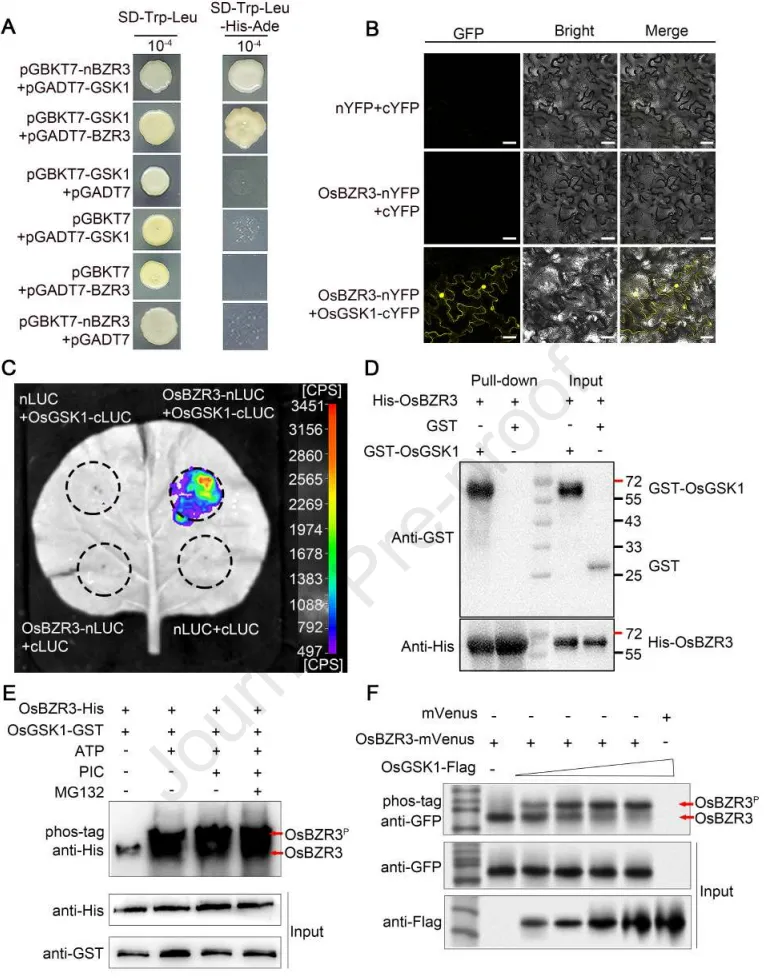
3.1 OsGSK1在油菜素内酯信号通路中与OsBZR3互作并使其磷酸化
突变体创制:利用CRISPR-Cas9技术创制了osgsk1敲除突变体,其在盐胁迫下表现出更高的存活率、更低的Na⁺/K⁺比值以及更多的侧根。
亚细胞定位:OsGSK1蛋白同时定位于细胞质和细胞核,且在盐胁迫处理后,其在细胞质中的荧光信号显著增强。
蛋白互作:鉴定出OsBZR3为OsGSK1的互作蛋白,并通过酵母双杂交、双分子荧光互补、萤光素酶互补成像及体外Pull-down等实验充分证实了二者的物理结合。
磷酸化修饰:体内和体外磷酸化实验进一步表明,OsGSK1能够直接对OsBZR3进行磷酸化修饰。
信号通路:osgsk1突变体对油菜素内酯(BR)处理表现出超敏反应,而osbzr3突变体则表现出敏感性降低,证实两者均作为水稻BR信号通路的关键组件发挥作用。
为探究BR信号传导介导水稻盐胁迫下生长恢复的机制,研究首先锁定了类GSK3激酶OsGSK1,发现其突变能显著提升水稻的耐盐性。亚细胞定位与多种互作实验证实,OsGSK1能够与BES家族转录因子OsBZR3发生物理互作,并在体内外对其进行磷酸化修饰。结合两者突变体对BR处理的相反响应表型,研究明确了OsGSK1与OsBZR3共同作用于水稻BR信号通路,为后续解析其在盐胁迫恢复中的功能奠定了分子基础。(Fig.1)
图1. OsGSK1与OsBZR3互作并使其磷酸化。 (A) Y2H实验显示OsGSK1与OsBZR3互作。(B) 本氏烟草叶片的BiFC实验表明OsBZR3与OsGSK1存在互作。比例尺= 40 μm。(C) 本氏烟草叶片的LCI实验显示OsBZR3与OsGSK1互作。(D) OsBZR3与OsGSK1的体外互作。使用结合OsBZR3-His的琼脂糖磁珠拉下指定的GST融合蛋白。使用抗His和抗GST抗体进行免疫印迹检测。(E) 在磷酸酶抑制剂PIC和蛋白酶抑制剂MG132存在下,OsBZR3-His融合蛋白作为OsGSK1-GST蛋白的底物。OsBZR3-His蛋白在phos-tag SDS-PAGE凝胶中分离,并由抗His抗体检测。(F) OsGSK1在体内磷酸化OsBZR3。OsBZR3-mVenus和OsGSK1-Flag蛋白在本氏烟草中共同表达。OsBZR3-mVenus蛋白在phos-tag SDS-PAGE凝胶中分离,并由抗GFP抗体检测。在E和F中,OsBZR3P代表磷酸化的OsBZR3蛋白。
