在农业害虫防治的战场上,杆状病毒因其高致病性和环境友好性,被誉为“绿色生物农药”。然而,它有一个致命弱点:杀虫速度慢。最近,一项发表于《农业与食品化学杂志》的研究,揭示了一种颠覆认知的策略——病毒非但不躲避宿主的防御系统,反而“劫持”了宿主细胞内的一个关键“死亡开关”,从而让自己变得更致命。
这个“死亡开关”就是甜菜夜蛾的 Caspase-4。研究团队发现,当病毒(AcMNPV)入侵时,它会激活一个 “Caspase-5 → Caspase-4 → Caspase-1” 的级联反应。其中,Caspase-4并非传统认知的旁观者,而是一个承上启下的 “死亡放大器”。最巧妙的是,科学家们利用这一发现,将Caspase-4的基因插入病毒基因组,制造出一种重组病毒。这种病毒能更高效地启动宿主的凋亡程序,显著加速害虫的死亡,为开发下一代高效、安全的生物杀虫剂开辟了新路径。
下面,让我们通过论文中的核心实验图片,一步步拆解这个精妙的“借刀杀人”战术。
图1:SeCaspase-4的身份鉴定——一个独特的“刽子手”
图片描述(基于原文图1):
A部分:SeCaspase-4的蛋白质序列结构示意图。它包含一个N端的前结构域(Prodomain)和C端由切割位点(ISID, VESD)分隔开的大亚基(Large Subunit, P1)与小亚基(Small Subunit, P2)。大亚基包含底物结合位点(LTHG)和催化活性中心(QACRG),小亚基包含另一个结合位点(RNELYGS)。
