MPP | 中国林业科学研究院:过表达花青素还原酶通过增加黄酮类化合物含量抵御油桐木质部镰刀菌枯萎病
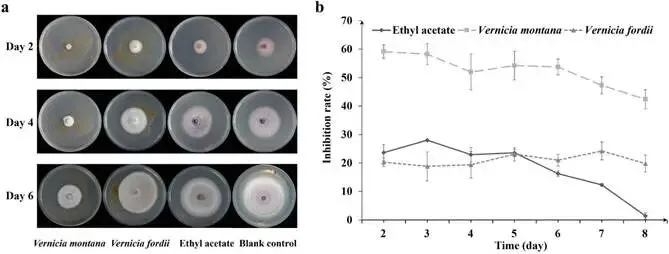
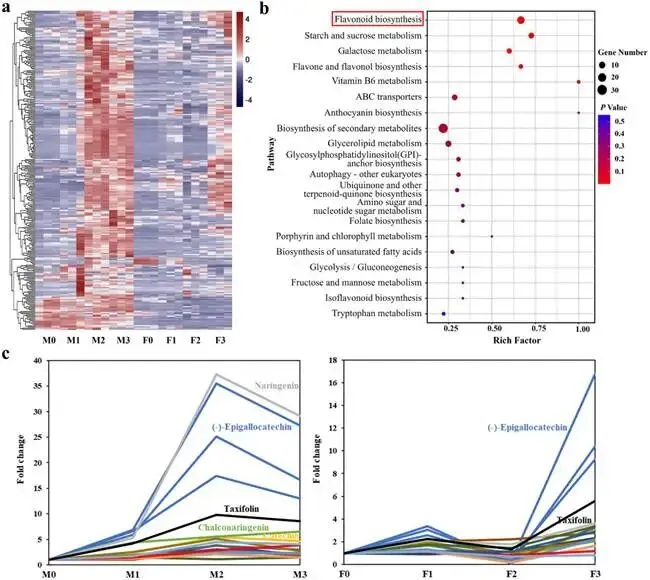
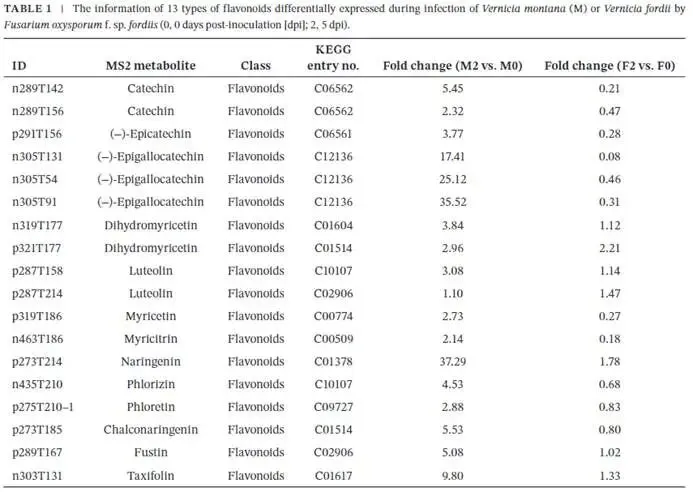
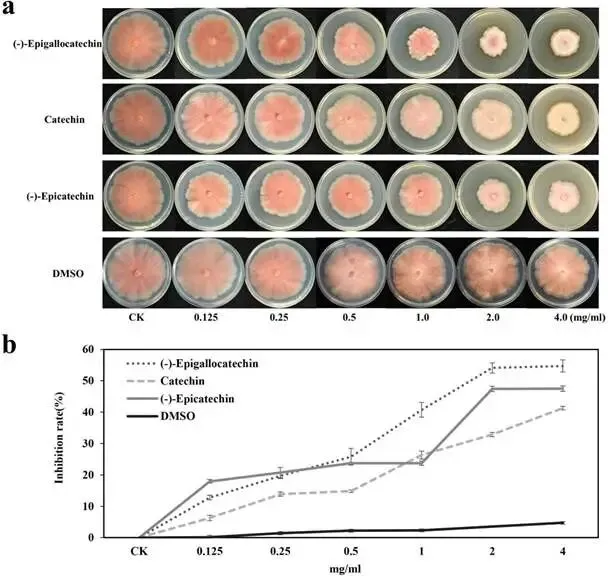
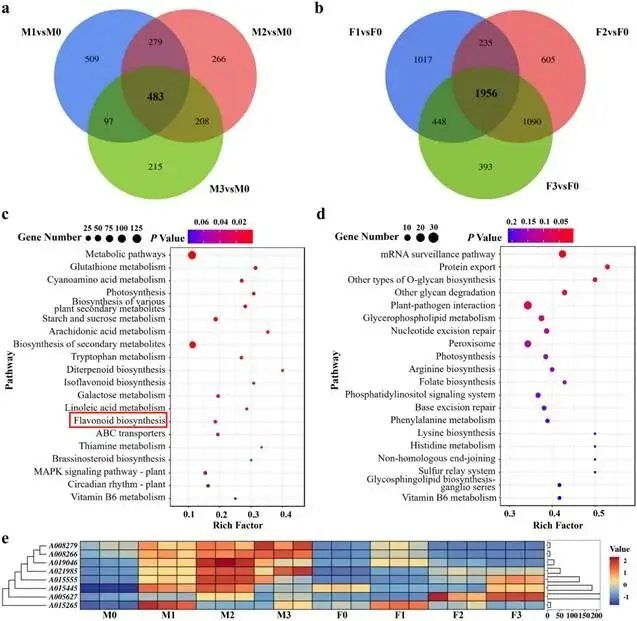
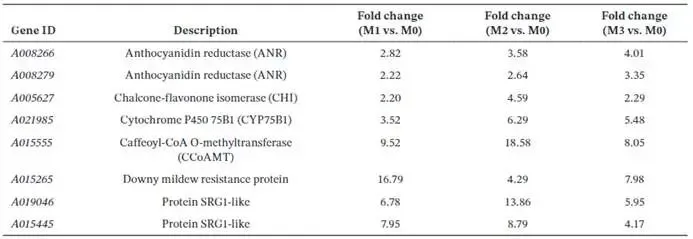
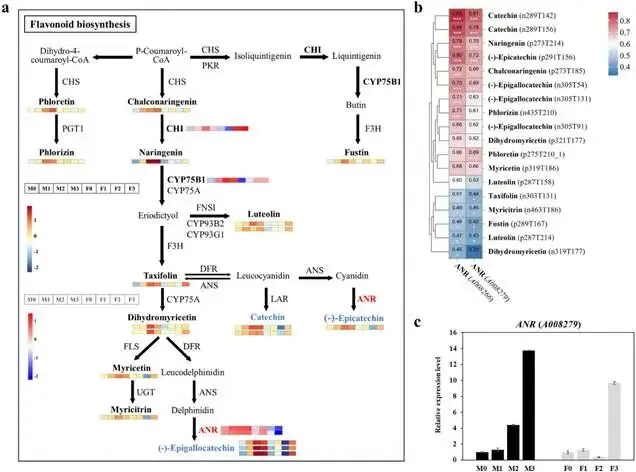
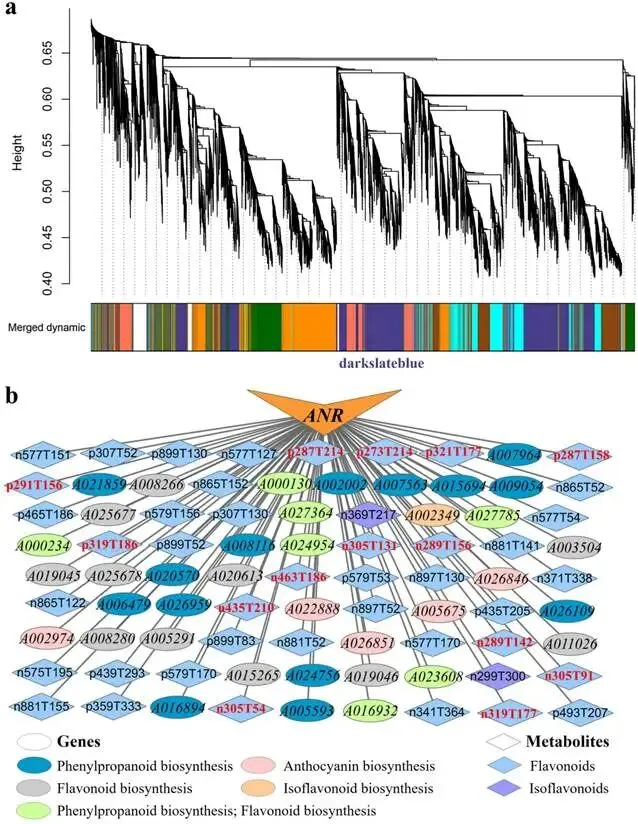
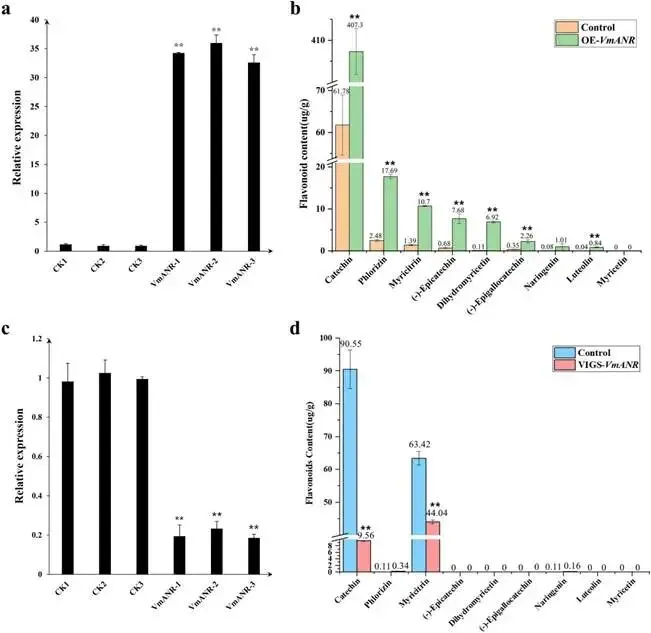
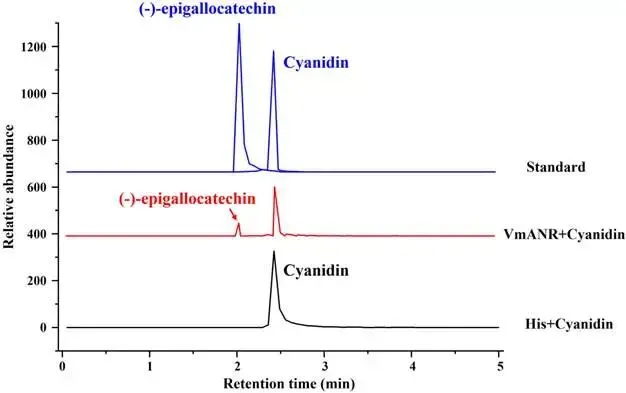
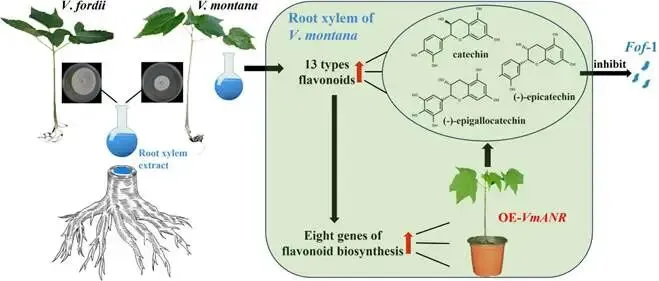
由尖孢镰刀菌油桐专化型1号生理小种(Fusarium oxysporum f. sp. fordiis, Fof-1)引起的油桐枯萎病,对油桐(Vernicia fordii)的生产构成了严重威胁。