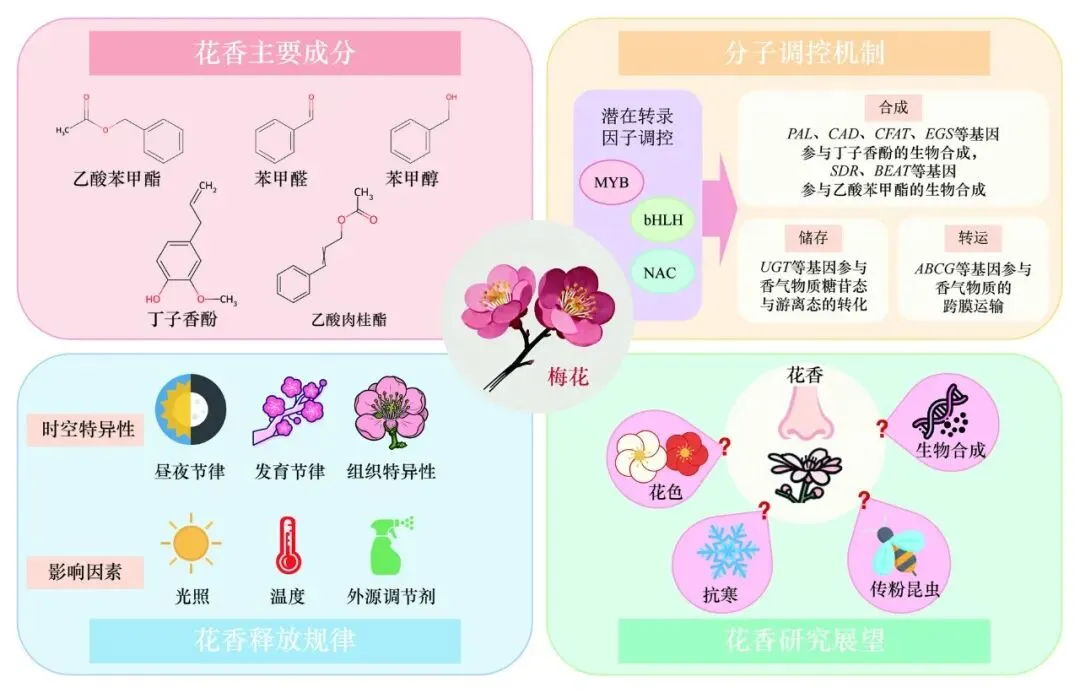
摘 要:梅花(Prunus mume)花香主要由乙酸苯甲酯、苯甲醇、丁子香酚等苯环型/苯丙素类挥发性有机化合物组成,其花香的合成与释放具有时空特异性,同时也会受到光照、温度、外源调节剂等多种因素的影响。本文中综述了近年来梅花花香研究的主要进展,重点总结了花香主要成分及其生物合成途径、分子调控机制和花香释放规律,并对未来研究方向进行了展望,以期为解析梅花花香形成机制提供参考,也为高效培育梅花新异花香品种提供重要研究思路。梅花(Prunus mume)是原产于中国的蔷薇科(Rosaceae)李属(Prunus)落叶小乔木。作为中国传统十大名花,梅花不仅承载着深厚的历史文化内涵,而且兼具重要的观赏价值与经济价值(Zhang et al.,2012,2018)。梅花花香清甜幽雅,香远高洁,作为其重要的园艺观赏性状备受关注与推崇,具有较高的综合开发潜力与广阔的应用前景。植物花香通常由多类挥发性有机化合物共同构成,其中萜烯类、脂肪族化合物以及苯环型/苯丙素类化合物是最主要的组成成分。近年来,以萜烯类化合物为主要香气成分的植物,在香气成分解析、生物合成途径及分子调控机制等方面取得了较为系统的进展(Bao et al.,2023;Li et al.,2024,2025;Shang et al.,2024)。相比之下,梅花花香以苯环型/苯丙素类挥发性化合物为主要特征,其香气组成及形成机制具有明显的物种特异性,但相关研究相对不足。解析苯环型/苯丙素类挥发性化合物的合成途径,挖掘梅花花香形成的遗传调控机制,对于梅花花香研究具有重要意义。本文中系统综述了梅花花香主要成分、分子调控机制、花香释放规律等方面的研究进展,旨在为深入解析梅花特征香气物质的合成与调控网络提供参考,为高效培育梅花新异花香品种提供思路。1 梅花花香主要成分
1.1 梅花特征花香的主要成分
国内外对于梅花花香的研究早期主要集中于不同品种花香成分的鉴定。梅花的分类,主要依据陈俊愉院士在2009年提出的梅花品种群分类新方案(陈俊愉和陈瑞丹,2009),将梅花分为两组三系11个品种群,一组是种内起源的真梅系,另一组是以梅花为亲本杂交而来的种间杂种杏梅系与樱李梅系。真梅系根据枝条、花瓣、花萼等性状分为11个品种群:单瓣品种群(江梅品种群)、宫粉品种群、玉蝶品种群、黄香品种群、绿萼品种群、跳枝品种群(洒金品种群)、朱砂品种群、垂枝品种群、龙游品种群,杏梅系下的杏梅品种群和樱李梅系下的美人品种群。
研究人员分析了不同品种梅花花朵的挥发性物质成分,发现苯环型/苯丙素类化合物种类最多且相对含量最高,乙酸苯甲酯(Benzyl acetate)、苯甲醇(Benzyl alcohol)、苯甲醛(Benzaldehyde)、丁子香酚(Eugenol)、肉桂醇(Cinnamic alcohol)、乙酸肉桂酯(Cinnamyl acetate)等对梅花典型香气形成有重要贡献(王利平 等,2003;金荷仙 等,2005;曹慧 等,2009;赵印泉 等,2010a;郝瑞杰,2014;杨钰 等,2024;Ma et al.,2025;Zhao et al.,2025)。
