摘 要:园艺作物香气品质是由挥发性有机化合物构成的重要经济性状,其合成受内部调控网络与外部环境因子的协同调控。茉莉酸、乙烯、生长素等植物激素通过激活MYB和bHLH等转录因子形成复杂的调控网络,并通过交叉互作精确调控萜类和苯丙烷类等香气物质的生物合成。钙离子、过氧化氢和一氧化氮等信号分子通过介导激素信号与环境刺激参与调控香气合成基因的表达。光照和温度等环境因子在园艺作物中不仅独立调控其香气合成,还通过光受体与温度传感器等的相互作用产生协同效应。本文中对近年来植物激素、信号分子及环境因子调控园艺作物香气合成的研究进展进行了综述,旨在为植物花香理论研究和高香气园艺植物的培育提供参考依据。香气物质是植物体内重要的次生代谢产物,是由低分子量、低水溶性和低极性挥发性化合物组成的复杂混合物(Knudsen et al.,2006;Dudareva et al.,2013),赋予园艺作物独特的香气。不同植物的香气物质种类和含量差异明显,形成丰富多样的香气体验(吴琦等,2017;陈雨馨等,2025)。香气挥发物提取物广泛应用于调味料、香水、化妆品制造(Zhao et al.,2017),并可用于缓解精神压力、调节情绪状态(Churchill et al.,2010;Sarid et al.,2016)。相较于园艺植物形态、颜色等性状,香气育种及基因工程改良研究起步较晚(兰熙等,2012)。近年来,随着分子工具的快速发展,香气研究取得突破性进展——挥发物合成通路逐步解析,关键酶基因陆续被鉴定,为“定制香气”提供了可能(赵前等,2025)。香气挥发物生物合成源于初级代谢途径,包括异戊二烯类、聚酮类、生物碱和类黄酮化合物,按合成途径和方式可分为萜类、苯环类/苯丙素类化合物和脂肪酸衍生物三大类(Dudareva et al.,2013;李海燕等,2018;王文静等,2021;Yang et al.,2022a)。香气物质的积累不仅取决于合成途径关键基因,也与园艺作物从发育、成熟到采后全过程中受到的内部激素、信号网络和外部环境因子的动态协同调控密切相关。茉莉酸和乙烯等植物激素可促进香气挥发物合成(刘晓洲,2018;吴琦等,2018),而温度、光照等环境因子通过影响代谢酶活性调控香气释放。值得注意的是,高温可能抑制特定物种香气释放,而传粉者可对花香组分施加选择性压力(Farré-Armengol et al.,2013,2015)。尽管植物香气合成调控已有一定研究,但植物激素、信号分子和环境因子在园艺作物香气合成中的调控机制仍缺乏系统梳理,制约了高香气园艺作物的育种进程。本文中围绕植物激素、信号分子和环境因子在调控园艺作物香气合成中的作用,梳理了香气挥发物中激素介导的调控网络及环境因子影响机制的最新研究进展,以期为高香气园艺作物的品质改良与育种实践提供参考。1 激素与信号分子物质在香气释放中的调控作用
1.1 激素对香气物质代谢的调控
植物挥发性香气物质的合成与释放受内部激素信号与外部环境因子的协同调控。近年来研究表明,茉莉酸、乙烯、生长素等植物激素作为核心内源信号,通过激活或抑制下游MYB、AP2/ERF、ARF等关键转录因子调控萜类和苯丙烷类等挥发性物质的合成(Abbaset al.,2021a;Luo et al.,2021;Wanget al.,2022)。
1.1.1 茉莉酸
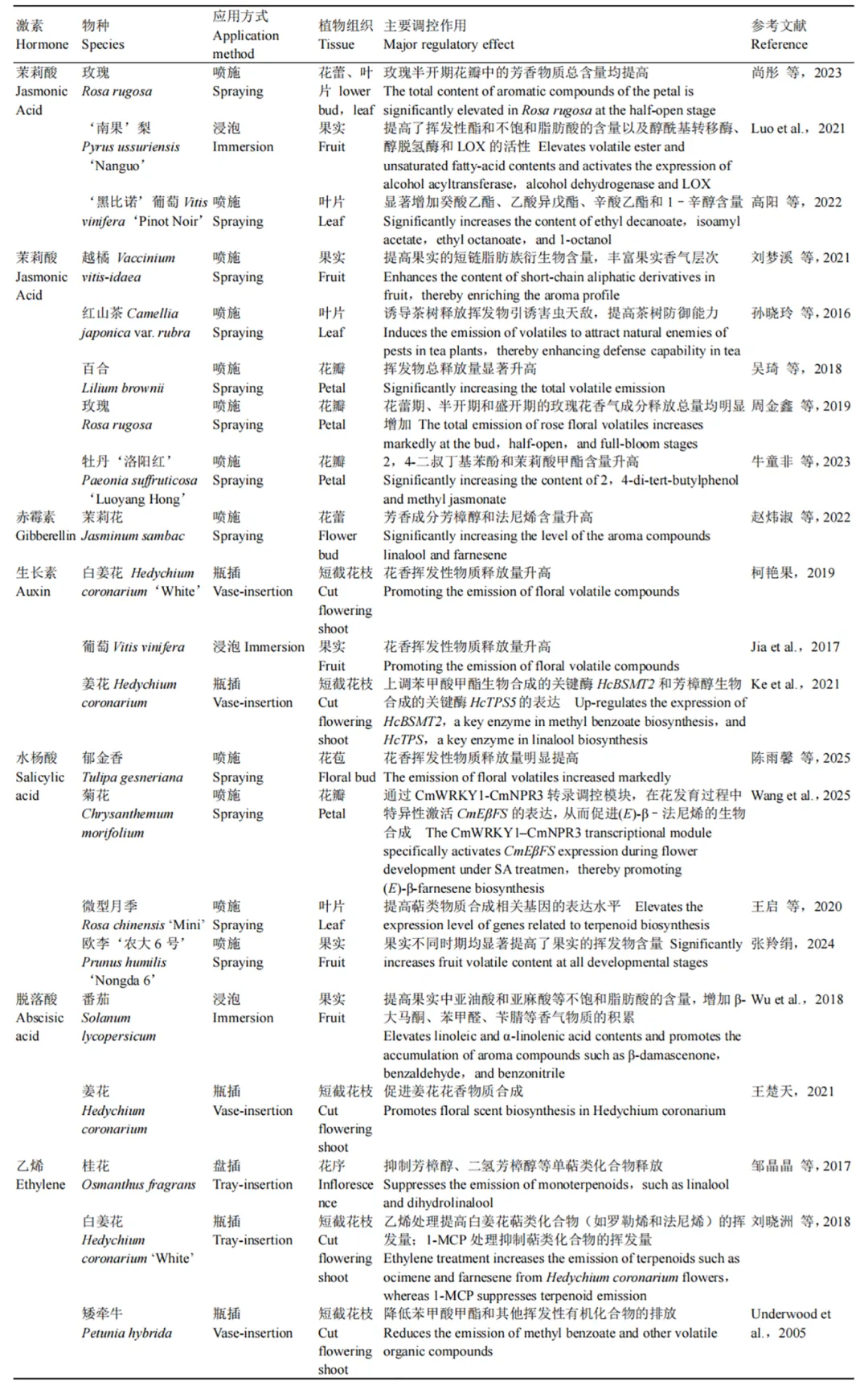
茉莉酸(jasmonic acid,JA)作为一类重要的植物激素,在植物生长发育及防御反应中具有核心调控作用(de Geyter et al.,2012),其衍生物茉莉酸甲酯(methyl jasmonate,MeJA)更被证实是改善香气品质的关键因子(Ayala-Zavala et al.,2005;Kim et al.,2006;Belhadj et al.,2008)。作为最早从茉莉(Jasminum sambac)花中分离出来的挥发性衍生物,MeJA在提升园艺产品香气品质方面表现突出,尤其是在改善水果、蔬菜和花卉的香气品质上起关键作用(Qiu et al.,2009;Alvarez et al.,2010;Chen et al.,2011)。例如,外源喷施MeJA能诱导茶树(Camellia japonica)叶片挥发性物质(香叶烯、法呢烯、苯甲醇等)的合成与释放,如(反)–β–罗勒烯相对释放量达到35%,(反,反)–α–法呢烯达到76.15%(孙晓玲等,2016)。在百合(Lilium spp.)和玫瑰(Rosa rugosa)中,外源喷施MeJA能显著增加萜烯类、醇类等挥发性物质的释放量(吴琦等,2018;周金鑫等,2019)。牛童非等(2023)发现100 μmol · L-1MeJA处理能有效提高牡丹‘洛阳红’的花香物质含量。对于浆果类水果而言,外源茉莉酸不仅能显著提高‘黑比诺’葡萄(Vitis vinifera)中癸酸乙酯、乙酸异戊酯、辛酸乙酯及1–辛醇等关键香气物质的含量,还能提升越橘(Vaccinium vitis-idaea)果实中短链脂肪族衍生物的积累量,丰富果实的香气层次(刘梦溪等,2021;高阳等,2022)。
