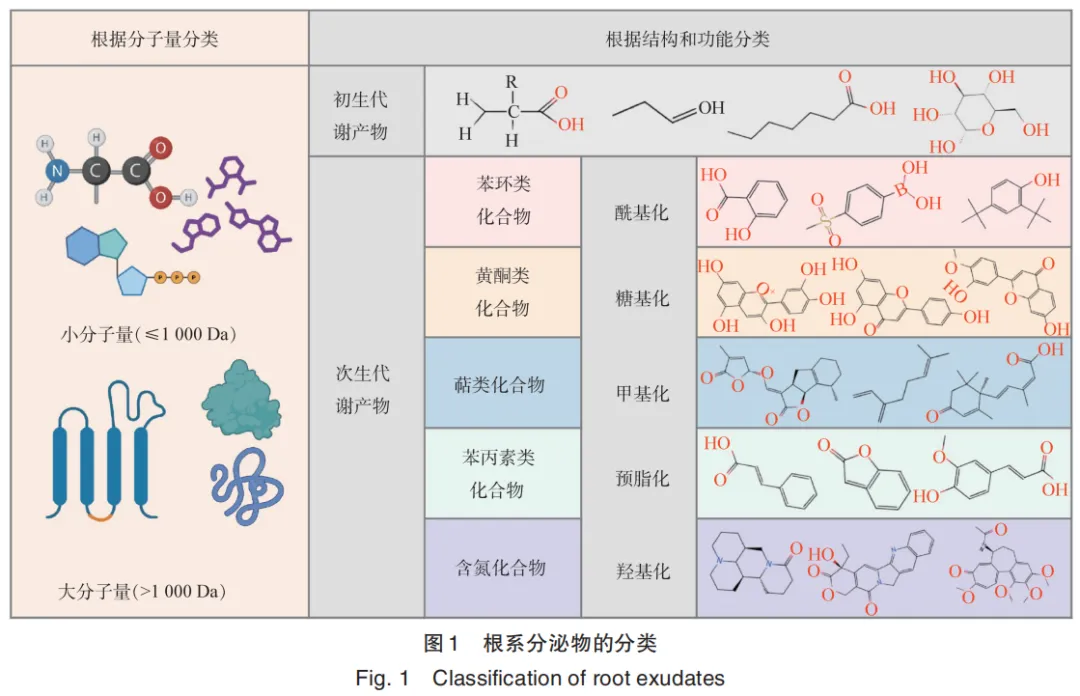
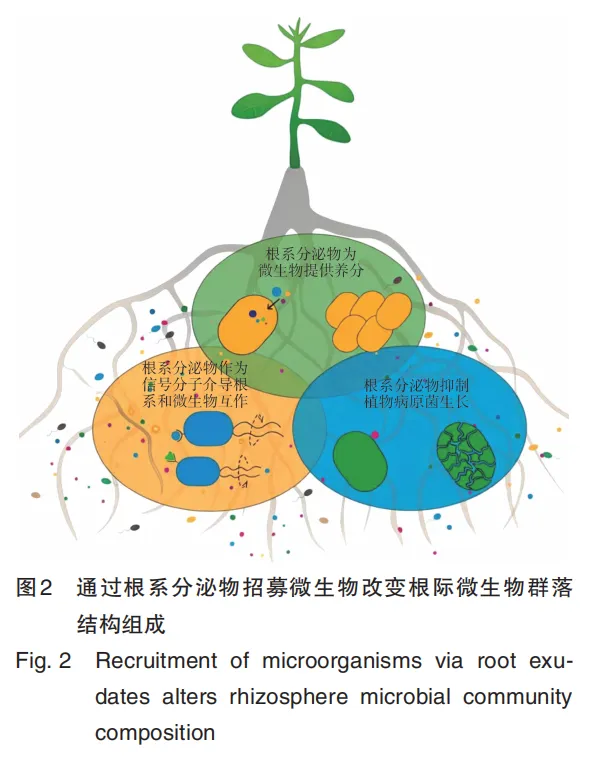
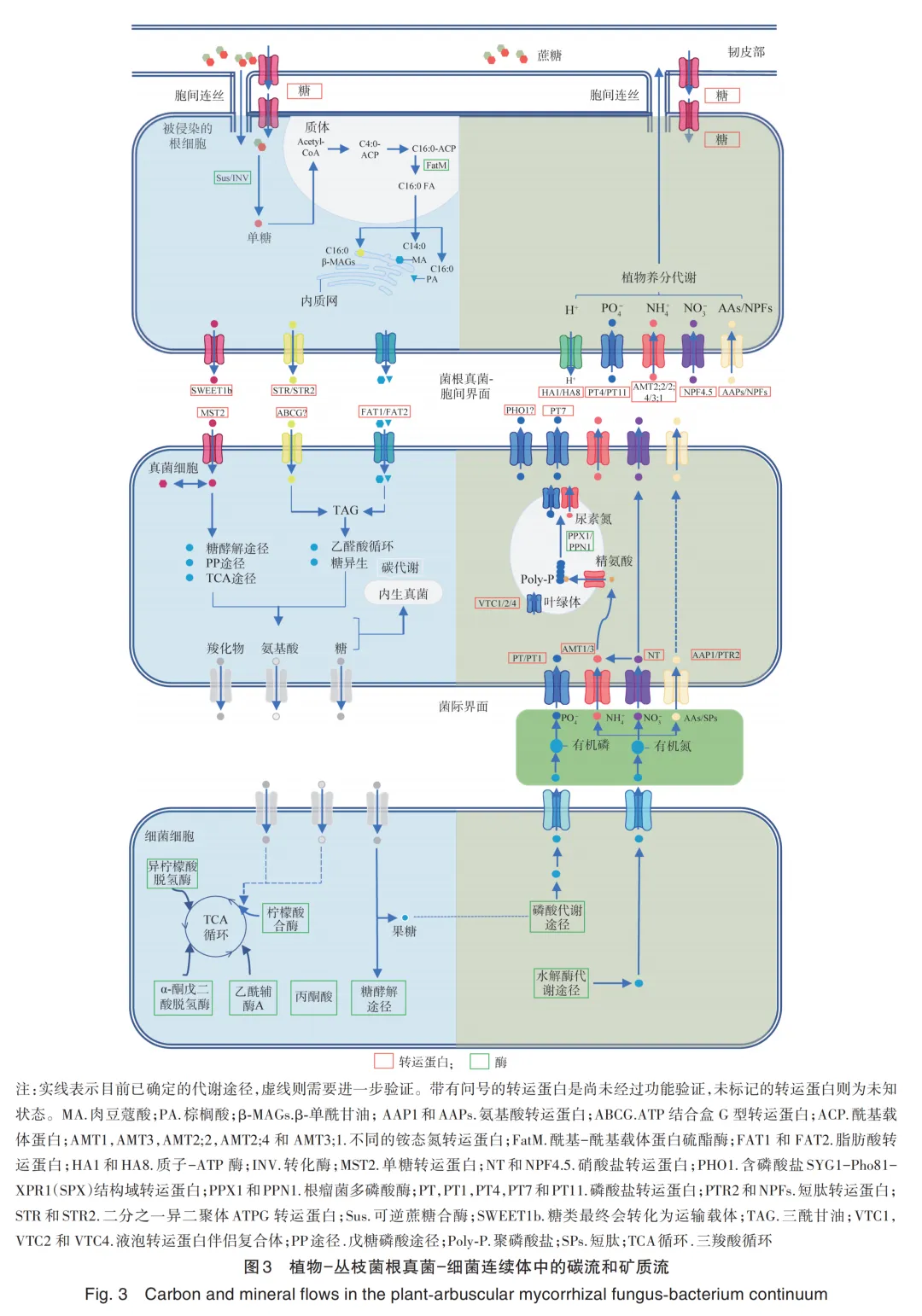
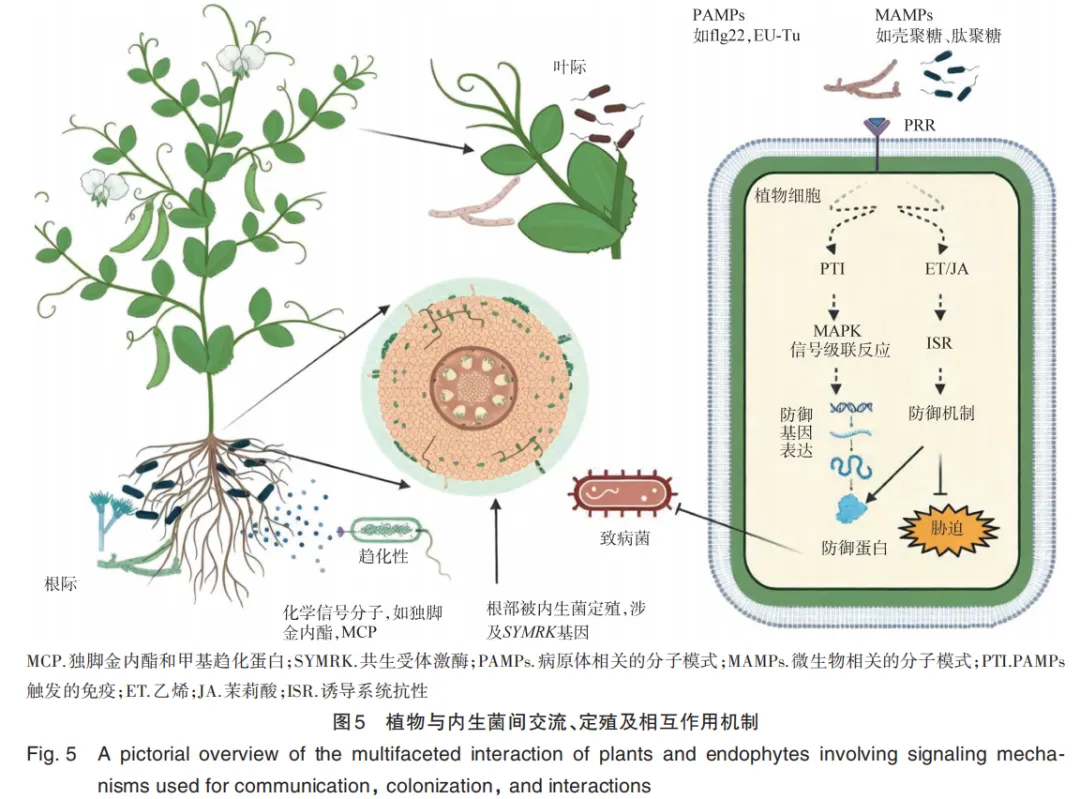
1. Li J N, Wang B, Ma H Y, et al. Root exudates drive plant⁃microbiome interactions influencing the quality of cultivated Rheum tanguticum during different growth development stages[J]. Industrial Crops and Products, 2025, 237: 122157.
2. Mazzio E, Barnes A, Badisa R, et al. Functional immune boosters; the herb or its dead microbiome? Antigenic TLR4 agonist MAMPs found in 65 medicinal roots and algae's[J]. Journal of Functional Foods, 2023, 107: 105687.
3. Sharma A, Das N, Pandey P, et al. Plant‑microbiome responses under drought stress and their metabolite‑mediated interactions towards enhanced crop resilience[J]. Current Plant Biology, 2025, 43: 100513.
4. Nikalje G C, Yadav K, Vashisht M, et al. Synergistic activity of endophytic phytobiome enhances stress tolerance and secondary metabolite production in plants[J]. Plant Stress, 2026, 19: 101140.
5. Xie L Y, Liu L F, Luo Y J, et al. Bacillus velezensis YC89-mediated recruitment of rhizosphere bacteria improves resistance against sugarcane red rot[J]. Chemical and Biological Technologies in Agriculture, 2024, 11: 114.