在鉴定不同品种梅花的挥发性化合物方面,传统的气相色谱—质谱联用技术发挥了重要作用,但化学分析与感官感知之间仍然存在一定的距离。研究者将食品科学中成熟的感官组学方法应用于观赏植物花香评价,基于感官评价将梅花香气分为甜香、清香、花香、果香、草香和无典型梅香6种类型;基于气味活度值筛选出浓香、清香、淡香和甜香4种香型及关键芳香化合物,其中甜香型表现出最高的辨别性和偏好(Zhao et al.,2025)。代谢组学—感官组学整合表明,苯甲醇、丁子香酚和乙酸苯甲酯是造成梅花独特甜香气的特征香气化合物(Zhao et al.,2025)。
目前初步确定了以苯环型/苯丙素类挥发性化合物为主的物质是导致典型梅花香味形成的主要成分(表1)。
表1 梅花花香中主要挥发性有机化合物
1.2 梅花不同品种的花香差异
梅花11个品种群中有些香气表现相似,但也有一些品种群明显不同,甚至无香。赵印泉等(2011a)鉴定了真梅系、杏梅系、樱李梅系共5个梅花品种的香气成分,发现不同品种挥发性化合物种类有明显差异,具有香气的真梅系品种‘青岛朱砂’的挥发性化合物种类数量大于只有微香的杏梅系品种‘淡丰后’,而几无香气的杏梅系品种‘丰后’‘燕杏’和樱李梅系品种‘黑美人’的数量最少;来自真梅系4个不同品种群、2个色系的8个梅花品种中,白色系花香的主要成分是乙酸苯甲酯和丁子香酚,而粉红色系主要成分除这两种物质外还有乙酸肉桂酯、肉桂醇等(Zhang et al.,2020b);Li等(2023)的研究结果也表明红色系的‘骨里红’和‘宫粉’香气更为相似,白色系的‘江梅’和‘绿萼’香气更为相似。这表明不同梅花品种所包含的挥发性化合物成分存在差异,且花香与花色似乎存在一些关联,但目前尚未形成明确且统一的结论。从代谢层面来看,这种潜在关联是具有其生物化学基础的。梅花花色形成相关色素(如黄酮类、花青素类化合物等)与主要的挥发性芳香物质(乙酸苯甲酯、丁子香酚等)在合成过程中均以苯丙氨酸为共同的前体物质,且共享部分苯环型/苯丙素类代谢通路。因此,不同花色的梅花在苯丙氨酸及其产物代谢通量分配上的差异,可能在一定程度上影响其挥发性香气成分的种类与含量。从调控层面来看,花色与花香的关联可能不仅体现在代谢途径的重叠上,还涉及转录调控与代谢调控的协同作用。例如,某些转录因子可能同时调控色素合成基因与香气合成基因的表达,导致色素积累与香气释放呈现出相关性或协同变化,从而在遗传调控层面建立两者之间的联系。然而,目前关于梅花中花色、花香竞争途径与协同调控机制的研究较为有限。未来可通过转录组与代谢组联合分析,筛选同时参与花色与香气合成调控的代谢节点和关键基因,明确花色与花香之间的内在联系,解析梅花不同品种花色、花香代谢的竞争或协同调控机制。
1.3 梅花远缘杂交后代与李属植物的花香差异
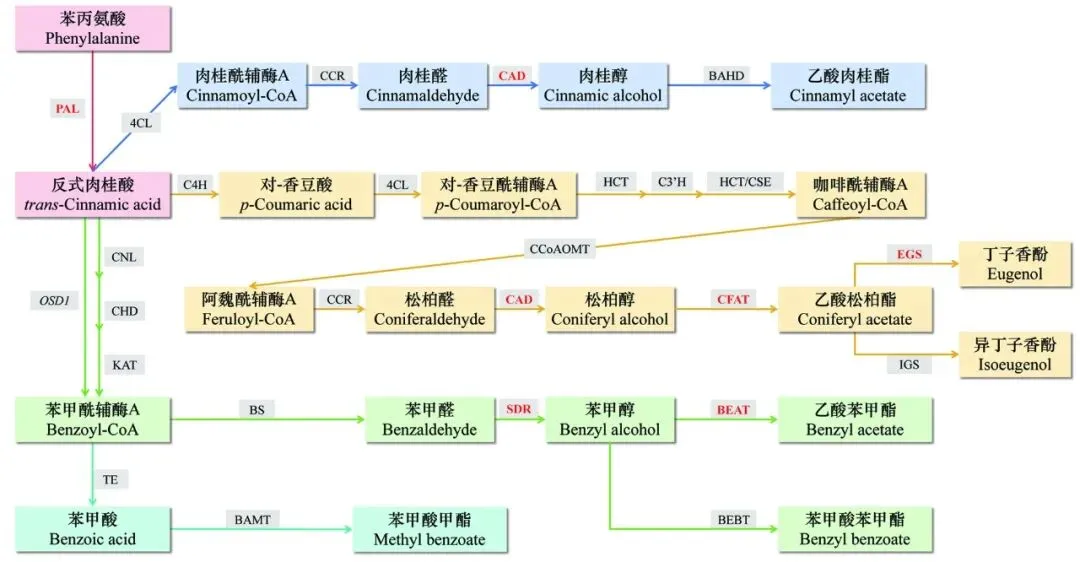