Keinänen等(2001)发现茉莉酸能够显著提升烟草(Nicotiana tabacum L.)中生物碱、酚类物质及二萜糖苷的积累,这一效应源于外源茉莉酸通过激活茉莉酸生物合成通路及信号转导途径的核心调控基因(如LOX和MYC2),进而调控香气合成酶基因的表达(Hong et al.,2012;尚彤等,2023)。在梨(Pyrus)采后保鲜中,MeJA处理通过上调PuMYB108-like基因表达,有效缓解了冷藏导致的香气物质降解问题(Luo et al.,2021)。MYB9和MYB11在苹果(Malus ×domestica)中参与了茉莉酸诱导的花青素和原花青素生物合成的调节(An et al.,2015),而在草莓(Fragaria × ananassa)中调控C6挥发性化合物的生物合成(Lu et al.,2020,2021)。除MYB转录因子外,其他转录因子家族也参与茉莉酸介导的香气合成调控。在青蒿中,JA诱导AP2/ERF转录因子家族成员ERF1和ERF2的表达从而调控青蒿素的合成(Yu et al.,2012)。在长春花(Catharanthus roseus)中,茉莉酸响应因子ORCA3正向调节类萜吲哚类化合物的生物合成(van der Fits & Memelink,2000)。同样,NaWRKY3和NaWRKY6通过激活JA的生物合成进而提高烟草中的挥发性物质含量,这些挥发物能吸引天敌捕食者从而有效减少了食草性幼虫的数量(Skibbe et al.,2008)。
茉莉酸信号与其他激素信号通路存在复杂的交叉作用。例如,在茉莉酸和赤霉素的诱导下,茉莉酸信号转导途径关键基因MYC2与赤霉素信号通路的负调控因子DELLA蛋白发生互作,通过靶向结合倍半萜合成酶基因TPS21和TPS11启动子的E-box元件,以提高基因表达的方式调控拟南芥(Arabidopsis thaliana)倍半萜的生物合成(Hong et al.,2012)。此外,茉莉酸还与乙烯合成有关。茉莉酸能够通过MdMYC2转录因子与MdACS1启动子上的G-box基序结合,从而诱导乙烯合成(Li et al.,2017)。这一关系在后续研究中得到了证实:无论是MeJA还是MeJA + 1–甲基环丙烯(1-methylcyclopropene,1-MCP)处理,均能显著提高辣椒(Capsicum annuum L.)的内源乙烯水平(Magalhaes et al.,2021)。
这些研究揭示了茉莉酸信号通过MYB等转录因子在植物香气合成中发挥重要调控作用。这种复杂的转录调控网络确保了植物能够根据发育需求和环境变化精确调控香气物质的合成与释放。尽管JA对花香合成的正向调控机制已取得部分进展,但不同物种中JA信号与花香代谢网络的互作模式仍需进一步解析。
1.1.2 乙烯
乙烯(ethylene,ETH)在园艺作物香气物质的释放过程中发挥着关键的调控作用(刘晓洲等,2018)。研究表明,乙烯可通过多种途径影响香气代谢。在矮牵牛(Petunia hybrida)中,乙烯抑制苯甲酸甲酯合成相关基因 PAL 和 BPBT 的表达(Underwood et al.,2005);在番茄中调控果实挥发性物质的释放(McDonald et al., 1996);在番木瓜(Carica papaya L.)中还影响脂肪酸与氨基酸代谢途径,进而促进特定挥发性物质的形成(Balbontin et al., 2007)。乙烯对多种花卉中的挥发性有机化合物(volatile organic compounds,VOC)均具有显著影响,且香味释放水平与切花寿命密切相关(Sexton et al.,2005;Underwood et al.,2005;Borda et al.,2007)。乙烯对萜类化合物的调控呈现双向性:一方面,乙烯能够上调白姜花(Hedychium coronarium)中罗勒烯和法尼烯等萜类化合物的挥发量(刘晓洲等,2018);另一方面,不同花卉种类对乙烯的响应存在显著差异,如乙烯处理后香豌豆(Lathyrus odoratus)和桂花(Osmanthus fragrans)的萜类化合物挥发量下降,而对香石竹(Dianthus caryophyllus)和月季(Rosa chinensis)中的影响不明显(Schade et al.,2001;Sexton et al.,2005;Borda et al.,2011;邹晶晶等,2017)。这些研究表明,乙烯对香气物质释放的调控具有物种特异性,其作用机制可能涉及多条代谢途径的复杂互作。
乙烯通过乙烯信号转导级联反应激活乙烯响应因子(ethylene response factor,ERF),进而精确调控香气物质的生物合成。1-MCP作为乙烯作用抑制剂广泛用于研究乙烯对香气合成的调控。在桃(Prunus persica)中,PpERF5和PpERF7的相互作用上调PpLOX4表达,促进果实香气物质的生物合成(Wanget al.,2022)。在柑橘(Citrus reticulata)中,乙烯通过增强CitAP2.10(AP2/ERF转录因子)的表达,促进了倍半萜(+)-valencene的合成,而1-MCP能够消除这种作用(Shen et al.,2016)。1-MCP不仅降低桃果实中内酯合成相关基因PpFAD1和PpLOX4的表达,延迟内酯产生;还通过破坏LOX、HPL、ADH和AAT等关键酶基因的表达,影响多种香气物质的合成(Cai et al.,2018,2019)。
1.1.3 生长素
生长素(auxin,IAA)显著影响植物挥发性物质的合成与释放。外源生长素处理能够有效提升姜花中萜烯类和苯丙烷类挥发性物质的含量,而生长素信号作用抑制剂对氯苯氧异丁酸(p-chlorophenoxyisobutyric acid,PCIB)处理则会导致花香物质的释放量显著降低(柯艳果等,2019)。Ke等(2021)发现,施用吲哚乙酸(indole-3-acetic acid,IAA)后姜花花瓣中主要花香挥发物罗勒烯、芳樟醇和苯甲酸甲酯的含量分别增加了16%、17%和20%。在葡萄中,生长素处理通过多种途径影响挥发性化合物的积累以及香味相关基因的表达(Jia et al.,2017)。在草莓中,生长素处理增强了2–苯乙醇、苯乙醛和苯甲酸甲酯等酚类挥发物的生成,抑制了苯基氰化物、2–异丁基噻唑、1–己醇和1–硝基–2–苯乙烷的产生。外源生长素处理还改变了与气味挥发物生物合成途径相关的几个关键基因的表达,包括苯甲醛还原酶基因SlPAR1、SLPAR2和SLSAMT1(Wu et al.,2018)。这些研究结果充分表明,生长素在植物挥发性物质的合成与释放过程中发挥着重要的调控作用。