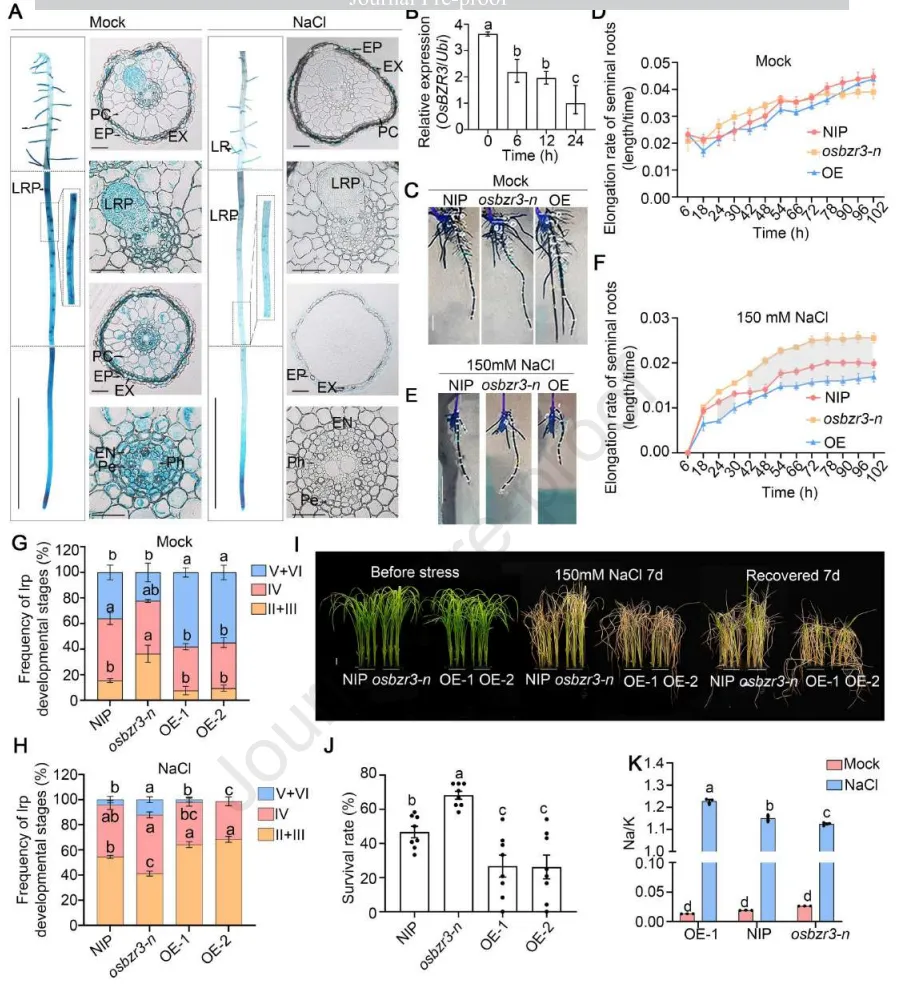
3.2 OsBZR3调控根系生长恢复与幼苗耐盐性
表达模式:OsBZR3主要在种子根伸长区及侧根原基中特异性表达,且其转录水平和启动子活性在盐胁迫下均受到显著抑制。
根系生长:在盐胁迫下,osbzr3突变体的主根分生区缩短,但其种子根伸长与细胞活力得以更好维持,而过表达株系则表现出更高的盐敏感性。
侧根发育:盐胁迫强烈抑制了野生型和过表达株系的侧根原基形成,但osbzr3突变体受到的抑制较轻,能够形成更多成熟的侧根原基。
幼苗耐盐性:在长期盐胁迫及恢复期后,osbzr3突变体表现出显著更高的存活率和更低的地上部Na⁺/K⁺比值,证明其耐盐性增强。
离子稳态:非损伤微测技术(NMT)揭示,osbzr3和osgsk1突变体根系伸长区的Na⁺外排率显著增加,表明该模块可能通过限制Na⁺外排来负调控耐盐性。
深入探究OsBZR3在盐胁迫下的生物学功能发现,该基因主要在根系活跃生长区域表达并受盐胁迫抑制。表型分析表明,OsBZR3在长期盐胁迫下作为根系生长恢复的负调控因子,其功能缺失不仅能够维持根系伸长和侧根发育,还能显著提高幼苗的整体存活率。进一步的生理检测证实,这种耐盐性的提升与突变体根系更强的Na⁺外排能力密切相关,揭示了OsGSK1-OsBZR3模块在限制离子稳态和阻碍盐胁迫恢复中的关键作用。(Fig.2)
图2. OsBZR3调控根系生长恢复与幼苗耐盐性。 (A) 0 mM或150 mM NaCl处理48小时后,5日龄pOsBZR3:GUS株系种子根的β-葡萄糖醛酸酶(GUS)染色(比例尺= 1 cm)及其横切面(比例尺= 0.1 mm)。EP=表皮,EX=外皮层,PC=薄壁组织,EN=内皮层,PE=中柱鞘,Ph=韧皮部,LR=侧根,LRP=侧根原基。(B) 150 mM NaCl处理0、6、12或24小时后,10日龄野生型NIP根系中OsBZR3的表达量(n= 4个重复)。采用Duncan多重比较法进行统计分析(p < 0.05)。