李属植物中,梅花具有区别于其他植物的独特香气。在探究梅花种间香气差异的同时,研究者也关注到了蔷薇科李属植物间香气的异同。赵印泉等(2011a)将梅花品种与近缘种山桃(Prunus davidiana)进行了对比,发现苯环型/苯丙素类化合物是它们的主要挥发物成分;进一步扩大比较范围,测定了杏(Prunus armeniaca)、山杏(Prunus sibirica)、紫叶李(Prunus cerasifera‘Pissardii’)、李(Prunus salicina)、榆叶梅(Prunus triloba)、日本晚樱(Prunus serrulata var. lannesiana)、桃(Prunus persica)、山桃等李属植物的芳香化合物的内源含量和顶空挥发物释放量,明确了乙酸苯甲酯是梅花区别于李属其他植物花香特征的重要成分(郝瑞杰,2014)。真梅系梅花品种与抗寒性强的李属植物(如杏、山杏、紫叶李等)杂交获得的后代出现了抗寒性增强而梅花典型香味丢失的现象。为研究这一问题,郝瑞杰等重点关注了苯甲醛、苯甲醇、乙酸苯甲酯和苯甲酸苯甲酯(Benzyl benzoate)在梅花及远缘杂交子代中的含量,结果表明花香缺失的原因是缺少了乙酸苯甲酯和丁子香酚;通过提供外源醇类底物,发现苯甲醇底物供应不足及存在竞争代谢途径可能是真梅系与其远缘杂交子代香味产生差异的原因(郝瑞杰 等,2014;Hao et al.,2014a,2014b)。绝大部分杏梅系和樱李梅系品种不具有典型的梅花香味,但杏梅系‘淡丰后’与真梅系‘北京玉蝶’的杂交后代‘香瑞白’具有典型梅香,花香性状恢复(陈瑞丹 等,2005)。Bao等(2020)通过比较‘香瑞白’及其亲本的香气成分,结合转录组数据分析,提出杏梅品种花香缺陷的两个主要原因是苯甲酸苯甲酯途径的激活和苯甲醇合成途径的抑制。典型梅花香味主要成分苯环型/苯丙素类物质生物合成途径的研究主要集中在矮牵牛(Petunia hybrida)等草本植物中(Huang et al.,2025),梅花为木本植物,生长周期较长,无稳定转基因体系,在基因功能研究中进行本体验证较为困难,目前多依赖转化模式植物或体外酶活实验确定基因功能。目前,在梅花中,已有部分苯环型/苯丙素类物质合成途径上的关键基因功能得到了初步验证(图1)。图1 梅花中苯环型/苯丙素类物质的生物合成途径
在生成起始阶段,苯丙氨酸经PAL催化生成反式肉桂酸(Zhang et al.,2020a;Wang et al.,2022),并通过4CL、C4H、CNL等参与的一系列酶促反应,生成肉桂酰辅酶A、对–香豆酰辅酶A、苯甲酰辅酶A等不同中间产物(Zhao et al.,2017;Wang et al.,2022),进而生成不同的最终产物;其中肉桂酰辅酶A在CCR作用下生成肉桂醛(Zhang et al.,2022;Wang et al.,2022),肉桂醛经CAD催化生成肉桂醇(Zhang et al.,2022;刘巳塬等,2025),肉桂醇在酰基转移酶作用下生成乙酸肉桂酯;对–香豆酰辅酶A在多步酶促反应下生成松柏醇,PmCFAT1和PmEGS参与了由松柏醇到乙酸松柏酯再到丁子香酚的转化过程(安阳,2016;Zhang et al.,2019),并可能经OMT催化形成甲基丁子香酚(李旭东 等,2013;赵印泉和张启翔,2016);苯甲酰辅酶A可能由梅花中尚未鉴定的PmBSα和PmBSβ合成苯甲醛(Huang et al,2022);短链脱氢酶(如PmSDR108E3/17)可能参与苯甲醛还原为苯甲醇过程(李玉,2024),而苯甲醇又在PmBEAT36/37及PmBAHD3/16/25等酰基转移酶作用下生成乙酸苯甲酯(Bao et al.,2019;Hao et al.,2022a)。