幸运的是,中国的另一种本土油桐属植物——千年桐(Vernicia montana,又称木油桐)表现出对该病原体的高度抗性。这种抗病与感病的油桐材料为研究抗枯萎病的机制提供了基础。与感病的油桐相比,抗病千年桐的根部木质部提取物能显著抑制Fof-1的生长。对千年桐根部木质部的代谢组学分析表明,在受到Fof-1侵染后,有13类黄酮类化合物的含量增加。对这13种黄酮类化合物的抗菌活性测定显示,儿茶素、(−)-表儿茶素和(−)-表没食子儿茶素对Fof-1的生长具有明显的抑制作用。转录组学分析进一步揭示,在千年桐受到Fof-1侵染后,多个上调表达基因也富集在黄酮类生物合成途径中。其中,花色素还原酶(ANR)基因直接参与具有抗菌活性的(−)-表儿茶素和(−)-表没食子儿茶素的生物合成。此外,过表达 VmANR 基因的转基因千年桐株系中,有8类黄酮类化合物的浓度升高;而沉默 VmANR 则导致儿茶素和杨梅苷的水平显著降低。体外酶活性测定进一步证实,VmANR能够催化底物矢车菊素生成(−)-表没食子儿茶素。本研究鉴定出 VmANR 是一个关键基因,它通过促进抗菌类黄酮的生物合成来塑造对Fof-1侵染的抗性,并为培育抗镰刀菌的油桐树种提供了有效的策略。