(C-F) 模拟(0 mM)或150 mM NaCl处理下,野生型NIP、osbzr3-n敲除突变体和OE株系的根系伸长率(n= 10株幼苗)。比例尺= 1 cm。在D和F中,通过T检验进行统计分析(p < 0.05)。阴影区域代表统计学显著性。(G)和(H) 重力刺激用于诱导根弯曲处侧根原基(LRP)的同步形成。0 mM(模拟)(G)或150 mM NaCl (H)下重力诱导18小时后LRP阶段的频率(n= 至少30个生物学重复)。(I) NIP、osbzr3-n突变体和OsBZR3过表达株系(OE-1和OE-2)在150 mM NaCl处理7天后恢复7天的代表性图像。比例尺= 1 cm。(J) 150 mM NaCl处理7天后恢复7天的14日龄NIP、osbzr3-n和OsBZR3过表达株系OE-1和OE-2的存活率。每次重复包含24株幼苗(n=3,至少进行了两次独立实验)。(K) 150 mM NaCl处理5天的14日龄NIP、osbzr3-n和OE-1幼苗的地上部Na⁺/K⁺比值(n=3,将12株幼苗合并为一个重复)。数据为平均值±SEM。使用Duncan多重比较法进行统计分析(p < 0.05)。
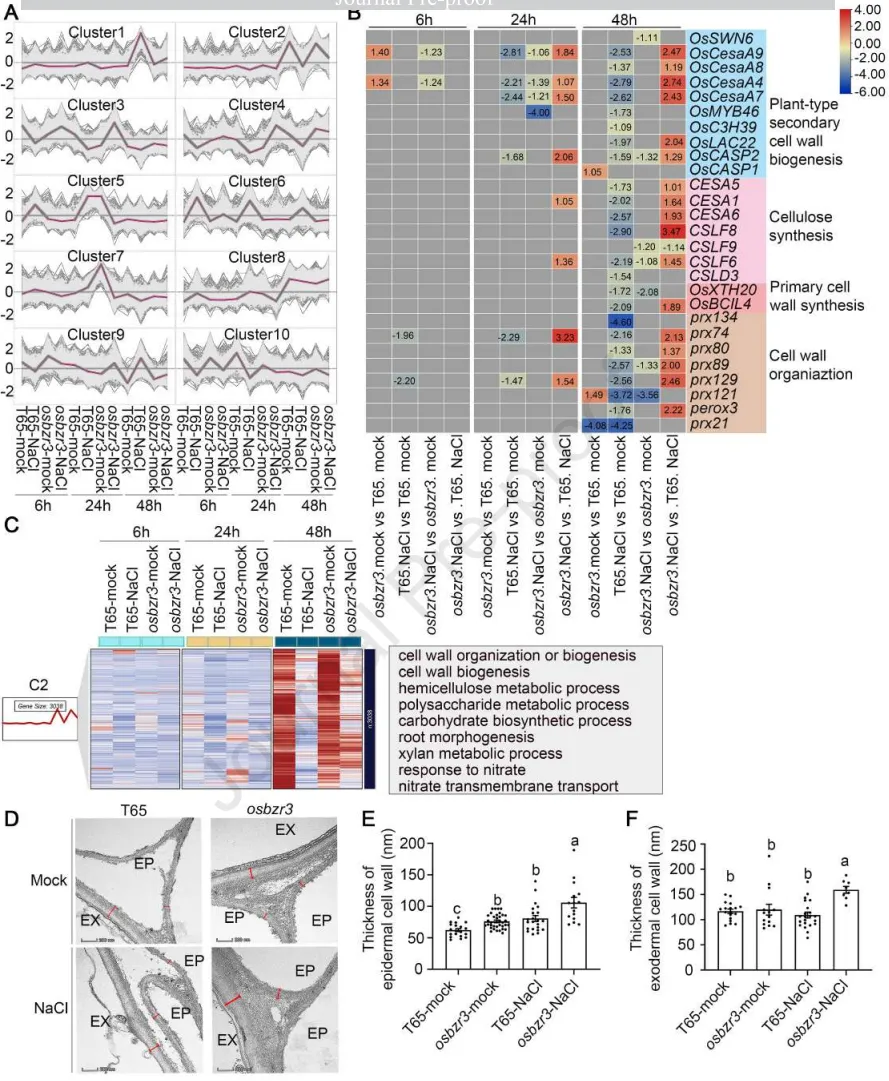
3.3 OsBZR3在根系盐恢复期间抑制细胞壁重塑基因并激活硝酸盐响应
转录组分析:时序转录组测序鉴定出大量差异表达基因,聚类分析显示这些基因在盐胁迫下呈现出早期和晚期不同的响应模式。
基因富集:晚期响应的Cluster 2基因表现出依赖于OsBZR3的转录抑制模式,且GO富集分析显示细胞壁重塑与硝酸盐响应通路高度共富集。
细胞壁基因:在盐胁迫后期,野生型中次生细胞壁合成相关基因(如CeSAs、CSLFs、PRXs)被显著抑制,而在osbzr3突变体中则得以维持。
细胞壁结构:透射电镜(TEM)观察证实,无论在正常还是盐胁迫条件下,osbzr3突变体根系表皮和外皮层的细胞壁均显著加厚且结构更完整。
为了阐明OsBZR3介导根系恢复的分子机制,研究通过转录组学分析揭示了其在盐胁迫后期的关键调控网络。分析发现,OsBZR3主要抑制了一系列与细胞壁重塑和硝酸盐响应相关的基因表达。这一转录层面的发现随后在细胞学层面得到了印证:缺失OsBZR3会导致根系表皮和外皮层细胞壁显著增厚。结合前述的Na⁺外排数据,研究推断OsBZR3通过调控细胞壁修饰来影响根系在长期盐胁迫下的离子屏障功能与生长恢复。(Fig.3)