生长素主要通过生长素响应因子(auxin response factor,ARF)家族转录因子发挥作用。拟南芥生长素反应因子AtARF6和AtARF8可以诱导倍半萜的产生(Mandaokar et al.,2006;Reeves et al.,2012)。在姜花中,HcARF5能结合编码β–罗勒烯合成的关键酶基因HcTPS3的启动子,从而参与β–罗勒烯含量的调节。此外,HcARF5还能与关键的气味相关蛋白(HcIAA4、HcIAA6和HcMYB1)相互作用,从而调控花香物质合成(Abbaset al.,2021a)。
生长素与其他植物激素在调控香气合成过程中存在复杂的相互作用。生长素响应因子AtARF6和AtARF8通过激活转录因子AtMYB21和AtMYB24,进而促进茉莉酸的生物合成。这条由ARF6/ARF8启动的、依赖茉莉酸的调控网络,直接促进了花朵中倍半萜类等挥发性物质的合成与释放(Reeves et al.,2012)。在姜花中,褪黑素处理能通过诱导生长素信号转导基因(如HcARF)的表达来提升芳香化合物的水平(Abbas et al.,2021b)。番茄中生长素诱导了苯丙氨酸、异亮氨酸和缬氨酸的积累,而这些挥发性有机化合物的生成能被乙烯所抑制(Tobaruela et al.,2024)。这些激素通过协同或拮抗作用,实现对香气合成途径的精确调控。
值得注意的是,生长素对香气合成的调控具有组织特异性和发育阶段依赖性。在幼嫩组织中,生长素通常表现出对香气合成的促进作用,而在成熟组织中则往往起抑制作用(Medina-Puche et al.,2014,2015)。这种双重调控作用使植物能够根据发育需求,适时调整香气物质的合成与释放。
1.1.4 脱落酸
脱落酸(abscisic acid,ABA)在园艺作物采后香气形成中发挥关键调控作用,其调控范围不仅局限于酯类物质,还可显著影响萜类物质的合成与释放。
在酯类香气物质调控方面,ABA可通过增强合成相关酶活性及生物合成基因的表达,促进短链酯类化合物的积累。例如,外源ABA处理可显著提升猕猴桃(Actinidia spp.)低温贮藏期间果实酯类物质含量(Han et al.,2022);在梨果实采后贮藏过程中,ABA在加速果实软化的同时,还能促进酯类挥发性芳香物质的形成与释放(Xu et al.,2024)。针对‘王林’(Orin)苹果的研究进一步表明,1.9 mmol · L-1ABA能够加速己基丙酸酯、乙基–2–甲基丁酸酯等短链酯的合成,同时上调酯类合成关键基因MdAAT2和MdBCAT1的表达水平;与此同时,ABA可促进乙烯合成基因MdACS1/3和MdACO1的表达,进而诱导乙烯生成,表明ABA可能与乙烯协同正向调控苹果成熟阶段的酯类香气合成(Wang et al.,2018)。
在萜类香气物质调控方面,ABA对萜类合成代谢的调控效应同样显著。10 μmol · L-1ABA处理可显著上调青蒿(Artemisia annua)中AaBPS、AaTPS5、AaBFS等多个萜类合酶(TPS)基因的表达水平,进而促进萜类物质合成(Shu et al.,2025);10 μmol · L-1 ABA与10 μmol · L-1油菜素甾醇(brassinosteroids,BR)联用,可通过调控萜烯类化合物的释放,有效缓解‘阳光玫瑰’(Shine Muscat)葡萄冷藏期间的香气损失(Dong et al.,2025);在400 mg · g-1 ABA处理条件下,葡萄果实中糖苷化香叶醇、橙花醇及β–香茅醇的含量分别显著提升48.7%、84.7%和112.3%,萜类香气物质积累量显著增加(Wang et al.,2022)。
在分子调控机制层面,ABA信号通路可通过靶向调控MYB类转录因子及下游关键合成酶基因,实现对香气物质合成的精准调控。以姜花为例,外源ABA处理可使HcMYB145基因表达水平提升19倍,并进一步诱导HcMYB75、HcMYB79及HcMYB238等基因的表达;其中HcMYB238可直接激活苯甲酸甲酯合成关键基因HcBSMT2的启动子,HcMYB79则能结合萜类合成基因HcTPS10与HcBSMT2的启动子区域,最终促进萜类物质的合成与释放(Abbas et al.,2021b)。此外,外源ABA与JA协同处理还可增强HcbZIP80和HcMYC2-1对萜类合成关键基因HcTPS1启动子的转录激活活性,进而提升1,8–桉油醇等主要花香物质的积累量(王楚天,2021)。
这些发现揭示了ABA在园艺作物香气品质形成中的核心作用,明确了其对酯类、萜类等多类香气物质的调控能力及分子通路,为采后香气品质定向调控和高风味园艺作物育种提供了重要的理论依据。
1.1.5 水杨酸
水杨酸(salicylic acid,SA)对园艺作物萜类物质代谢的调控作用具有显著的物种特异性和时间依赖性。在风轮菜(Satureja hortensis)中,叶面喷施SA处理可显著提高单萜烃含量,但同时导致倍半萜烯和含氧单萜含量降低(Pirbalouti et al.,2014)。这种差异同样体现在丹参(Salvia miltiorrhiza)中,SA处理初期可诱导SmDXR基因表达上调,但随时间推移其表达量呈现下降的趋势(Yan et al.,2009)。这些研究结果揭示了SA在园艺作物萜类代谢调控中的复杂作用机制。