B部分:系统进化树。显示SeCaspase-4(标红)与鳞翅目其他昆虫的Caspase-4及果蝇的Damm聚为一支,而与鳞翅目的Caspase-1及果蝇的Drice明显分开。
图片解析:
这张图确立了SeCaspase-4的“身份”。A图 表明它拥有典型的效应Caspase结构,其催化核心非常保守,暗示它具有执行凋亡的功能。B图 的进化树分析则是关键的“身份证”——它明确将SeCaspase-4归类为一个独立于已知效应Caspase-1的独特分支,提示它在功能上可能有别于已知通路,为后续的新发现埋下伏笔。
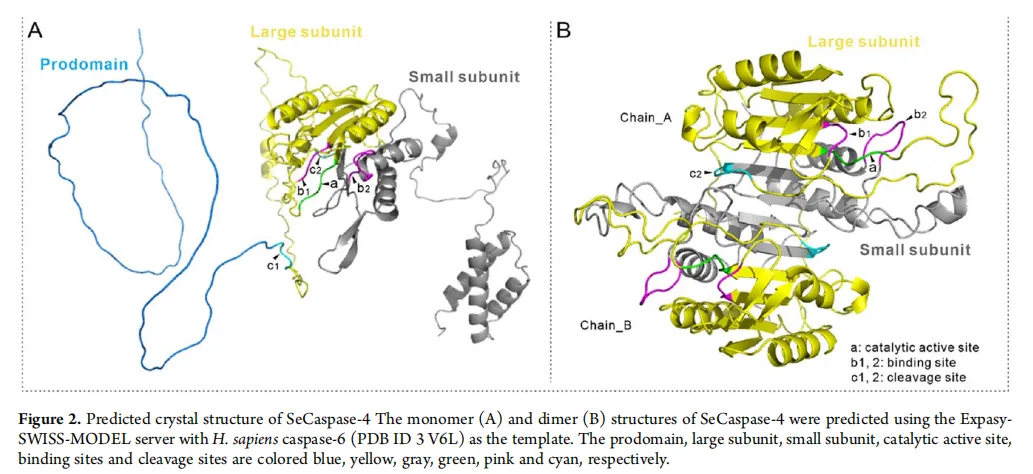
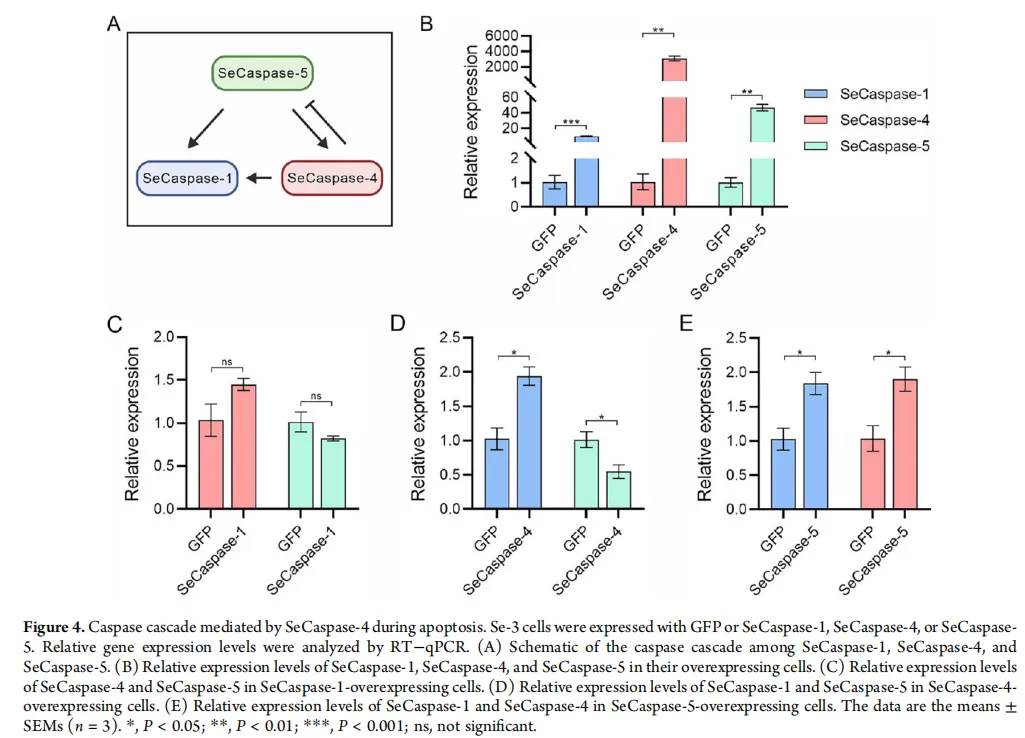
图2:催化核心与作用网络——放大死亡的“引擎”与“线路图”
图片描述(基于原文图2 & 4):
结构预测图:计算机模拟的SeCaspase-4单体(A)和二聚体(B)三维结构。显示其前结构域为柔性环,大、小亚基构成核心催化结构域,活性位点暴露在表面。
级联反应图:一张展示SeCaspase-5、SeCaspase-4和SeCaspase-1三者关系的模式图。箭头指示激活方向:SeCaspase-5可以激活SeCaspase-4和SeCaspase-1;而被激活的SeCaspase-4又可以进一步激活SeCaspase-1。
柱状图:通过定量PCR验证上述级联关系。数据显示,当过表达SeCaspase-5时,SeCaspase-4和SeCaspase-1的mRNA水平均显著上升;当过表达SeCaspase-4时,SeCaspase-1的水平显著上升,而SeCaspase-5的水平下降。