图3. OsBZR3在根系盐恢复期间抑制细胞壁重塑基因并激活硝酸盐响应。 (A) 通过C-均值聚类将所有差异表达基因(DEGs)(|log2FC| > 1.0,padj < 0.05)分为10个簇。每个聚类中的灰色图表示单个基因的标准化表达动态,紫色线代表平均表达动态。(B) 富集在C2中的DEGs的热图表达模式和GO富集。(C) C2中细胞壁组织和生物合成途径基因的表达。(D) 在含有0 mM(模拟)或150 mM NaCl的改良Kimura液体培养基中水培4天的T65和osbzr3突变体(两个独立等位基因的合并)种子根横切面的透射电子显微镜(TEM)分析代表性图像。切片取自种子根尖上方1 cm处。EP=表皮;EX=外皮层。在36,000倍放大率下观察细胞壁厚度(比例尺=200 μm)。(E)和(F) T65、osbzr3突变体(两个独立等位基因的合并)根系表皮(E)和外皮层(F)细胞壁厚度的定量分析(n= 3个生物学重复,每个生物学重复至少测量4-5个细胞)。数据为平均值±SEM。通过Duncan多重比较法确定统计分析(p < 0.05)。
3.4 OsGSK1-OsBZR3模块协调硝酸盐信号以介导盐胁迫下的水稻生长恢复
硝酸盐响应:OsBZR3的表达特异性受KNO₃下调;盐胁迫下高硝酸盐能改变OsBZR3的亚细胞定位,并抑制OsGSK1对其的磷酸化。
根系恢复:KNO₃以剂量依赖性方式缓解盐胁迫对根系生长的抑制,其中osbzr3突变体响应最弱,而过表达株系对硝酸盐的响应最强。
幼苗存活:高浓度KNO₃能够以依赖于OsBZR3的方式,显著恢复盐胁迫下幼苗的存活率,并大幅降低地上部的Na⁺/K⁺比值。
氮素维持:在盐胁迫下,osbzr3突变体能够比野生型和过表达株系更好地维持体内总氮水平,并保持较高的根系硝酸盐吸收活性。