植物香气物质的生物合成是受多因素调控的复杂过程,其中植物激素及其抑制剂的动态平衡发挥着核心调控作用。研究表明,生长素、赤霉素、茉莉酸等内源激素通过激活特定转录因子或代谢酶基因的表达,显著影响了苯丙烷类和萜烯类等香气物质的合成路径(表1、表2)。然而,当前研究对激素信号转导与香气物质积累之间的分子级联反应仍缺乏系统性认知,特别是不同激素间协同或拮抗作用的调控网络尚未完全阐明。
表1 调节植物挥发性香气化合物产生的主要转录因子
(滑动图片查看表1)
表2 植物激素对挥发性化合物合成的调控作用
(滑动图片查看表2)
1.2 信号分子物质调控香气物质生物合成
1.2.1 钙离子
钙离子作为植物细胞内重要的第二信使,在信号转导和胁迫响应中发挥着关键作用(Lecourieux et al.,2002;Dodd et al.,2010)。然而,近年来的研究表明钙调蛋白也是调控香气物质生物合成的关键信号。Asai等(2009)发现香气物质能够诱导拟南芥叶片中细胞质钙离子浓度([Ca2+]cyt)瞬时升高。钙离子浓度的增加与佛手柑(Citrus medica)、薄荷(Mentha canadensis)和柠檬草(Cymbopogon citratus)中倍半萜及挥发性成分含量的提高密切相关(Kothari et al.,1987)。钙离子抑制剂抑制钙离子的流动速率,并显著抑制主要香气物质的释放。如LaCl3和EGTA处理百合后,净Ca2+流入受到抑制,导致单萜类物质释放总量减少约60%;光照刺激能够触发Ca2+内流至细胞质,激活TPS(萜类合成酶)的表达,进而调控单萜的生物合成和释放(Hu et al.,2015)。这一发现揭示了钙信号在环境因子调控香气合成中的传导作用。钙离子在园艺作物采后处理中也发挥着香气调控的作用。研究表明,采用氯化钙处理能够促进苹果果实在贮藏期间香气物质的生物合成(Ortiz et al.,2009)。特别是在苹果的商业成熟期进行该处理,可更显著地提升其贮藏过程中的香味物质含量(Ortiz et al.,2010a,2010b)。这些发现为采后保鲜技术的创新开发提供了新的思路。
1.2.2 过氧化氢
过氧化氢(hydrogen peroxide,H2O2)在萜类化合物的代谢调控中扮演着重要角色。Petrovic等(2025)以荆芥属(Nepeta)的光花荆芥(N. nuda)和大花荆芥(N. grandiflora)为研究材料,通过水涝胁迫(waterlogging stress,W)、H2O2(100 μmol · L-1)及W + H2O2处理48和120 h后发现,3种处理均能有效激活植株H2O2响应信号通路,进而调控萜烯与酚类物质的生物合成过程;其中,光花荆芥和大花荆芥对H2O2处理的响应存在显著差异,光花荆芥关键香气成分荆芥内酯(nepetalactone,NL)含量最高较对照提升12倍,而大花荆芥中1,5,9–表脱氧木香内酯(1,5,9-epi- deoxyloganic acid,1,5,9-eDLA)含量最高仅较对照提升1.5倍;分子研究进一步表明,光花荆芥和大花荆芥的NEPS基因家族成员NnNEPS1/2基因和NgNEPS3基因在短期W或H2O2处理下显著上调,揭示了H2O2介导的胁迫信号调控荆芥属植物香气合成的分子途径。
H2O2对香气合成的调控作用在球根花卉中也得到验证,石雪珺等(2020)在百合上的研究发现,施加苯并噻二唑(2,1,3-benzothiadiazole,BTH)可通过显著抑制百合体内H2O2的积累,进而减少单萜类香气化合物的释放,从反向证实了H2O2在香气合成中的正向调控作用。
综上所述,H2O2可通过激活植株氧化应激反应,调控酚类与萜烯合成通路关键基因的表达,进而影响园艺作物香气相关代谢物的积累,且其调控效应存在物种特异性及与其他信号分子/环境胁迫的协同性。目前,关于H2O2调控园艺作物香气合成的分子网络仍需进一步解析,深入探究其核心调控节点,可为园艺作物香气品质的定向改良提供更精准的理论依据与技术支撑。
1.2.3 一氧化氮
一氧化氮(nitric oxide,NO)作为参与植物发育、激素和环境反应的重要信号分子(Neill et al.,2003),在植物防御反应中发挥着关键调节作用。Souza等(2013)发现,NO和臭氧类似,都能诱导高萜烯(E)-DMNT、芳樟醇和2–丁酮等挥发性有机化合物的排放,但在NO单独处理下,这些化合物的排放水平较低;此外,暴露于O3的植物具有更高的VOC排放量和更低的抗氧化水平,这主要表现为单萜类物质的排放增加以及谷胱甘肽(GSH/GSSG)、谷胱甘肽还原酶(GR)和过氧化氢酶(CAT)活性的降低;这些结果表明NO处理后挥发性有机化合物排放的减少可能是植物抗臭氧胁迫的一个途径。
NO处理在果实采后贮藏中表现出多重生理效应。NO不仅可以通过调节桃果实中脂氧合酶基因(如与C6醛形成相关的PpLOX2、PpLOX3,以及与酯类、内酯形成相关的PpLOX1、PpLOX4)的表达来调控其活性,进而影响果实的贮藏品质,还可以通过抑制乙烯的产生和提高抗氧化酶活性来增强果实在贮藏期间的抗逆性(Zhang et al.,2010,2019;Cai et al.,2020;Zhao et al.,2020,2021)。这一机制在扁桃(Amygdalus communis)中得到印证:NO处理不仅提高了扁桃果实的脂肪酸含量,减轻了冷害症状,还为冷藏期间香气物质的合成提供了充足的前体物质;在低温贮藏中,NO处理增强了乙醇脱氢酶(alcohol dehydrogenase,ADH)和氢过氧化物裂解酶(hydroperoxide lyase,HPL)的活性,有效延缓C6醛、醇和酯等挥发性化合物的损失(Song et al.,2024)。该研究表明了低温处理和一氧化氮处理的联合应用可以有效减少VOC的损失,并有可能抵消单一低温处理在冷藏期间的不利影响,这为扁桃果实采后保鲜技术研究提供了新的思路。2.1 光照对香气物质合成的影响