6. Guerrieri E, Rasmann S. Exposing belowground plant communication[J]. Science, 2024, 384(6693): 272-273.
7. Feng H C, Fu R X, Luo J Y, et al. Listening to plant's Esperanto via root exudates: Reprogramming the functional expression of plant growth‑promoting rhizobacteria[J]. New Phytologist, 2023, 239(6): 2307-2319.
8. Liu H J, Su Y W, Ye C, et al. Nucleotides enriched under heat stress recruit beneficial rhizomicrobes to protect plants from heat and root‑rot stresses[J]. Microbiome, 2025, 13: 160.
9. Tsai H H, Tang Y J, Jiang L M, et al. Localized glutamine leakage drives the spatial structure of root microbial colonization[J]. Science, 2025, 390(6768): eadu4235.
10. Liu Y P, Zhang H H, Wang J, et al. Nonpathogenic Pseudomonas syringae derivatives and its metabolites trigger the plant “cry for help” response to assemble disease suppressing and growth promoting rhizomicrobiome[J]. Nature Communications, 2024, 15: 1907.
11. Fang D L, He S D, Li L L, et al. Host plants shape rhizosphere microbiomes to counteract Fusarium: Molecular mechanisms and translational strategies[J]. Physiological and Molecular Plant Pathology, 2026, 141: 103044.
12. Chai Y N, Schachtman D P. Root exudates impact plant performance under abiotic stress[J]. Trends in Plant Science, 2022, 27(1): 80-91.
13. Warembourg F R, Billes G. Estimating carbon transfers in the plant rhizosphere[M]//The Soil-Root Interface. Amsterdam: Elsevier, 1979: 183-196.
14. Bais H P, Weir T L, Perry L G, et al. The role of root exudates in rhizosphere interactions with plants and other organisms[J]. Annual Review of Plant Biology, 2006, 57: 233-266.
15. Zhalnina K, Louie K B, Hao Z, et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly[J]. Nature Microbiology, 2018, 3(4): 470-480.
16. Jha Y, Macwan A A, Ghanaim A M, et al. Management of abiotic and biotic stresses by microbiome‑based engineering of the rhizosphere[J]. Biocatalysis and Agricultural Biotechnology, 2024, 61: 103365.
17. Wen Z H, Yu P, Shen J B, et al. Do rhizosphere microbiomes match root functional traits?[J]. Trends in Ecology & Evolution, 2025, 40(9): 885-899.
18. Fan X Y, Ge A H, Qi S S, et al. Root exudates and microbial metabolites: Signals and nutrients in plant‑microbe interactions[J]. Science China Life Sciences, 2025, 68(8): 2290-2302.
19. Wang J, Wang J R, Liu T T, et al. Bacillus amyloliquefaciens FG14 as a potential biocontrol strain against rusty root rot of Panax ginseng, and its impact on the rhizosphere microbial community[J]. Biological Control, 2023, 182: 105221.
20. Kim B S, Park S J, Kim M K, et al. Inhibitory effects of Chrysanthemum boreale essential oil on biofilm formation and virulence factor expression of Streptococcus mutans[J]. Evidence‑ Based Complementary and Alternative Medicine, 2015, 2015: 616309.
21. Carson C F, Mee B J, Riley T V. Mechanism of action of Melaleuca alternifolia(Tea tree) oil on Staphylococcus aureus Determined by time‑kill, lysis, leakage, and salt tolerance assays and electron microscopy[J]. Antimicrobial Agents and Chemotherapy, 2002, 46(6): 1914-1920.
22. Liao J M, Xia P G. Continuous cropping obstacles of medicinal plants: Focus on the plant‑soil‑microbe interaction system in the rhizosphere[J]. Scientia Horticulturae, 2024, 328: 112927.
23. 张爱华, 马文丽, 雷锋杰, 等. 人参黑斑病菌对人参根系分泌物中氨基酸的化学趋向性响应研究[J]. 中国中药杂志, 2017, 42(11): 2052-2057.