但需要注意的是,上述部分关键基因是仅基于已知通路与转录组数据进行关联预测得到的,并未进行严格的体外酶活直接验证。今后有必要在构建梅花瞬时沉默或稳定转基因体系的基础上深入验证上述基因在梅花香气形成中的功能。除结构基因外,转录因子调控也会影响梅花挥发性芳香化合物的产生。结构基因的表达可能会受到MYB、bHLH、NAC等转录因子家族的潜在调控(Zhao et al.,2017;Wang et al.,2022),但在梅花中具体的转录因子–结构基因调控网络尚未清晰。今后需继续深入挖掘调控关键结构基因的转录因子,解析其上下游调控关系,构建梅花花香物质合成的调控网络,完善梅花苯环型/苯丙素类物质合成途径。2.2 花香物质储存
在植物中,芳香物质并非简单地以单一形式存在,而是常以挥发态、游离态和糖苷结合态等形态存在。梅花有大量芳香物质以糖苷结合态的形式存在,其种类与含量也具有明显的时空特异性,随着花朵开放阶段变化;花瓣中糖苷结合态芳香物质种类最多,花药中最少;在糖苷态芳香物质中,又以醛类物质的种类最为丰富(牟晓庆,2022)。在不同开花阶段和部位,苯甲醛糖苷都是最主要的糖苷结合态芳香物质;且苯甲醛糖苷含量随着花朵逐渐盛开而下降,这与梅花花香释放表现相对应,表明苯甲醛逐步向下游芳香物质如苯甲醇、乙酸苯甲酯等转化(牟晓庆,2022)。β–葡萄糖苷酶(β-glucosidase)可以通过水解β-D–葡萄糖苷键参与香气物质糖苷态与游离态的转化,尿苷二磷酸–糖基转移酶(UDP-glycosyltransferase,UGT)是一类能够以尿苷二磷酸–糖为活化糖基供体,将糖基转移至特定受体分子上的酶,PmBGLU18-like,PmUGT71A16和PmUGT73C6可能通过参与上述过程影响梅花香气形成(Wang et al.,2022),PmUGTL9和PmUGTL13在各花期的表达模式与各花期苯甲醛糖苷态含量强相关,可能参与苯甲醛糖苷态与游离态形式的转化(Jia et al.,2024)。2.3 花香物质转运
游离态芳香化合物从植物细胞内到细胞外的运输是一个多步骤、多途径的过程。芳香化合物在植物细胞的细胞质和质体等中合成后以亲脂性的游离形态存在,它们主要通过被动扩散的方式,顺浓度梯度穿越磷脂双分子层构成的细胞膜,从细胞内运输至细胞外。此外,囊泡运输介导的胞吐作用以及ABC转运蛋白参与的主动转运,可能作为补充或特异性调控途径,共同协助化合物向胞外空间的输送。穿过细胞膜后,芳香化合物经由多孔的细胞壁扩散至质外体,并最终抵达如花瓣的表皮细胞或其他特化的分泌结构等释放位点,在这些位点逸散至空气中,最终被人类闻到(Bergman et al.,2025)。