篇名: Overexpression of Anthocyanidin Reductase Increases Flavonoids Content to Combat Fusarium Wilt in the Root Xylem of Vernicia montana期刊: Molecular Plant Pathology本研究以油桐(V. montana 与V. fordii)及病原菌 Fof-1 为材料。采用乙醇超声提取根系木质部代谢物,通过平板培养法测定其及13类黄酮化合物对病原菌的抑制活性。对接种病原菌不同时间点(0、2、5、9 dpi)的样本,进行转录组(Illumina Novaseq 6000测序)与代谢组(LC-MS)联合分析,并利用WGCNA构建基因-代谢物关联网络。通过实时荧光定量PCR(RT-qPCR)、毛根转化(V. montana 过表达与VIGS沉默 VmANR 基因)及原核表达纯化VmANR蛋白进行酶活验证,系统解析黄酮合成通路响应病原侵染的分子机制。(1) 千年桐根部木质部提取物对Fof-1生长表现出明显的抑制作用通过观察病原菌Fof-1在马铃薯葡萄糖琼脂(PDA)平板上的生长情况,测定了千年桐与油桐根部木质部提取物对Fof-1的抑制差异。研究发现,将千年桐的根部木质部提取物加入PDA后,能强烈抑制Fof-1的分生孢子萌发和菌丝生长(图1a)。相比之下,油桐的根部木质部提取物与提取溶剂对照(乙酸乙酯)及空白对照相比,对Fof-1的生长均无抑制作用(图1a)。图1. 千年桐和油桐根部木质部提取物对尖孢镰刀菌油桐专化型在马铃薯葡萄糖琼脂平板上生长的抑制情况 (a) 及抑制率 (b)。