图片解析:
结构图(A, B)直观展示了SeCaspase-4作为功能蛋白的“工作形态”,特别是二聚体形式是其发挥催化活性所必需的。这从结构上支持了它是一个功能完整的效应Caspase。
模式图与柱状图 共同构成了本研究的核心发现。它们清晰地描绘出一个线性的信号放大通路:起始Caspase-5接收到病毒入侵的“死亡信号”后,首先激活SeCaspase-4,后者再将信号传递给另一个效应Caspase-1。这种级联反应就像一个不断放大的警报系统,确保凋亡信号被快速、有效地执行。基因表达数据为这张“线路图”提供了坚实的实验证据。
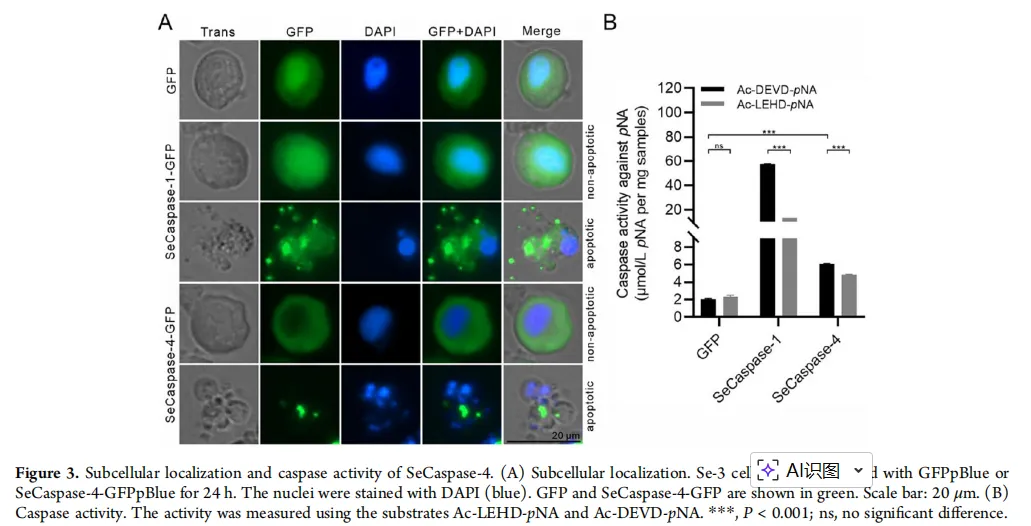
图3:功能验证——谁是真正的“死亡执行者”?
图片描述(基于原文图6):
A部分:一系列SeCaspase-4突变体的结构示意图,包括缺失前结构域(△Pro)、缺失大亚基(△P1)、缺失小亚基(△P2)、同时缺失大小亚基(△P1P2)以及催化关键位点突变(C287A)。
B部分:荧光显微镜照片。显示在昆虫细胞中过表达野生型(WT)SeCaspase-4和△Pro突变体时,细胞出现典型的凋亡形态(细胞皱缩、产生凋亡小体);而过表达其他缺失催化核心(△P1, △P2, △P1P2)或活性位点突变体(C287A)时,细胞形态正常。