芳香植物可以通过渗透团、腺毛体、分泌腔等特化的分泌结构释放芳香物质(Chen et al.,2025),但经扫描电镜观察,梅花花瓣和花丝的表皮结构中未发现特殊的分泌结构(Hao et al.,2021)。挥发性有机化合物通过质膜的过程依赖于蛋白质介导的转运,质膜中的转运蛋白调节挥发性有机化合物的释放(Chen et al.,2025)。ABC转运蛋白(ATP-binding cassette transporter,ABC)是一类广泛存在于生物膜上的“分子泵”,它们可以利用ATP能量,主动跨膜转运包括苯环型/苯丙素类化合物、萜烯类化合物和植物激素在内的多种底物(Adebesin et al.,2017;Ying et al.,2024;Zhou et al.,2024;Li et al.,2025),促进膜内外挥发物浓度的平衡。在梅花中,苯环型/苯丙素类化合物的转运由不同的ABC转运蛋白负责,PmABCG9、PmABCG13和PmABCG23可能参与乙酸苯甲酯的跨膜转运(Hao et al.,2021),PmABCG17可能参与苯甲醛的跨膜转运(Hao et al.,2022b),PmABCG9参与苯甲醇的跨膜转运(郝瑞杰 等,2023)。3 梅花花香释放规律
3.1 香气合成与释放的时空特异性
3.1.1 组织特异性
不同花器官在香气合成与释放中扮演着不同的角色,体现了高度的功能分化。与桂花(Osmanthus fragrans)(Han et al.,2019)、睡莲(Nymphaea prolifera)(Liu et al.,2023)、月季(Rosa gigantea)(Zhou et al.,2024)、兰属(Cymbidium)(汪锦 等,2025)、彗星兰(Angraecum sesquipedale)(Jiang et al.,2024)、有髯鸢尾(Iris × barbata)(孙俊明 等,2025)等香花植物一样,梅花香气物质的合成与释放也具有组织特异性。大多数植物主要通过花瓣组织释放香气,梅花除花瓣外,雄蕊、雌蕊、萼片和蜜腺也都可以释放芳香性挥发物(赵印泉 等,2010b;郝瑞杰 等,2014;Hao et al.,2021;Wang et al.,2022)。梅花不同花器官释放的主要挥发性物质种类不同,如花瓣和雄蕊主要释放苯环型/苯丙素类化合物,而花萼、花盘和雌蕊释放的化合物类型较丰富,除苯环型/苯丙素类化合物外还有萜烯类化合物,如在花萼、花盘和雌蕊中发现释放4种单萜类化合物,α–蒎烯、莰烯、柠檬烯和樟脑(赵印泉 等,2010b)。不同花器官释放的挥发性芳香化合物含量也有很大的差异。花瓣和雄蕊中苯甲醛、乙酸苯甲酯和丁子香酚释放强度较高,花瓣为苯甲醇的主要挥发部位,达到了整体苯甲醇挥发量的56.89%,而乙酸己酯则主要在花萼、花盘和雌蕊群中释放(赵印泉等,2010b;杨姝婷,2021;Hao et al.,2022a);其中乙酸苯甲酯的浓度以雄蕊为中心向四周逐渐减弱,这种浓度梯度可以引导昆虫准确定位花蜜位置(Hao et al.,2021)。