2.1.1 光强的调控效应及机制
光强通过影响植物光合作用效率及次生代谢通路关键基因的表达,直接调控香气物质的合成总量与组分比例,且多数植物的香气合成呈现明显的光依赖特征。在对草本与球根花卉的研究中,Hu等(2013)发现光照强度从50 μmol · m-2 · s-1提升至200 μmol · m-2· s-1时,百合花香成分总量显著提升47.8%,其中芳樟醇等关键单萜类挥发物含量与光强正相关,明确了光强对单萜合成的促进作用。分子机制层面,百合中LiMYB108转录因子具有光强依赖表达特征,该因子可直接结合萜类合成酶基因LoTPS1启动子的MBS基序,使罗勒烯和芳樟醇合成酶活性提升2.1倍,进而促进目标香气物质的合成积累(Yang et al.,2023),揭示了光强调控香气合成的转录调控路径。光强不足则会显著抑制香气物质合成并诱发不良风味积累。如在黄瓜中,遮荫处理导致光合效率下降,进而影响脂肪酸代谢通路,以(E)-2–壬烯醛为主的不良风味在商业成熟果实中增加1.56倍,糖类和脂肪酸含量分别下降了28%和12%;而1–壬醇、1–己醇等特征醇类香气物质含量显著降低,最终降低了果实的香气品质(Xue et al.,2024)。这一结果表明,适宜的光强是维持作物正常香气代谢的基础。2.1.2 光质调控香气物质合成的特异性
蓝光对单萜类香气物质的合成具有突出的促进作用。Yang等(2022)比较红光(660 nm)、绿光(520 nm)和蓝光(450 nm)对金鱼草(Antirrhinum majus)的处理效果,发现蓝光处理可使单萜含量提升2.3倍,显著优于其他波段。Han等(2022)进一步解析其分子机制,证实蓝光信号关键受体AmCRY1可通过蛋白互作激活转录因子AmMYB24,进而上调AmOCS基因表达,最终使罗勒烯合成量增加40%,明确了蓝光介导的香气合成调控通路。此外,在采后领域,蓝光处理也展现出应用价值,对草莓进行48 h蓝光照射,可显著提升酯类等主要香气VOC的浓度,同时促进花青素积累,实现品质提升(Farneti et al.,2026)。红光可协同调控香气物质合成与果实综合品质。在草莓栽培中,红色薄膜可显著上调FaNES1(法呢烯合成酶)基因表达,定向促进萜类物质的释放(Peng et al.,2020)。在采后保鲜领域,红光处理同样效果显著:4 ℃条件下红光照射鲜切西瓜,可显著减少表面色差并延缓香气劣化(Shi et al.,2020);对草莓进行48 h红光处理,可显著提升酯类香气物质含量,有效减轻提前采收导致的品质下降问题(Farneti et al.,2026)。紫外线(UV-C、UV-A)通过调控脂氧化酶途径促进香气物质合成。研究表明,1.5 kJ · m-2剂量的UV-C照射可提升桃果实中不饱和脂肪酸水平,诱导脂氧化酶通路关键酶的表达与活性,进而促进酯类和内酯类香气物质积累;同时,该处理还能保持果肉紧实度、抑制呼吸速率,维持采后品质(Zhou et al.,2024)。不同光质(波段光谱)调控香气物质合成的特异性为实现香气品质的精准调控提供了可能。2.1.3 复合光的协同与拮抗效应
复合光对香气物质的调控机制较单色光更为复杂,不同光质比例组合可靶向调控不同类型香气物质的合成。Carvalho等(2016)证实蓝/红/绿或蓝/红/黄复合光可显著提高罗勒(Ocimum basilicum)单萜类挥发物产量,而蓝/红/远红光组合则特异性促进倍半萜类物质合成,表明复合光质可通过比例调控实现香气组分的定向优化。任婷婷等(2025)分析红蓝光(1︰4)补光处理可显著促进‘阳光玫瑰’葡萄果实中单萜化合物积累,提升果实香气品质。人工光植物工厂(plant factory with artificial lighting,PFAL)进一步验证了复合光的调控价值。在番茄生产中,以白光为基础,添加高日光积分(DLI)绿光与低DLI红光组合会抑制主要风味物质积累,尤其是绿光可导致与新鲜番茄香气相关的2–异丁基噻唑比例下降(Fan et al.,2023)。2.1.4 光周期对香气释放的时序调控
光周期对香气合成的调控能够通过甲羟戊酸途径实现。Kawoosa等(2010)研究发现,光周期可显著调节胡黄连(Picrorhiza scrophulariiflora)中萜类合成关键基因HMGR(编码3–羟基–3–甲基戊二酰辅酶A还原酶)和DXR(编码1–脱氧–D–木酮糖–5–磷酸还原异构酶)的表达水平,进而显著提升萜类物质含量。昼夜节律系统通过调控香气相关基因的节律性表达,决定香气释放的峰值时间。PhLHY基因作为昼夜节律核心调控因子,可调控矮牵牛(Petunia hybrida)中苯丙素类挥发物相关基因的时序表达,使该类香气物质在午夜达到释放峰值(Boatright et al.,2004)。值得注意的是,昼夜节律与光信号通路存在交叉调控,且内源激素(如乙烯)参与这一过程。乙烯可通过下调PhBSMT1/2等基因转录,显著抑制矮牵牛中花香挥发性苯/苯丙素类化合物(floral volatile benzenoids/phenylpropanoids,FVBP)的合成,表明内源激素与光信号的协同作用是香气时序调控的关键(Colquhoun et al.,2010)。2.2 温度对香气物质合成的影响