24. Ding Y J, Yao J B, Li F Q, et al. Unlocking rhizosphere phosphorus: Root exudate‑microbe synergy drives phosphorus activation in mixed Chinese fir species plantation[J]. Industrial Crops and Products, 2025, 237: 122220.
25. Liu R G, Wang C, Jin J, et al. Achnatherum inebrians tolerance to salt stress is linked with the changes of rhizosphere microbiome and root exudates[J]. Soil Ecology Letters, 2025, 7(4): 250359.
26. Li Y M, Yang F, Liu X, et al. Changes in composition and concentration of differential metabolites in root exudates are associated with aluminum‑tolerance of Ricinus communis under a high CO2 environment[J]. Plant Physiology and Biochemistry, 2024, 217: 109231.
27. Deng L M, Luo L F, Li Y, et al. Autotoxic ginsenoside stress induces changes in root exudates to recruit the beneficial Burkholderia strain B36 as revealed by transcriptomic and metabolomic approaches[J]. Journal of Agricultural and Food Chemistry, 2023, 71(11): 4536-4549.
28. Feng F J, Yang C Y, Li M J, et al. Key molecular events involved in root exudates‑mediated replanted disease of Rehmannia glutinosa[J]. Plant Physiology and Biochemistry, 2022, 172: 136-150.
29. Thakur R, Yadav S. Exopolysaccharides and biofilm forming microbial inoculant AB-13 acting in a consortium promotes growth of economically important medicinal plant Catharanthus roseus[J]. International Journal of Biological Macromolecules, 2025, 319: 145122.
30. Dong L G, Hu S L, Xu H, et al. Drought‑growth promoting bacteria enhance drought tolerance by improving root conformation of Caragana microphylla[J]. Rhizosphere, 2025, 34: 101095.
31. Ke D X, Li X Y, Han Y P, et al. ROP6 is involved in root hair deformation induced by Nod factors in Lotus japonicus[J]. Plant Physiology and Biochemistry, 2016, 108: 488-498.
32. Peng Z, Cao M Y, Zhu S D, et al. Interspecific rhizosphere interactions enhance secondary metabolite synthesis and volatile oil content in Atractylodes lancea through root exudates[J]. Industrial Crops and Products, 2025, 236: 122084.
33. Zhu L, Zhou W, Wang J F, et al. Root exudate‑mediated assemblage of rhizo‑microbiome enhances Fusarium wilt suppression in Chrysanthemum[J]. Microbiological Research, 2025, 292: 128031.
34. Guo X P, Zhang S L, Yin R S, et al. Soil nutrients, rhizosphere bacteria, and endophytes interact to influence the accumulation of amino acid‑short peptide components in Angelica sinensis roots, each playing distinct roles[J]. Rhizosphere, 2025, 33: 101027.
35. Zhao X Y, Du C Y, Zeng Q, et al. Root exudate‑microbiota interaction: Novel strategies for sustainable crop disease control[J]. Rhizosphere, 2026, 37: 101262.
36. 李笑淳, 宋凯, 陈博, 等. 植物根际促生菌: 作用机制与未来[J]. 激光生物学报, 2024, 33(3): 193-200.
37. Walker T S, Bais H P, Grotewold E, et al. Root exudation and rhizosphere biology[J]. Plant Physiology, 2003, 132(1): 44-51.
38. Iannucci A, Canfora L, Nigro F, et al. Relationships between root morphology, root exudate compounds and rhizosphere microbial community in durum wheat[J]. Applied Soil Ecology, 2021, 158: 103781.
39. Xu Z L, Zhang L, Shi X Y, et al. Response of soil enzyme activity, microbial community and root exudates in wheat rhizosphere to copper hydroxide nanopesticide[J]. Journal of Hazardous Materials, 2025, 499: 140197.
40. Fan L L, Wang J Q, Leng F F, et al. Effects of time‑space conversion on microflora structure, secondary metabolites composition and antioxidant capacity of Codonopsis pilosula root[J]. Plant Physiology and Biochemistry, 2023, 198: 107659.