作为乙酸苯甲酯挥发中心的雄蕊,各部位的释放效率也不同,花药处含量最高,但挥发量却很低,主要在花丝处挥发(杨姝婷,2021),这可能与花药和花丝的细胞形态与结构、植物高效精准传粉策略有关。此外,梅花花瓣单细胞测序的完成与单细胞图谱的构建,将梅花花香的研究从宏观尺度深入到了细胞尺度。通过鉴别花瓣这一花香主要释放部位的6种主要细胞类型(包括表皮细胞、薄壁细胞、木质部薄壁细胞、韧皮部薄壁细胞、木质部导管和纤维、筛管和伴胞),结合转录组数据分析差异表达基因,确定PmPAL2、PmCAD1、PmBAHD3、PmBAHD5和PmEGS1等是参与梅花花香合成的关键基因,主要在花瓣的表皮细胞、薄壁细胞和大部分维管组织中表达,说明这些细胞可能是合成乙酸苯甲酯和丁子香酚等梅花典型花香物质的主要位点。3.1.2 发育节律
梅花香气物质的释放贯穿于花朵发育的整个过程。在不同开花阶段,关键香气成分呈现规律性的动态变化。挥发物种类的数量在盛花期最多,萜烯类化合物种类由多变少,脂肪酸衍生物则相反,由少变多,而苯环型/苯丙素类化合物种类经历了少—多—少的过程(赵印泉 等,2010b,2011b)。不同开花阶段挥发物的含量也有较大差异,有的呈现高低波动,有的持续增减,如乙酸苯甲酯的释放量在花蕾期较少,在盛花期达到峰值,并随着花瓣的衰老而逐渐减少,总体呈现“低—高—低”的单峰趋势(郝瑞杰 等,2014),苯甲醛则表现为相反的“高—低—高”模式,单萜类α-蒎烯、莰烯含量在开花过程中持续降低,而丁子香酚则不断增加(赵印泉 等,2010b,2011b)。植物为实现传粉效率最大化,通常选择在花朵成熟即将授粉时释放最多的香气物质,之后逐渐减少,这是为了通过气味吸引传粉者的到来;一旦完成授粉,为防止传粉者再次访问造成花朵损害,挥发物的水平也会降低(Shen et al.,2024)。梅花挥发物的释放具有多种调节模式,与花香物质的特定代谢途径相关,但具体的机制尚未探明。不同种类的挥发性物质在梅花中有着不同的释放高峰时段,苯环型/苯丙素类化合物、萜烯类化合物、脂肪酸衍生物、分别在2:00、6:00、14:00达到高峰(赵印泉 等,2011b)。植物通常在特定时段释放挥发性物质作为信号,以匹配传粉昆虫的活动规律。这一花香节律性释放的进化意义在于通过高效利用资源,精准吸引目标传粉媒介,但调控花香释放时间的生物钟机制尚未解。3.2 影响花香的环境因子
花香的形成往往由植物自身遗传特性决定,但众多外界因素也会影响植物芳香物质的合成与挥发,如温度、光照、外源调节剂等。
光照和温度等环境因素可以通过调控相关基因的表达进而影响香气。有研究表明,光照和温度正向调节梅花苯甲醇乙酰转移酶基因PmBEAT的表达,光照条件下PmBEAT的表达量高于黑暗条件下,在一定范围内PmBEAT的表达水平会随着温度的增加而显著增加(Bao et al.,2019)。需要注意的是,基因表达、酶活性与产物生成量之间有时会出现不一致的情况,可能是由于底物含量差异或存在未知的转录后调控。