环境温度与园艺作物的香气物质合成与释放也密切相关。研究表明,温度升高可显著激活萜烯类化合物合成酶的活性(Kivimäenpää et al.,2022)。光照和高温通过下调VvWRKY70基因的表达促进了葡萄果实中降异戊二烯和黄酮醇积累(Wei et al.,2023)。极端热胁迫则可能通过破坏腺毛结构或改变其表面渗透性,促使脂氧合酶(LOX)途径衍生的挥发性物质及萜类化合物释放。如菊花脑(Chrysanthemum nankingense)叶片在热胁迫下倍半萜含量显著上升,且仅在45 ℃处理条件下特异性检测到单萜樟脑与倍半萜(E)-β–石竹烯的生成(文典,2022)。然而,持续的高温胁迫可能引发光合系统不可逆损伤,进而抑制植物正常生长发育(Hüve et al.,2011)。虽然这些研究表明高温环境与园艺作物香气品质之间存在较强的关联性,但高温环境通常对植物的生长和品质产生负面影响。如何在维持植物生理稳态的同时,又能精准调控园艺作物香气等品质性状仍是当前研究中亟待解决的问题。2.3 其他环境因子对香气物质的影响
除光照外,水分条件、二氧化碳(CO2)浓度和压力及采后条件等其他环境因子也可通过调控植物代谢过程,显著影响香气物质的合成、积累与采后保留。水分条件对香气的调控具有显著的剂量依赖性和物种特异性。在番茄栽培中,适度水分亏缺(最大田间含水能力的65%)可显著提升果实特征香气,同时伴随氨基酸和有机酸总量的增加(Jin et al.,2023);而水分不足会导致以‘黑比诺’为原料酿造的葡萄酒中黑加仑、泥土等香气增强,花香减弱(Ledderhof et al.,2014)。CO2浓度和压力对香气物质合成的影响因作物和处理方式而异。生长阶段的高浓度CO2环境可显著提升草莓果实中乙基己酸酯、芳樟醇等多种香气化合物含量,且浓度越高提升效果越显著(Wang et al.,2004);但采后不同的CO2气压贮藏环境对香气的影响存在差异,如8 kPa CO2可提升覆盆子(Rubus idaeus)的固有风味,而15 kPa CO2则会导致α–离子酮、柠檬烯等风味相关挥发物含量下降,同时增加发酵类异味物质(Islam et al.,2024)。此外,高CO2缺O2环境会改变苹果香气组分,使乙醛、乙酯类含量升高,醋酸酯和醛类含量降低,且这种变化品种间差异明显(Dixon et al.,2001)。采后干燥与储存技术直接影响农产品香气保留。在脱水处理中,红外干燥(infrared drying,ID)和相对湿度对流干燥(relative humidity convective drying,RHCD)可使菠萝切片保持较高香气浓度,而冻干(freeze drying,FD)样品的香气浓度最低(Tiliwa,2023)。在储存环境方面,低氧或动态气调(dynamic controlled atmosphere,DCA)储存可影响梨VOC组成,如提升‘Santa Maria’梨中乙酸酯类含量,但会降低多数梨品种的VOC总浓度(Wendt,2024)。3 激素、环境因子与信号分子的协同调控及互作机制
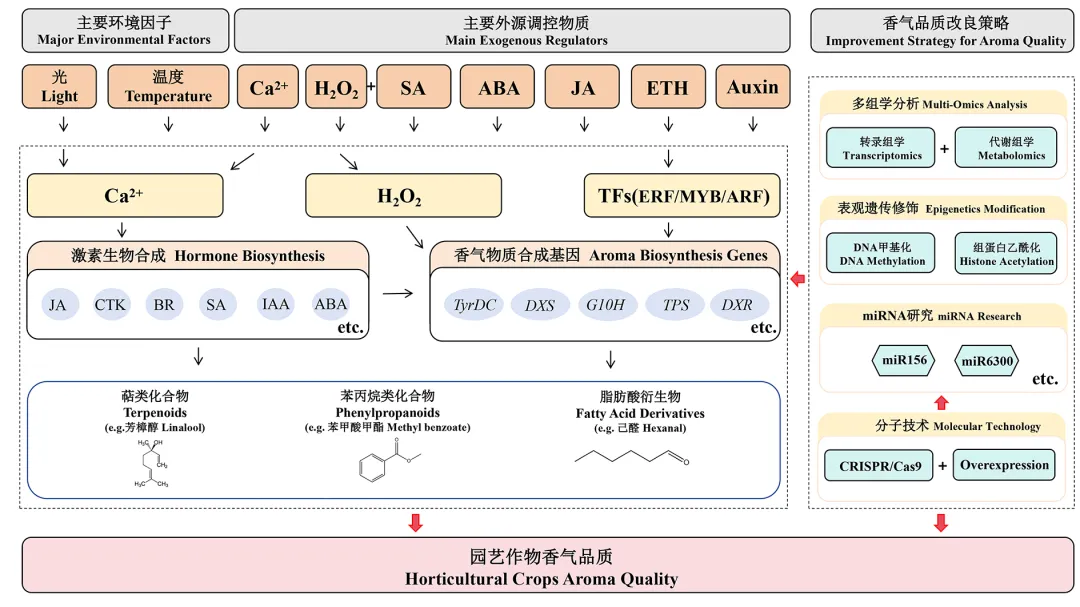
植物香气物质的合成与释放不仅决定植物的生态适应功能(如吸引传粉者、抵御逆境胁迫),也是园艺作物品质的核心评价指标之一。植物激素与多种信号小分子通过交叉作用形成复杂的调控网络,精准调控香气物质的代谢过程。钙离子作为关键第二信使,其动态变化可触发下游激素信号通路的级联反应,进而调控香气相关代谢物的合成。其中钙—茉莉酸(JA)信号模块的保守调控机制已得到证实。例如在金鱼草中,特定光质条件可激活细胞内钙信号通路,进而启动JA信号的级联放大反应,显著上调单萜合成酶基因的表达水平,最终实现对单萜类香气物质合成的精准调控(Yang et al.,2022b)。从分子调控机制来看,钙信号对激素通路的调控依赖于钙依赖蛋白激酶的信号解码作用。在拟南芥中,细胞质内Ca2+浓度的瞬时升高可激活钙依赖蛋白激酶 CPK28,该激酶在生殖生长阶段以负调控方式抑制JA及其代谢物在中心莲座组织的特异性积累,同时间接调控赤霉素(GA)的合成与信号传导,维持JA与GA的拮抗平衡;而在幼苗阶段,CPK28则通过调控活性氧介导的防御信号发挥作用(Matschi et al.,2015)。值得注意的是,该调控通路具有明显的生长阶段和组织特异性,且与防御相关钙信号通路相互独立,不影响JA介导的抗逆防御反应,为解析钙—激素协同调控香气合成的特异性机制提供了重要参考。此外,钙—激素协同调控网络还可通过介导抗性代谢通路,实现抗性诱导与香气品质提升的双重效应。在库德里亚夫泽维氏酵母(Pichia kudriavzevii)诱导樱桃番茄果实抗性中,钙信号与激素信号的激活呈现时序特征:处理早期(0.5和1 h)先激活Ca2+信号相关基因(CML1、CDPK4等)及乙烯信号通路基因(ETR2、EFR1B),处理24 h后进一步激活SA、JA、BR等激素信号通路,并上调苯丙烷生物合成通路中萜类合成相关基因(4CL2、HMG2等)的表达,表明钙信号与激素可通过调控抗性代谢通路,间接实现对果实香气品质的协同改良(Zhou et al.,2025)。钙盐类植物生长调节剂调环酸钙(prohexadione-calcium,Pro-Ca)为通过外源调控激素—钙信号网络改良园艺作物香气品质提供了有效路径。对葡萄叶面喷施不同浓度Pro-Ca,可显著降低果实内源ABA含量,同时提高玉米素(Zeatin,ZT)、赤霉素(GA3)和生长素(IAA)水平,进而促进果实成熟及糖酸代谢相关酶活性提升;其中600 mg · L-1的处理效果最为显著,可使果实中醛、酯、醇、萜烯等7类挥发性化合物的多样性与含量显著提升,较对照增幅达51.46% ~ 423.85%(Li et al.,2024)。过氧化氢(H2O2)与水杨酸(SA)单独或协同作用可通过调控植物特异代谢过程影响香气物质合成,这一调控效应已在甜椒(Capsicum annuum)和细花首冠藤(Castilleja tenuiflora)中得到证实。