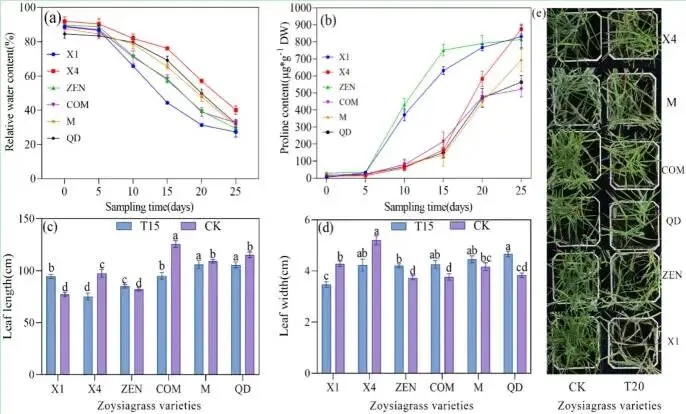
生理指标变化: 随干旱胁迫时间延长,叶片相对含水量(RWC)持续下降,脯氨酸含量稳步增加。
表型性状差异: 干旱处理15天后,不同品种表型出现显著差异;至20天时,X1品种萎蔫最为严重。
耐旱综合评价: 基于生理指标变化趋势,初步确立了六个结缕草品种的耐旱性排序为:X4 > M > COM > QD > ZEN > X1。
在干旱胁迫期间,不同结缕草品种的响应存在显著差异。随着干旱胁迫时间的延长,所有品种的叶片相对含水量(RWC)均持续下降,其中X1品种下降迅速,而X4品种则表现出更强的持水能力。相反,叶片脯氨酸含量则稳步上升,M、QD和X4品种的脯氨酸在第15天开始急剧增加。表型上,干旱处理15天后各品种间出现显著差异,至第20天,X1、M和ZEN品种的叶片开始萎蔫,X1品种尤为严重。Pearson相关性分析显示,RWC与脯氨酸含量呈极显著负相关(P < 0.01)。单因素方差分析表明,叶长、叶宽、RWC和脯氨酸含量在不同耐旱性品种间均存在极显著差异(P < 0.01),可作为评估结缕草耐旱性的可靠指标。基于此,初步的耐旱性排序为X4 > M > COM > QD > ZEN > X1。(Fig. 1)
图1. 结缕草的生理指标与表型生长。 a 结缕草叶片的相对含水量。b 结缕草叶片的脯氨酸含量。c, d 干旱胁迫处理15天后表型性状的比较。e 干旱胁迫处理20天后表型性状的比较。T15,结缕草在干旱胁迫处理下生长15天。T20,结缕草在干旱胁迫处理下生长20天。CK,对照组。同一颜色柱状图中,标有相同字母表示无显著差异(P > 0.05),标有相邻字母表示存在显著差异(P < 0.05),标有分离的字母表示存在显著差异。误差棒代表平均值的标准误差。
3.2 干旱响应基因的鉴定
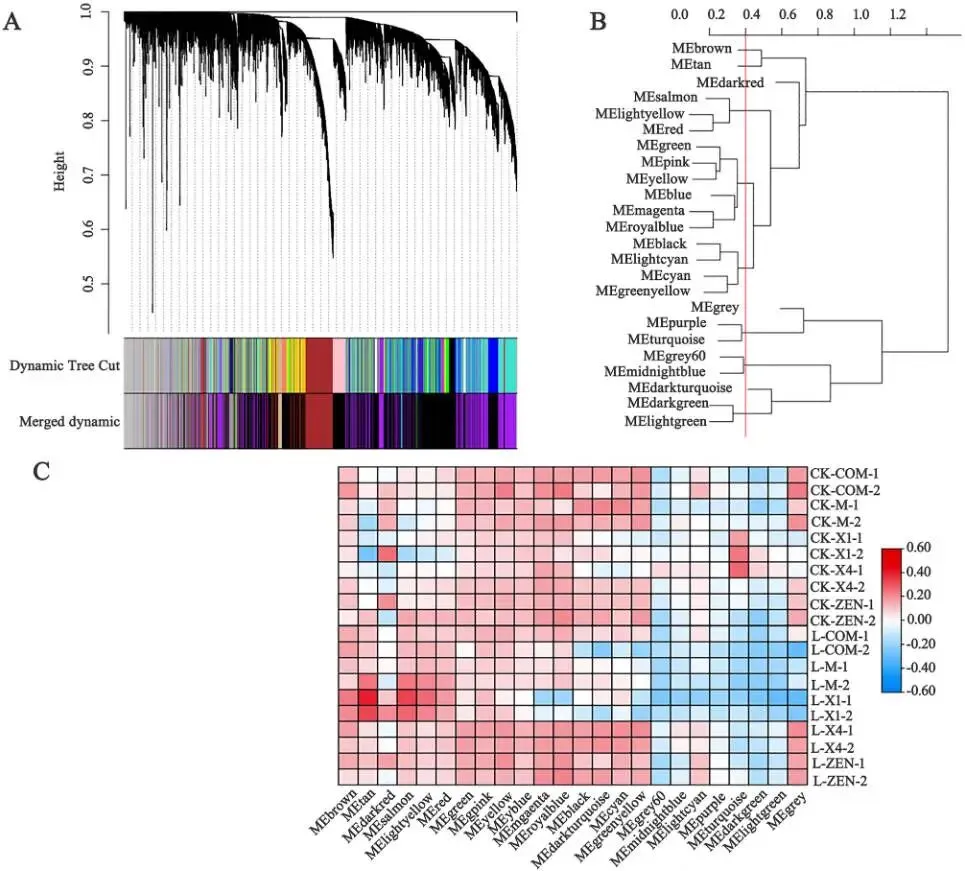