C/D部分:蛋白印记(Western Blot)验证各突变体成功表达;柱状图量化显示只有WT和△Pro能诱导约22-24%的细胞凋亡。
图片解析:
这张图通过精妙的“分子手术”(构建突变体),直接揭示了SeCaspase-4执行功能的“核心部件”。结果表明,它的前结构域并非必需(△Pro仍有功能),但大亚基、小亚基以及第287位的半胱氨酸(催化活性中心)对于其诱导凋亡的功能至关重要。一旦这些核心结构被破坏,SeCaspase-4就变成了一把“钝刀”,完全失去了杀伤力。这从分子层面精准定位了其功能域。
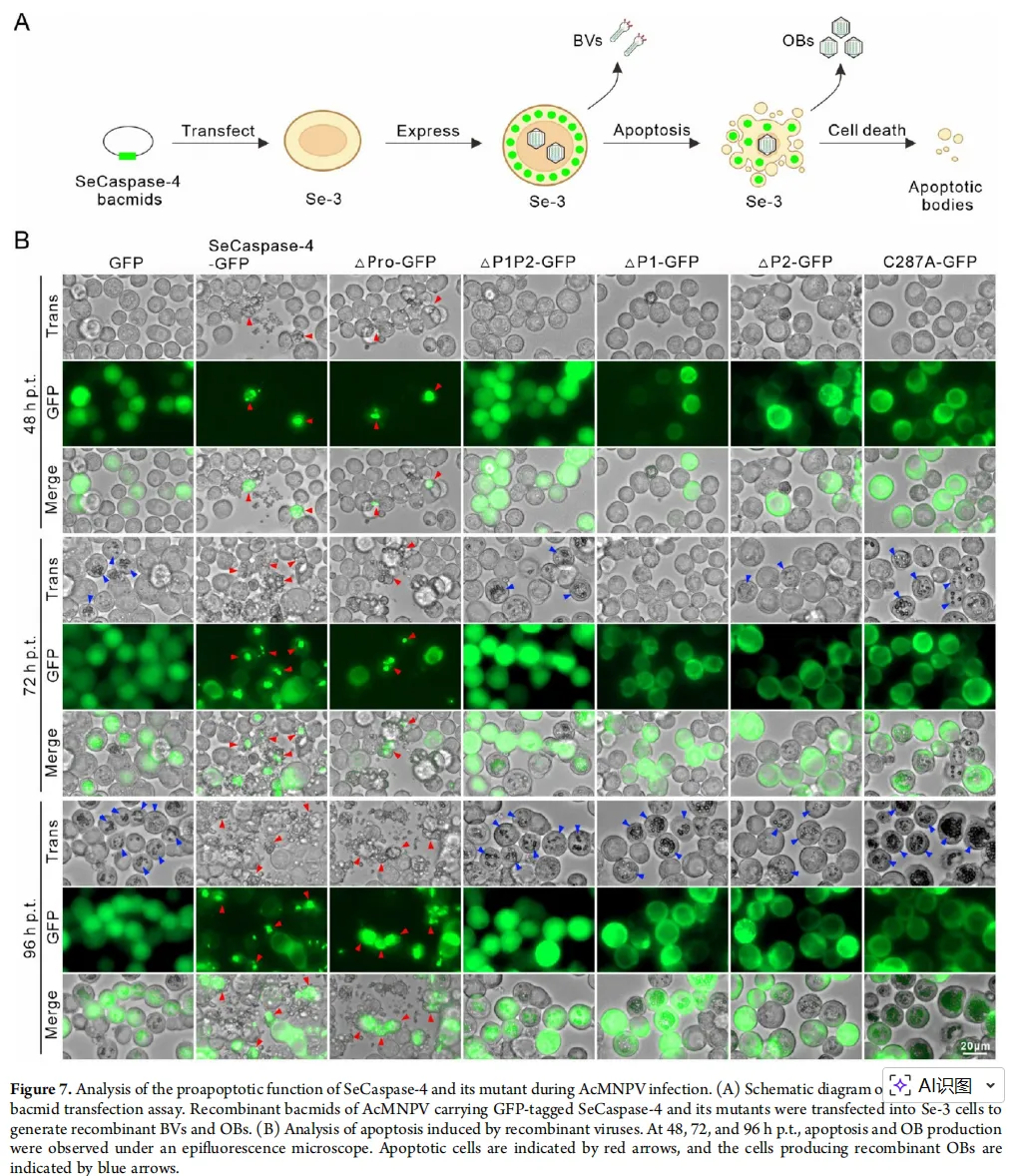
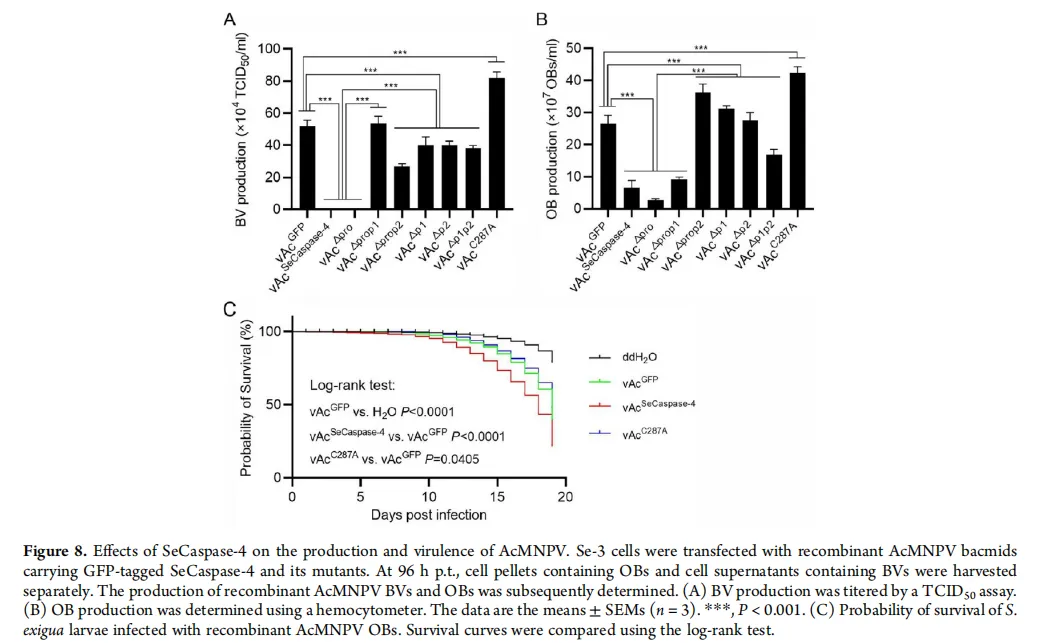
图4:应用效果——改造病毒,加速杀虫
图片描述(基于原文图7 & 8):
A部分(模式图):展示如何将SeCaspase-4及其突变体的基因构建到杆状病毒(AcMNPV)的基因组中,形成重组病毒。
B部分(显微镜照片):比较不同重组病毒感染细胞后的表型。携带野生型SeCaspase-4基因的病毒(vAcSeCaspase-4)感染的细胞,在48-96小时内发生严重凋亡,几乎不产生病毒包含体(OBs, 病毒粒子工厂)。而携带失活突变体(vAcC287A)的病毒感染的细胞,生长正常,并产生大量OBs(蓝色箭头指示)。
C部分(生存曲线):用重组病毒喂食甜菜夜蛾幼虫。结果显示,感染了携带功能性SeCaspase-4病毒(vAcSeCaspase-4)的幼虫,死亡率显著高于感染对照病毒(vAcGFP)或携带失活突变体病毒(vAcC287A)的幼虫。