41. Chen L Y, He Z, Quan Q M, et al. Increasing nitrogen addition reduces root exudates and suppresses microbial activity in the rhizosphere of medicinal Epimedium pubescens[J]. Rhizosphere, 2025, 35: 101127.
42. Nayak J, Chattopadhyay D, Giri M K, et al. Unearthing the secrets of drought‑driven root system architecture: Nutrient acquisition and rhizosphere microbe interplay[J]. Plant Science, 2026, 363: 112855.
43. Han T T, Li W Y, Li L, et al. AMF and DSE diversity and colonization characteristics of Salvia miltiorrhiza in different genotypes and growth periods[J]. Rhizosphere, 2025, 34: 101088.
44. Li S Y, Li P R, Ding H, et al. Rhizosphere bacterial community confers drought tolerance to Astragalus mongholicus[J]. Agricultural Water Management, 2025, 315: 109543.
45. Xie W, Hao Z P, Zhou J, et al. Integrated transcriptomics and metabolomics reveal specific phenolic and flavonoid accumulation in licorice (Glycyrrhiza uralensis Fisch.) induced by arbuscular mycorrhiza symbiosis under drought stress[J]. Plant Physiology and Biochemistry, 2023, 205: 108173.
46. Sorty A M, Kudjordjie E N, Meena K K, et al. Plant root exudates: Advances in belowground signaling networks, resilience, and ecosystem functioning for sustainable agriculture[J]. Plant Stress, 2025, 17: 100907.
47. de Oliveira Chagas M B, Prazeres dos Santos I, Nascimento da Silva L C, et al. Antimicrobial activity of cultivable endophytic fungi associated with Hancornia Speciosa gomes bark[J]. The Open Microbiology Journal, 2017, 11(1): 179-188.
48. Xiong Y W, Li X W, Wang T T, et al. Root exudates‑driven rhizosphere recruitment of the plant growth‑promoting rhizobacterium Bacillus flexus KLBMP 4941 and its growth⁃promoting effect on the coastal halophyte Limonium sinense under salt stress[J]. Ecotoxicology and Environmental Safety, 2020, 194: 110374.
49. Jiang M Y, Zhang K X, He L J, et al. The flavonoids in root regulated rhizosphere microbiome of Angelica dahurica var. formosana in genuine producing area[J]. Industrial Crops and Products, 2024, 219: 119164.
50. Du H B, Wang B T, Dawood M, et al. Root diameter⁃associated exudates drive the changes in rhizosphere microbial communities[J]. Journal of Soil Science and Plant Nutrition, 2025, 25(2): 2438-2450.
51. Yuan X Q, Liu Y Y, Wang S C, et al. Trifolium repens L. recruits root‑associated Microbacterium species to adapt to heavy metal stress in an abandoned Pb‑Zn mining area[J]. Science of the Total Environment, 2024, 946: 174057.
52. Vimal S R, Singh J S, Kumar A, et al. The plant endomicrobiome: Structure and strategies to produce stress resilient future crop[J]. Current Research in Microbial Sciences, 2024, 6: 100236.
53. Long X N, Bi Y, Zheng T X, et al. A multi‑faceted mechanism of Cladosporium cladosporioides‑mediated arsenic reduction in Panax notoginseng taproots: Insights from integrated multi‑omics and microbiome analyses[J]. Industrial Crops and Products, 2026, 240: 122585.
54. Wu X H, Li S H, Deng Y, et al. Multi‑omics and field experiments reveal the mechanism of Pseudomonas palleroniana P6 promoting the growth and polyphyllins accumulation in Paris polyphylla[J]. BMC Plant Biology, 2025, 25: 1214.
55. Jia H M, Zhou J, Zhao W C, et al. β-Elemonic acid mediated enrichment of Paenibacillus to help Salvia miltiorrhiza Bunge alleviate drought stress[J]. Microbiome, 2025, 13: 153.
56. Sun Q M, Wu Y T, Wang Q X, et al. Mycolicibacterium frederiksbergense SR338 enhanced root growth and active compound accumulation of Scrophularia ningpoensis by reshaping its rhizosphere microbial community and root exudate profile[J]. Industrial Crops and Products, 2026, 242: 123037.