鉴于转录组中硝酸盐响应与细胞壁基因的共富集,研究进一步探讨了硝酸盐在OsBZR3介导的盐恢复中的作用。结果表明,高硝酸盐供应不仅能在蛋白水平上抑制OsGSK1对OsBZR3的磷酸化修饰,还能在表型上显著逆转盐胁迫对根系生长和幼苗存活的抑制。值得注意的是,这种硝酸盐介导的盐恢复高度依赖于OsBZR3的存在。同时,OsBZR3的缺失增强了植物在盐胁迫下维持氮素吸收和保留的能力,确立了OsGSK1-OsBZR3模块作为整合硝酸盐信号与盐胁迫响应的核心枢纽。(Fig.4)
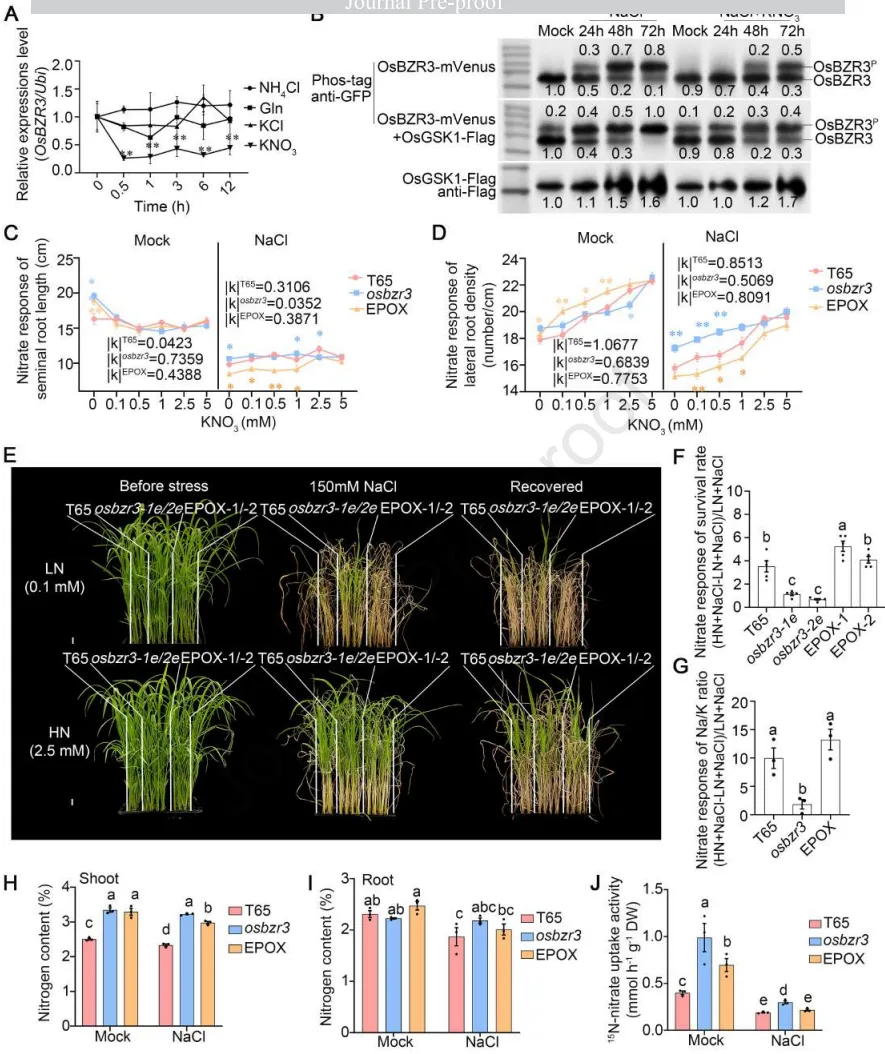
图4. OsGSK1-OsBZR3模块协调硝酸盐信号以介导水稻盐胁迫恢复。 (A) 经过不同氮源处理(0.5、1、3、6或12小时)后,水培8日龄野生型中花11 (ZH11) 幼苗根系中OsBZR3的时序表达(n= 4个重复)。在氮源处理前,幼苗经过3天的缺氮处理。NH₄Cl、谷氨酰胺(Gln)、KCl和KNO₃的浓度均为1.0 mM。使用T检验进行统计分析(与0小时相比,0.01 < p < 0.05,**p < 0.01)。(B) 在0 mM(模拟)、150 mM NaCl或150 mM NaCl结合2.5 mM KNO₃处理24、48或72小时的本氏烟草叶片中,OsBZR3-mVenus以及与OsGSK1-Flag共表达的OsBZR3-mVenus的磷酸化状态。数字表示蛋白质的相对表达水平。(C)和(D) 150 mM NaCl结合0、0.1、0.5、1.0、2.5或5.0 mM KNO₃处理7天后,2日龄T65、osbzr3和EPOX幼苗的种子根长度和侧根密度(n= 12株幼苗)。k代表折线图的斜率。随着k值的增加,侧根密度和主根长度的变化变得更加明显。通过与T65比较的T检验确定统计分析(**p < 0.01,0.01 < p < 0.05)。(E) 补充0.1 mM KNO₃ (LN) 或2.5 mM KNO₃ (HN) 并在150 mM NaCl下处理7天的14日龄T65、osbzr3-1e、osbzr3-2e、EPOX-1和EPOX-2幼苗的耐盐性测定。比例尺= 1 cm。(F) 与补充LN (KNO₃= 0.1 mM) 相比,补充HN (KNO₃= 2.5 mM) 的21日龄水稻幼苗存活率的硝酸盐响应。每次重复使用16株幼苗(n=5)。(G) 150 mM NaCl处理5天后地上部Na⁺/K⁺比值的硝酸盐响应(n= 3个重复,每个重复包含12株幼苗)。