基因共表达分析: 通过基因共表达网络分析将基因划分为24个模块,其中四个模块(MEbrown, MEtan, MEsalmon, MElightyellow)在干旱处理后表现出富集调控。
多组学整合分析: 转录组学与表观基因组学整合分析显示,ATAC-seq峰强度与基因表达水平呈正相关。
候选基因的筛选: 结合转录组差异表达基因与ATAC-seq差异调控峰,最终鉴定出63个与干旱胁迫相关的高置信度基因。
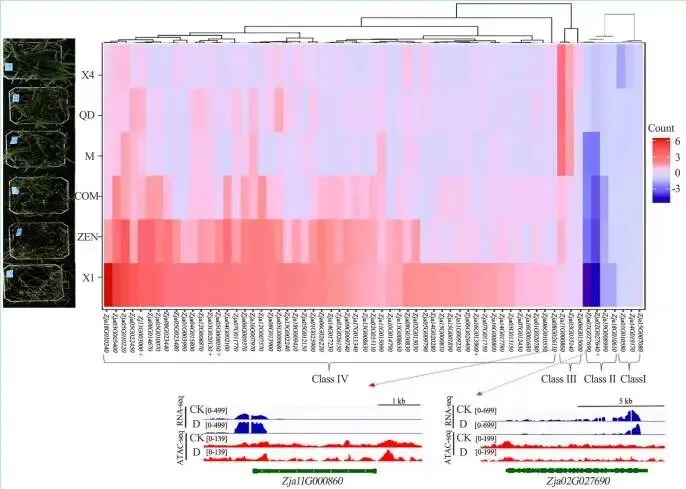
候选基因的验证: RT-qPCR结果与转录组数据的表达趋势基本一致;同源比对发现Zja03G031540和Zja11G000860与已知的关键抗旱基因高度同源,且其在不同品种间的差异表达模式与表型一致。
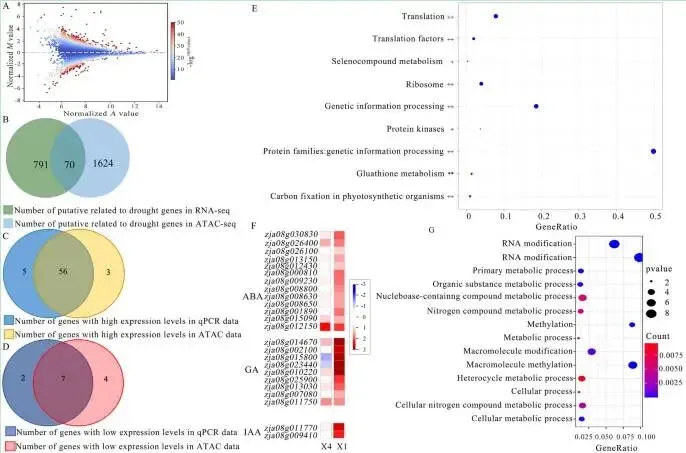
为进一步探索干旱响应基因,通过基因共表达网络分析构建了包含24个模块的进化树和表达热图,发现MEbrown、MEtan、MEsalmon和MElightyellow基因模块在干旱处理后受到富集调控。GO分析表明,这些模块富集在“细胞对缺水的响应”、“极长链脂肪酸代谢过程”等与干旱胁迫高度相关的生物学过程中。整合转录组学和表观基因组学分析发现,ATAC-seq峰强度与基因表达水平呈正相关,例如,基因Zja11G000860在干旱胁迫下高表达,其ATAC-seq峰强度也高于对照组,而Zja02G027690的表达则受到抑制,其峰强度也相应较低。通过对ATAC-seq数据进行差异峰分析,共鉴定出1694个受差异峰调控且与干旱处理相关的基因。将转录组鉴定的干旱响应基因与ATAC-seq数据进行联合分析,最终筛选出63个高置信度的干旱胁迫相关基因。RT-qPCR验证结果与转录组数据的表达趋势基本一致。同源性分析显示,结缕草基因Zja03G031540与SnRK2.3基因高度同源,而Zja11G000860与NAC2和NAC5基因高度同源。这两个基因在耐旱性强的X4、M和QD品种中显著上调,而在耐旱性弱的COM、ZEN和X1品种中显著下调,其表达模式与表型结果相符,表明它们是与结缕草干旱胁迫相关的候选基因。(Fig. 2, Fig. 3, Fig. 4)