图片解析:
这是将基础发现推向应用的关键一步。B图 直观地展示了“理论”如何在细胞层面变为“现实”:功能性SeCaspase-4能强力促进病毒感染后的细胞凋亡,从而抑制病毒自身的复制(OBs减少)。
然而,C图 揭示了整个研究最引人深思的“悖论”与创新点:尽管病毒在细胞水平的生产受损,但加速的细胞凋亡(导致宿主组织更快崩溃)却整体上加快了害虫的死亡进程。这意味着,通过“劫持”并增强宿主的凋亡反应,病毒牺牲了部分复制能力,却换来了更强的整体杀虫毒力。这为改良生物农药提供了一个全新的思路:不必一味追求病毒的高复制,转而利用宿主的免疫反应来“借力打力”。
总结与展望
这项研究如同一部精彩的微观世界“谍战片”,完整揭示了病毒如何“策反”宿主的内部安全官(Caspase-4),启动连锁死亡程序,最终达到更高效“斩首”目标(害虫)的目的。
其科学价值在于:
机制突破:首次在鳞翅目昆虫中解析了Caspase-4介导的、不同于经典通路的新型凋亡级联反应(Caspase-5→4→1)。
应用创新:提出了“利用宿主促凋亡因子增强病毒杀虫剂”的全新策略。与以往导入外源毒素基因的方法相比,该方法利用宿主自身基因,潜在环境风险和害虫抗性风险更低,代表了生物农药设计理念的一次重要转变。
未来,基于这一原理,科学家可以进一步优化设计,例如筛选凋亡诱导能力更强的宿主因子,或将其与其他增效策略结合,开发出起效更快、效果更稳定的新一代基因工程病毒杀虫剂,为可持续农业提供更强大的绿色武器。