在E-G中,LN代表低浓度硝酸盐(KNO₃= 0.1 mM),而HN表示高浓度硝酸盐(KNO₃= 2.5 mM)。(H)和(I) 150 mM盐处理5天后,14日龄T65和osbzr3幼苗地上部和根系的总氮水平(n= 3个重复,每个重复包含12株幼苗)。(J) 缺氮2天后,模拟(0 mM)或150 mM NaCl条件下T65、osbzr3和OsBZR3过表达EPOX株系中的K¹⁵NO₃吸收活性(n= 3个重复,每个重复包含8株幼苗)。数据为平均值±SEM。F-J中的统计分析使用Duncan多重比较法进行(p < 0.05)。在G-J中,osbzr3突变体是两个独立等位基因的合并,EPOX株系是两个独立转基因事件的合并。
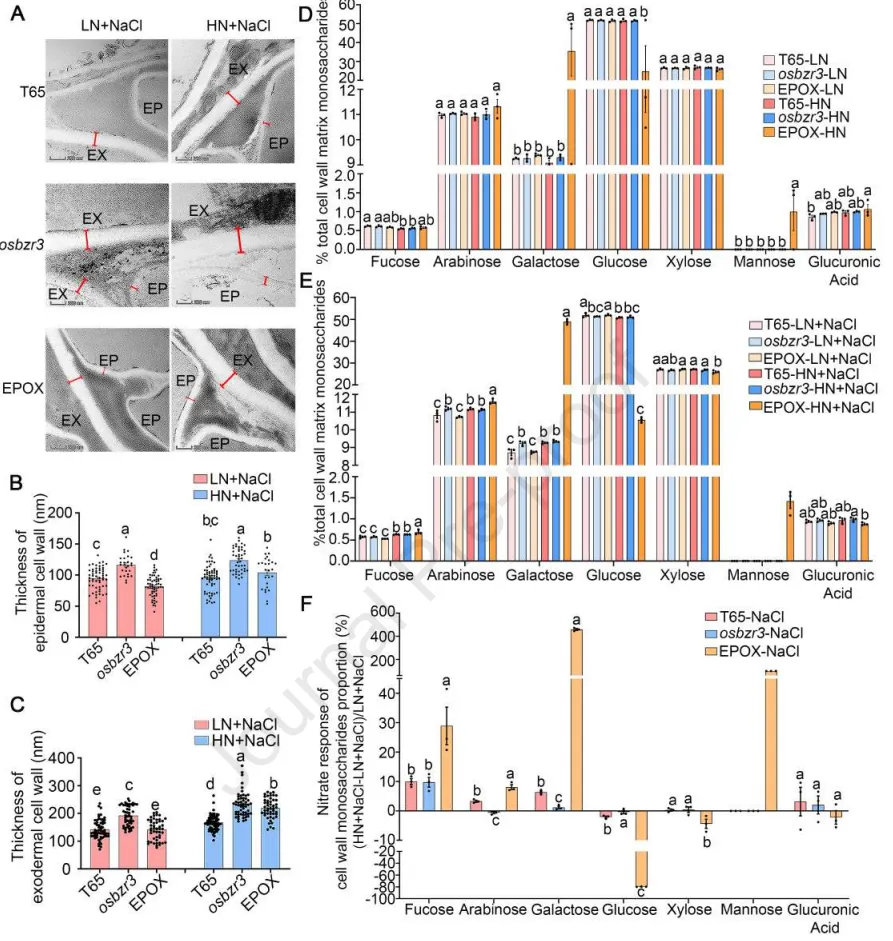
3.5 OsBZR3增强盐胁迫下细胞壁修饰的硝酸盐响应
细胞壁加厚:在盐胁迫下,高浓度KNO₃普遍促进了根系外皮层和表皮细胞壁的加厚,且在OsBZR3过表达株系中响应最为剧烈。
单糖组分:盐胁迫结合高硝酸盐处理显著改变了细胞壁中半乳糖、葡萄糖、甘露糖等单糖的组分比例,且该过程依赖于OsBZR3的活性。
结构成分:高浓度KNO₃特异性地触发了OsBZR3过表达株系在盐胁迫下纤维素和木质素的大量积累,凸显了其对结构成分的调控作用。
为深入解析OsBZR3介导的高硝酸盐诱导盐恢复机制,研究对根系细胞壁的超微结构与化学组分进行了精细分析。结果显示,高硝酸盐不仅能在物理形态上促进依赖于OsBZR3的细胞壁加厚,还能在生化组成上引发广泛的重塑,包括特定单糖比例的改变以及纤维素和木质素的积累。这些发现充分证明了OsBZR3在微调不同硝酸盐水平下的细胞壁适应性重塑中发挥着至关重要的作用。(Fig.5)
图5. OsBZR3增强盐胁迫下细胞壁修饰的硝酸盐响应。 (A) HN + NaCl (2.5 mM KNO₃ + 150 mM NaCl) 或 LN + NaCl (0.1 mM KNO₃ + 150 mM NaCl) 下根系表皮和外皮层细胞壁TEM分析的代表性图像。根切片取自10日龄T65、osbzr3和EPOX株系种子根尖上方1 cm处,这些植株在改良Kimura培养基中水培,并用150 mM NaCl结合LN (0.1 mM KNO₃) 或HN (2.5 mM KNO₃) 处理3天。