图2. 结缕草中的表达基因。 A 结缕草基因表达与聚类。B 模块进化树分析。C 模块的基因共表达热图。‘D’代表干旱处理,‘X4’代表X4结缕草品种,‘1’和‘2’代表重复处理的编号,‘D-X4-1’代表X4结缕草的干旱处理‘1’。其他数字含义相同。
图3. 结缕草中干旱响应候选基因的表达。 CK为实验对照组,D为干旱处理组,红色+标记为扩张基因标记,蓝色+标记为正选择基因标记。术语‘0–139,’ ‘0–199,’ ‘0–499,’ 和‘0–699’指FPKM值,用于表示基因表达水平的范围(彩图见线上版本)。
图4. 结缕草的差异表达基因分析。 A 结缕草的差异峰分析。B 结缕草中的干旱相关基因。C和D 结缕草中的干旱相关基因。E *表示I类基因富集的通路,**表示II类基因富集的通路,+表示III类基因富集的通路,++表示IV类基因富集的通路。F 结缕草的KEGG分析。G 结缕草的GO分析。
3.3 耐旱候选基因的功能分析
KEGG通路富集: 候选基因在遗传信息处理、翻译、核糖体及线粒体生物合成等通路中富集;同时,不同类别的基因分别富集于谷胱甘肽代谢、植物激素(ABA, GA, IAA)信号通路等。
GO功能富集: GO富集分析显示,这些候选基因主要富集在生物过程类别的代谢过程和甲基化功能中。
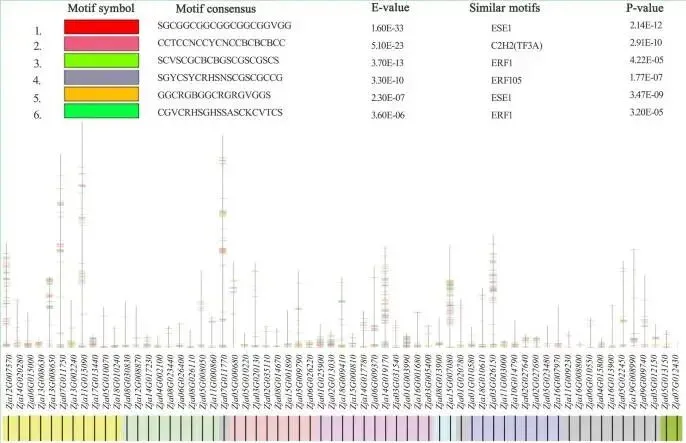
调控基序分析: 鉴定出六个高置信度基因的基序,这些基序与已发表的ERF和C2H2转录因子家族相似,并显著富集于差异调控峰上。
对候选基因的功能分析显示,KEGG富集分析表明这些基因富集于遗传信息处理、翻译、核糖体和线粒体生物发生等通路,这些对于植物在胁迫下调节和适应环境变化所需的蛋白质合成至关重要。其中,II类扩张基因Zja02G027690和Zja02G027640富集于谷胱甘肽代谢通路。