外源花香调节剂可以通过多种方式调控花香,核心是影响挥发性物质的合成、贮存和释放,具体来说如提供合成所需的前体物质,调节关键酶的活性,影响挥发性化合物的释放,调控植物信号转导等。研究发现,甘草酸、槲皮素、正钒酸钠处理可以在不影响花瓣表皮细胞形态及细胞活力的前提下,显著降低花香梅花挥发物的总量(邱晨 等,2022)。但梅花中的相关研究较少,目前生产中已确定有效的花香调节剂种类也较少,机制也尚未阐明,未来可从外源调节剂种类筛选、调节剂影响花香的生理与分子机制解析等方面开展相关研究。
近年来,关于梅花花香的研究已在不同品种群香气成分鉴定与香型分类、合成通路部分关键基因的功能验证等方面取得了一定进展(图2),为今后继续开展花香研究提供了基础。然而,从整体上看,梅花花香相关研究仍处于起步阶段,许多关键科学问题尚未得到系统阐明。在遗传进化层面,梅花所具有的独特花香并非蔷薇科李属植物的共有特征,该芳香性状的起源与演化机制尚不清晰。已有研究表明,李属多数物种花香较弱,而梅花在苯环型/苯丙素类挥发物的合成与释放方面表现出显著增强的特性,这可能与其经历了关键基因的获得、扩张或功能分化有关。目前蔷薇科李属多种代表性物种及部分梅花品种已完成高质量基因组测序,这为开展跨物种比较和群体层面的进化研究奠定了基础。未来有望通过蔷薇科李属植物的泛基因组分析深入阐明植物芳香性状“从无到有”的起源与演化机制。图2 梅花花香的主要研究方向
在代谢途径层面,苯环型/苯丙素类挥发性有机化合物的生物合成通路尚未完全打通,催化关键反应的酶尚未全部明晰。梅花花香以苯环型/苯丙素类化合物为主,可以成为解析该类芳香物质生物合成的理想研究材料。在今后的研究中可利用梅花丰富的遗传资源,根据不同品种苯环型/苯丙素类化合物含量差异显著的特点,通过性状关联分析锁定关键代谢节点基因,进一步完善苯环型/苯丙素类挥发性有机化合物的代谢网络。在调控机制方面,关于转录因子参与梅花苯环型/苯丙素类芳香物质合成调控的研究仍较为有限。今后需通过多组学手段,持续挖掘调控苯环型/苯丙素类物质合成关键结构基因的核心转录因子,解析其上下游调控关系与协同作用机制。除传统的转录调控外,未来研究还应拓展至更多层次的调控维度。例如,非编码RNA可能通过转录后调控精细调节关键酶基因的表达水平,影响芳香化合物的合成;转座子插入或重排引发的结构变异可能通过影响基因拷贝数、顺式作用元件或染色质构象,导致芳香性状的获得与丢失;DNA甲基化、组蛋白修饰等表观遗传调控机制也可能在花香合成的时空特异性调控中发挥重要作用。在性状互作方面,梅花花香并非孤立性状,其与花色、花发育、抗逆性、环境因子及传粉媒介之间可能存在复杂的关系,有待系统深入研究。阐明梅花花香与花色两类代谢途径的关系,有助于理解植物代谢通量的分配机制,为同时改良花色与花香性状提供理论依据。解析梅花花香合成与抗寒响应在代谢与调控层面的交叉节点,有助于理解植物在复杂生态环境中的适应与进化策略。通过整合苯环型/苯丙素类挥发物合成与逆境响应、发育调控之间的耦合机制,最终有望为定向培育兼具优良花色、浓郁花香与较强抗寒能力的梅花新品种提供理论基础。在育种应用层面,由于梅花童期漫长,传统杂交育种效率低,因此培育市场需求量大的抗寒芳香梅花新品种迫在眉睫,但相关育种工作仍未取得实质性突破,现有品种在数量、类型及综合性状表现上均难以满足产业发展的需要。