在36,000倍下观察细胞壁厚度(比例尺= 200 μm)。EP=表皮,EX=外皮层。(B)和(C) 在供应低硝酸盐(KNO₃= 0.1 mM, LN)或高硝酸盐(KNO₃= 2.5 mM, HN)的150 mM NaCl下处理3天后,T65、osbzr3和EPOX种子根尖上方1 cm处根系表皮(B)和外皮层(C)细胞层的细胞壁厚度定量。数据来自3株独立植株,每株植株至少测量5个细胞。(D)和(E) 在改良Kimura培养基中水培的10日龄T65、osbzr3和EPOX株系根系中的细胞壁单糖含量,用0 mM或150 mM NaCl结合LN (KNO₃= 0.1 mM) 或HN (KNO₃= 2.5 mM) 处理4天(n= 3个生物学重复,将24株幼苗合并为一个重复)。(F) 150 mM NaCl结合LN或HN处理4天后,10日龄水稻幼苗根系中细胞壁单糖含量的硝酸盐响应(n=3)。数据为平均值±SEM。B-I中的统计分析使用Duncan多重比较法进行(p < 0.05)。osbzr3突变体是两个独立等位基因的合并,EPOX株系是两个独立转基因事件的合并。在A至F中,LN代表低硝酸盐(KNO₃),HN代表高硝酸盐(KNO₃)。
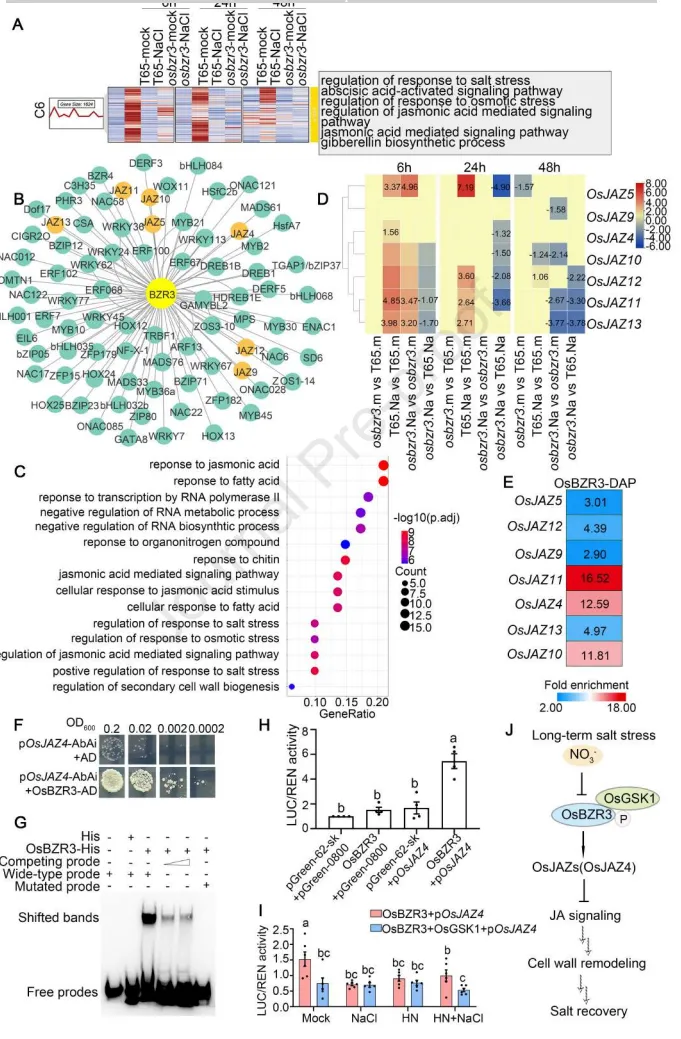
3.6 OsBZR3在OsJAZ4上游发挥作用,以响应盐胁迫和硝酸盐可用性来调节JA信号传导及其下游细胞壁重塑
靶基因预测:通过对转录组Cluster 6基因的网络推断分析,发现茉莉酸(JA)信号通路及多个OsJAZ基因是OsBZR3的潜在下游靶标。
启动子结合:DAP-seq、酵母单杂交及凝胶阻滞实验(EMSA)确证,OsBZR3能够直接特异性结合到OsJAZ4等基因的启动子区域。
信号抑制:双萤光素酶实验表明,OsBZR3能激活OsJAZ4的启动子,但该激活作用会被OsGSK1、盐胁迫及高硝酸盐处理所抑制。
细胞壁重塑:osjaz4突变体在盐胁迫下表现出与osbzr3相似的表型,包括细胞壁相关基因的表达失调以及维持更高的侧根密度。