III类基因Zja03G031540和Zja11G000860富集于硒化合物代谢和蛋白激酶通路,表明结缕草通过调节应激反应和抗氧化防御来应对环境胁迫。IV类基因则分别富集于脱落酸(ABA)、赤霉素(GA)和吲哚乙酸(IAA)通路,这些基因在不同品种间的表达差异与其耐旱性相关。GO富集分析进一步揭示,这些候选基因主要富集于代谢过程和甲基化功能。此外,通过基序分析鉴定出六个高置信度基因的 motifs,这些基序与已发表的ERF和C2H2转录因子家族相似,并且在37个差异调控峰上显著富集,表明它们参与了这些差异峰的调控。(Fig. 5)
图5. 结缕草干旱响应候选基因的基序分析。
3.4 通过酵母过表达系统观察干旱胁迫响应
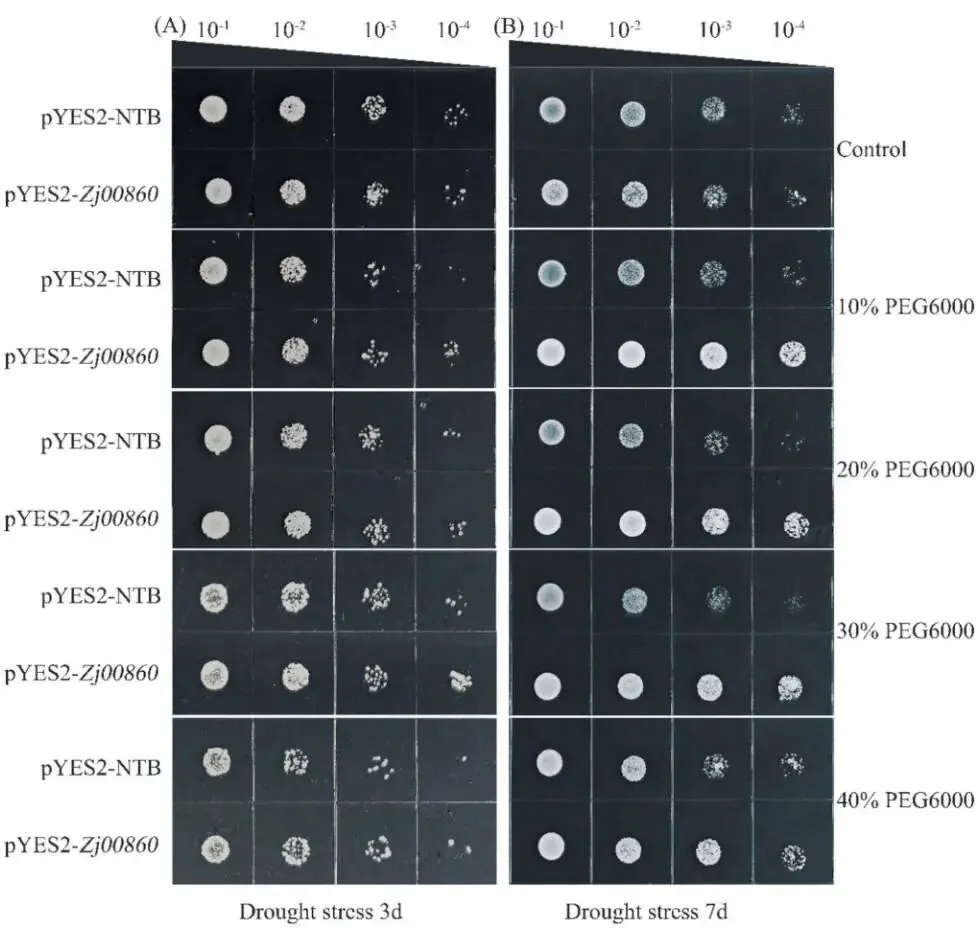
胁迫初期响应: 在干旱胁迫3天后,与对照组相比,菌落生长受到显著抑制,但两个菌株之间无明显差异。
持续胁迫响应: 胁迫持续7天后,两个菌株的生长模式出现分化。
基因功能验证: 转基因酵母菌株Zja11G000860-pYES2即使在40%的高浓度PEG6000下仍能维持稳健生长,表明Zja11G000860基因有助于转基因酵母在干旱胁迫下的生长。
为了验证候选基因的功能,在酵母过表达系统中观察了其对干旱胁迫的响应。在暴露于干旱胁迫3天后,与对照组相比,菌落生长受到显著抑制,但在不同PEG浓度梯度下,两个菌株之间未观察到显著差异。然而,当胁迫持续7天后,两个菌株在不同PEG浓度梯度下的生长模式出现分化。值得注意的是,表达Zja11G000860的转基因酵母菌株(Zja11G000860-pYES2)即使在40% (w/v) 的高浓度PEG6000下仍能维持稳健生长,其菌落计数相对于对照持续增加。这种增强的胁迫耐受性变得愈发明显,表明Zja11G000860基因在促进转基因酵母于干旱胁迫下生长方面发挥了作用。(Fig. 6)
图6. 在酵母表达系统中对Zja11G000860-pYES2耐旱性的分析。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