其核心瓶颈一方面在于梅花种质资源遗传背景不清、性状评价不系统,限制了其在育种中的高效利用;另一方面在于缺少梅花高效稳定遗传转化体系,限制了性状的精准改良。因此,厘清梅花种质资源的遗传基础,建立系统评价体系与高效稳定遗传转化体系,有助于抗寒芳香梅花新品种培育实现突破,也将推动梅花育种由传统育种转向分子设计育种。近年来,人工智能(artificial intelligence,AI)技术的快速发展与纳米技术的应用有望为梅花花香育种提供新的解决思路与技术路径,在梅花花香的基因筛选、表型预测与分子育种等方面发挥重要作用。通过整合基因组、转录组、代谢组、蛋白组及表型组等多组学数据,利用机器学习与深度学习算法,明确梅花花香在李属植物中的起源进化过程,挖掘调控花香合成、贮存、转运的关键基因,完善梅花香气形成的调控网络。通过构建基于深度神经网络的香型预测模型,在早期育种阶段预测杂交后代的花香类型,缩短育种周期;结合强化学习与模拟优化算法,实现虚拟杂交组合的优选与香气改良方案的智能决策。利用纳米载体高效递送外源基因、siRNA及CRISPR/Cas基因编辑系统,突破梅花等木本植物基因转化效率低与再生困难等瓶颈,建立梅花高效稳定遗传转化体系,结合性状标记基因早期筛选,实现梅花新异花香品种的创新育种,推动梅花花香产业化的高质量发展。该研究以北京林业大学为第一和通信单位。博士研究生岳芝伊为该综述的第一作者。孙丽丹教授为该综述的通信作者。

第一作者 岳芝伊
北京林业大学园林学院孙丽丹教授团队围绕梅花花香、花色、花型和抗寒等性状,致力于种质创新、性状解析、新品种培育和高效繁育栽培等核心技术研发。先后主持参与完成国家高层次人才特殊支持计划、青年拔尖人才计划、国家自然科学基金青年和面上项目、“十三五”国家重点研发计划等国家级、省部级科研项目10余项;获国家科学技术进步奖、教育部科技进步奖和教育部自然科学奖等国家级和省部级奖励9项;发表SCI收录论文62篇,以第一作者和通讯作者在Nature Communications、Trends in Plant Science和Plant Biotechnology Journal等高水平期刊发表论文36篇;参编中文专著2部,英文专著3部;参与制定国家林业行业标准2项;获得国家发明专利授权5项,国际专利1项。
团队照片
中央高校基本科研业务费专项资金资助(QNTD202503)
阅读推荐:
园艺学报花香专辑|南京林大桂花种质资源创新与利用团队:花香的生物功能及其合成机制研究进展
园艺学报花香专辑|中国农业大学洪波教授团队:菊花花香突变体香气成分鉴定与萜烯合酶基因表达特性分析
园艺学报花香专辑|广东农科院朱根发/杨凤玺团队:腋唇兰花香成分鉴定和关键调控基因挖掘
园艺学报花香专辑|北京林业大学孙明教授团队:‘梨香’菊花香成分鉴定及香气特征分析
园艺学报花香专辑|云南农科院月季创新团队:月季花香物质合成基因及其演化机制研究进展
园艺学报花香专辑|河南科技大学农学院侯小改教授团队:牡丹花香的生物合成及调控研究进展
园艺学报花香专辑|华南农业大学园艺学院范燕萍教授团队:观赏植物花香性状形成及调控机制研究进展