为了将硝酸盐信号、细胞壁适应与激素网络联系起来,研究深入挖掘了OsBZR3的直接转录靶标。通过多组学结合分子生物学手段,证实了OsBZR3能够直接结合并激活JA信号核心抑制因子OsJAZ4。更重要的是,这种激活作用受到上游OsGSK1激酶、盐胁迫以及高硝酸盐信号的联合抑制。结合osjaz4突变体在盐胁迫下表现出的细胞壁基因表达紊乱及耐盐表型,研究最终勾勒出一条完整的信号通路:OsGSK1-OsBZR3模块通过整合硝酸盐信号来微调OsJAZ4的活性,进而调控JA介导的细胞壁重塑,最终实现水稻在长期盐胁迫下的生长恢复。(Fig.6)
图6. OsBZR3在OsJAZ4上游发挥作用以调节JA信号传导及其下游细胞壁重塑。 (A) 簇6 (C6) 中DEGs的热图表达模式和GO富集。(B) 使用GRNBoost2对簇6中基因进行的OsBZR3下游直接靶标的转录调控网络推断。(C) C6中OsBZR3中心网络转录因子的GO(生物学过程)富集分析。列出了前15个GO术语。(D) 模拟(m)或150 mM NaCl (Na)处理6、24或48小时后,T65和osbzr3突变体根系中OsJAZs的表达模式(log2FC)。(E) 使用DAP-seq分析的OsBZR3靶标全基因组分析中OsJAZs基因的富集。(F) 在Y1H实验中,OsBZR3靶向OsJAZ4的启动子。(G) 凝胶阻滞实验验证了OsBZR3直接结合到OsJAZ4启动子中的相应基序。His蛋白用作阴性对照。(H) 双萤光素酶报告基因实验显示OsBZR3对OsJAZ4启动子的激活作用(n=4)。(I) 双萤光素酶报告基因实验表明,在0 mM或150 mM NaCl结合2.5 mM KNO₃下,OsBZR3对OsJAZ4启动子的激活受到OsGSK1存在的影响(n=6)。LN代表低硝酸盐(KNO₃),HN代表高硝酸盐(KNO₃)。在H和I中,数据为平均值±SEM。通过Duncan多重比较法确定统计分析(p < 0.05)。(J) OsGSK1-OsBZR3级联整合硝酸盐信号以调节OsJAZ4并影响下游细胞壁重塑,从而协调长期胁迫下水稻根系的盐恢复和稳态。P代表蛋白质磷酸化。实线箭头代表被证明具有正向调节作用的步骤,而实线平头箭头代表负向调节作用。虚线箭头代表需要进一步探索的机制步骤。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