57. Nicol R W, Yousef L, Traquair J A, et al. Ginsenosides stimulate the growth of soilborne pathogens of American ginseng[J]. Phytochemistry, 2003, 64(1): 257-264.
58. Zhang J X, Ye C, Luo L F, et al. Leaf‑herbivory induced trade‑off: Aboveground defense sacrifices root flavonoids to enrich microbes for dual resistance[J]. Agriculture, Ecosystems & Environment, 2025, 394: 109900.
59. Attia M S, Abdelaziz A M, Abdelglil M I, et al. Nutrient therapies: Boosting plant immunity for a sustainable future[J]. Physiological and Molecular Plant Pathology, 2025, 139: 102788.
60. Zhou Q, Wang Y, Yue L, et al. Impacts of continuous cropping on the rhizospheric and endospheric microbial communities and root exudates of Astragalus mongholicus[J]. BMC Plant Biology, 2024, 24: 340.
61. Duan S L, Feng G, Limpens E, et al. Cross‑Kingdom nutrient exchange in the plant‑arbuscular mycorrhizal fungus-bacterium continuum[J]. Nature Reviews Microbiology, 2024, 22(12): 773-790.
62. Rolfe S A, Griffiths J, Ton J. Crying out for help with root exudates: adaptive mechanisms by which stressed plants assemble health‑promoting soil microbiomes[J]. Current Opinion in Microbiology, 2019, 49: 73-82.
63. Feng W M, Ma R, Guo Y C, et al. Rhizosphere metagenomics and metabolomes provide new insights into the relationship between rhizosphere microecology and early bolting of Angelica dahurica[J]. Microbiological Research, 2026, 305: 128435.
64. Shafi Z, Shahid M. Root exudates as molecular architects shaping the rhizobacterial community: A review[J]. Rhizosphere, 2025, 36: 101212.
65. Guo X J, Dai H Y, Jia Z Y, et al. Reactive oxygen species in the rhizosphere orchestrate the recruitment of beneficial bacteria[J]. The EMBO Journal, 2026, 45(4): 1245-1263.
66. Mitter B, Petric A, Shin M W, et al. Comparative genome analysis of Burkholderia phytofirmans PsJN reveals a wide spectrum of endophytic lifestyles based on interaction strategies with host plants[J]. Frontiers in Plant Science, 2013, 4: 120.
67. Schulz B, Boyle C. The endophytic continuum[J]. Mycological Research, 2005, 109(6): 661-686.
68. Moricca S, Ragazzi A. Fungal endophytes in Mediterranean oak forests: A lesson from Discula quercina[J]. Phytopathology, 2008, 98(4): 380-386.
69. Biswas P R, Roy N, Ghosh S, et al. A review of molecular signaling and communication between root tissues and their endophytes[J]. Rhizosphere, 2025, 36: 101188.
70. Khan A L, Hamayun M, Kang S M, et al. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: an example of Paecilomyces formosus LHL10[J]. BMC Microbiology, 2012, 12: 3.
71. Boller T, Felix G. A renaissance of elicitors: Perception of microbe‑associated molecular patterns and danger signals by pattern‑recognition receptors[J]. Annual Review of Plant Biology, 2009, 60: 379-406.
72. Lv B C, Sun H, Cao W Y, et al. Intercropping between Panax ginseng and Arisaema amurense improves ginseng quality by improving soil properties and microbial communities[J]. Horticulturae, 2025, 11(2): 172.
73. Mei X G, Zhang L, Xu H B, et al. Intercropping improves plant biochemistry and soil microecology to realize healthy and sustainable production of Platycodon grandiflorus[J]. Journal of Cleaner Production, 2025, 507: 145529.
74. Jia H Q, Yao C H, Zhao L Z, et al. Hub metabolites at the root microbiome interface: Ecological targets for alleviating continuous cropping obstacles in Eleutherococcus senticosus[J]. Industrial Crops and Products, 2026, 240: 122593.