Jimenez-Garcia等(2018)发现,36 mmol · L-1 H2O2单独处理、0.1 mmol · L-1 SA单独处理及0.1 mmol · L-1SA + 36 mmol · L-1 H2O2混合处理均能显著促进甜椒香气、风味等感官品质变化,且混合处理因二者协同效应使甜椒总体香气强度达到最高;同时,随着H2O2浓度增加,整体香气强度也随之升高,而低浓度组合处理(0.01 mmol · L-1 SA + 18 mmol · L-1 H2O2)则效果最弱;主成分分析显示,36 mmol · L-1 H2O2单独处理与植物性草本香、果香等多种香气高度相关,与SA联用时更偏向调控植物性新鲜风味、辛辣味等特性。Rubio-Rodríguez等(2023)在细花首冠藤中的研究进一步揭示其分子机制,75 μmol · L-1 SA、150 μmol · L-1 H2O2单独及混合诱导均能激活植株氧化应激反应并调控特异代谢,其中混合诱导效果可使总酚含量增加3倍、苯丙氨酸氨裂解酶(PAL)活性提升11.5倍,同时显著上调Cte-TyrDC、Cte-PAL1等酚类合成关键基因及Cte-DXS1、Cte-G10H等萜烯通路基因表达,最终促进苯乙醇、冬青素等香气相关代谢物积累。表明SA与H2O2可通过生理及分子层面的协同调控激活香气合成相关代谢通路,其调控效果优于单独处理,且与处理浓度、植物品种密切相关。光照作为植物生长发育的能量来源与环境信号,可通过光受体感知光质、光强等信号,调控JA、乙烯、ABA等激素的合成与信号传导,最终靶向调控香气合成关键基因的表达,实现对香气物质合成的精准调控。脉冲光作为一种特殊的光照处理方式,可以通过调控激素信号通路影响香气物质保留:3 J · cm-2的脉冲光处理可显著下调甜椒乙烯合成基因(ACO、ACCH、ACS)和ABA生物合成关键基因(NCED1、ABA2、CYP707A)的表达,最终保持甜椒特有的芳香醇、α–蒎烯等,延缓采后品质劣变(包括香气变化)(Jiang et al.,2025)。这一结果表明,人工光照调控技术可通过靶向激素信号通路,实现园艺作物香气品质的精准维持。温度是影响园艺作物生长发育及采后品质的核心环境因子,低温冷藏或高温胁迫均会打破植株体内激素平衡,诱导活性氧(ROS)、钙信号、褪黑素(MT)等信号分子积累,形成激素—信号分子协同响应网络。该网络在调控作物逆境适应性的同时,通过多途径显著影响香气物质的合成与积累,是温度调控园艺作物香气品质的关键机制。低温胁迫下,作物可通过脱落酸(ABA)、MT、ROS等信号分子的协同作用,一方面调控香气合成相关基因的特异性表达,另一方面维持细胞结构完整性,为香气物质的正常合成与积累提供保障。在‘阳光玫瑰’葡萄中,MT可通过调控萜烯生物合成通路、激素信号转导相关基因及香气化合物合成关键基因的表达,有效维持葡萄冷藏期间的固有香气品质(Dong et al.,2025);而冷藏处理会显著改变草莓体内激素代谢水平、ROS积累量及转录因子表达模式,进而影响萜烯类、氨基酸衍生物及苯丙醇类等香气前体物质的合成,最终导致果实香气品质改变(Zheng et al.,2022)。高温胁迫同样会触发激素与信号分子的协同响应,干扰香气物质的合成与积累。在‘阳光玫瑰’葡萄35 ℃高温储存试验中,MT处理可通过调控激素信号转导通路缓解高温引起的细胞损伤,同时维持香气合成相关基因的正常表达,减少特征香气物质流失(Dong et al.,2025);高温处理还会改变草莓体内激素代谢与ROS积累的动态平衡,通过影响萜烯类等关键香气物质的合成过程,导致果实香气品质发生改变(Zheng et al.,2022)。4 结论与展望
园艺作物香气合成的调控机制研究在激素调控、信号分子调控和环境因子调控三个层面均取得了显著进展。在激素调控方面,茉莉酸、乙烯、生长素和脱落酸等植物激素通过复杂的转录调控网络调节香气物质的合成(表2)。茉莉酸诱导MYB转录因子的表达进而调控萜类物质的释放(孙晓玲等,2016);乙烯则主要通过ERF转录因子家族调控香气合成(Li et al.,2017)。不同激素信号通路之间存在复杂的交叉调控网络,例如乙烯可以增强茉莉酸的信号输出,共同促进特定挥发物的合成;此外,乙烯与茉莉酸的协同作用以及生长素与脱落酸在果实发育中的时序性交接,进一步体现了激素网络在调控香气合成时空特异性中的核心作用(Medina-Puche et al.,2014,2015;Magalhaes et al.,2021)。信号分子相关研究揭示了钙离子、活性氧和一氧化氮等在香气合成中的关键作用。钙信号通过钙调蛋白等感受器解码外界刺激,进而调控下游香气合成途径(Asai et al.,2009)。过氧化氢作为重要的信号分子通过调控香气合成基因水平从而增加萜类挥发物释放量(Jimenez-Garcia et al.,2018)。一氧化氮则通过调控挥发性有机化合物的释放进而提高果实贮藏品质(Zhao et al.,2021)。环境因子调控研究显示,光照、温度等环境因素通过多种机制影响香气合成(Han et al.,2022;文典,2022)。这些环境因子之间还存在协同效应,如在胭脂虫栎(Quercus coccifera)中,温度与光照表现出明显的协同作用,控制挥发性有机化合物的排放(Staudt et al.,2011)。尽管园艺作物香气合成与调控研究取得了显著进展,但仍存在诸多挑战。如不同激素信号通路之间的交叉作用机制仍需系统解析,特别是激素信号在转录水平和转录后水平的调控网络,以及不同激素信号在时间和空间维度上的动态变化规律值得深入研究。钙信号、活性氧和一氧化氮等信号分子之间的复杂网络关系尚未完全阐明,这些信号分子如何协同调控下游转录因子和代谢酶活性仍需深入探索。此外,光照和温度等环境因子之间的互作机制尚不明确。近年来,随着多组学技术发展与基因编辑工具革新,花香研究已从单一基因解析迈入系统生物学层面的“全局调控网络构建”阶段,为园艺作物品质精准改良提供了关键策略(表1)。在香气成分调控研究中,代谢组学与转录组学的联合分析成为主流方法(Li et al.,2025;Huang et al.,2025),如野生蔷薇与栽培品种的比较转录组研究显示,MYB转录因子家族成员可通过激活CYP450和TPS基因表达,显著调控萜类香气物质合成(Yang et al.,2025);与此同时,表观遗传学的发展也为花香多样性研究开辟了新维度,其中DNA甲基化差异与香气强度密切相关,相关研究发现,梅花中差异甲基化基因广泛参与花香合成途径(Yuan et al.,2023),组蛋白乙酰化同样调控花香合成基因表达,矮牵牛花中特定香气合成基因在开花后第2天呈高水平组蛋白H3乙酰化,这体现出表观遗传修饰与花香形成时空特异性的高度关联(Patrick et al.,2021);此外,小RNA测序技术鉴定出的一批靶向香气相关基因的miRNA,这类非编码RNA通过介导mRNA降解或翻译抑制参与调控,如miR156、miR6300等分别通过靶向特定基因影响萜类合成,为解析花香形成机制提供了新视角(Yu et al.,2015;Zhao et al.,2025)。现代分子生物学技术的突破为园艺作物香气性状的精准改良提供了有效手段。转基因作为成熟的分子技术,已广泛应用于园艺作物香气品质的改良。Zhang等(2018)在西伯利亚百合中过表达LiDXS和LiDXR基因,显著提高了转基因植株的挥发物水平;Wang等(2001)在辣薄荷(Mentha × piperita)中转入DXR基因,使挥发油产量提高50%。在香气品质改良方面,多基因和多层次的协同调控策略已展现出广阔应用前景。Matsui等(2001)早期利用反义RNA技术抑制康乃馨中F3H基因的表达,有效促进了苯甲酸类花香物质的积累;付琪等(2025)在月季中过表达RcCCD4基因,成功提高了二氢–β–紫罗兰酮的含量;Li等(2025)则通过鉴定出调控丁香酚合成的关键基因SCREP,为香气代谢工程提供了新的靶点。此外,Bednarczyk 等(2025)发现矮牵牛B类MADS-box基因PhDEF能够通过激活MYB转录因子EOB1和EOB2,进而调控苯丙烷类挥发物的合成。这些研究从不同调控层次揭示了香气合成的分子基础,为园艺作物香气品质育种提供了多样化的技术路径与宝贵的基因资源。以CRISPR/Cas9为代表的基因编辑技术通过靶向修饰关键调控基因,已在植物抗性增强、花色改良及花期调控等方面取得系列进展(Shan et al.,2013;Wang et al.,2016;Soyk et al.,2017)。在牵牛花中,Skaliter等(2023)利用CRISPR-Cas9敲除R2R3-MYB转录因子EVER显著提高了苯甲醇、苯甲酸苄酯、苯乙醇等挥发性物质的释放水平。这些进展表明,分子育种技术正推动园艺作物品质改良向精准化与高效化方向发展。尽管在园艺作物香气合成研究中仍然存在诸多挑战,但通过整合多组学分析与基因功能验证,有望构建激素—信号—环境因子协同调控香气的系统模型,为园艺作物香气品质的精准育种提供理论依据。未来可通过整合基因组编辑、表观遗传调控和多组学分析,系统解析激素、信号分子和环境因子与香气合成的协同调控网络,进而通过分子标记辅助选择与基因编辑技术,培育兼具高品质性状与环境适应性的园艺作物新品种。这些分析技术的融合应用不仅将揭示香气多样性的进化机制与生态功能,更将通过技术创新驱动园艺作物产业向功能化、个性化方向发展,满足市场对特色园艺作物品种的多元化需求(图1)。图1 园艺作物香气物质合成与改良研究进展
该研究以中国农业大学园艺学院观赏园艺系,中国农业大学三亚研究院为第一和通信单位。许弘朋为该论文的第一作者。高俊平教授和梁跃副教授为该论文的通信作者。
国家自然科学基金青年项目(32202528);湖北省自然科学基金青年项目(2021CFB132);海南省三亚崖州湾科技城科技创新联合项目(ZDYF2025GXJS144)
阅读推荐:
园艺学报花香专辑|南京林大桂花种质资源创新与利用团队:花香的生物功能及其合成机制研究进展
园艺学报花香专辑|中国农业大学洪波教授团队:菊花花香突变体香气成分鉴定与萜烯合酶基因表达特性分析
园艺学报花香专辑|广东农科院朱根发/杨凤玺团队:腋唇兰花香成分鉴定和关键调控基因挖掘
园艺学报花香专辑|北京林业大学孙明教授团队:‘梨香’菊花香成分鉴定及香气特征分析
园艺学报花香专辑|云南农科院月季创新团队:月季花香物质合成基因及其演化机制研究进展
园艺学报花香专辑|河南科技大学农学院侯小改教授团队:牡丹花香的生物合成及调控研究进展
园艺学报花香专辑|华南农业大学园艺学院范燕萍教授团队:观赏植物花香性状形成及调控机制研究进展
园艺学报花香专辑|北京林业大学孙丽丹教授团队:梅花花香研究进展
园艺学报花香专辑|北京林业大学包菲副教授团队:MYB转录因子调控园艺植物芳香化合物合成研究进展
园艺学报花香专辑|中国科学院植物研究所:香根鸢尾种质资源及其挥发性成分研究进展
园艺学报花香专辑|中国热科院热带观赏植物产业技术创新团队:基于代谢组与转录组联合分析解析秋石斛花香形成机制
园艺学报花香专辑|福建农科院花卉创新团队:山茶花花香生物合成相关基因的实时荧光定量PCR内参基因的筛选及验证
园艺学报花香专辑|扬州大学冯立国教授团队:外源褪黑素对玫瑰花香的影响









 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