至第2天,千年桐提取物的抑制率达到最高值59.2%(图1b)。为比较两种提取物的抑制差异,我们计算了第2至8天根部木质部提取物的平均抑制率。千年桐和油桐提取物的平均抑制率分别为52.4%和21.0%。(2) Fof-1侵染后千年桐根部木质部中黄酮类化合物富集在千年桐和油桐的根部木质部中共检测到1921种次生代谢物。为观察这些代谢物在Fof-1侵染四个时期的含量变化,我们使用MeV4软件包以代谢物的归一化含量值绘制了热图(图S1)。结果显示,在千年桐的M2和M3阶段,有269种代谢物的含量增加。然而在油桐中,这269种代谢物表现出滞后性,仅在F3阶段含量才增加(图2a)。KEGG富集分析表明,这269种差异代谢物在以下通路中显著富集:黄酮类生物合成、淀粉和蔗糖代谢、半乳糖代谢、黄酮和黄酮醇生物合成以及维生素B6代谢(p < 0.05)(图2b,表S1)。值得注意的是,其中黄酮类生物合成通路的p值最为显著。共鉴定出该通路中的18种代谢物,并将其归类为13种黄酮类化合物:儿茶素、(−)-表儿茶素、(−)-表没食子儿茶素、二氢杨梅素、木犀草素、杨梅素、杨梅苷、根皮素、根皮苷、查耳酮柚皮素、柚皮素、紫杉叶素和花旗松素。图2c展示了这13种黄酮类化合物在千年桐和油桐四个侵染时期的含量倍数变化。我们观察到,千年桐中这13种黄酮类化合物的倍数变化均显著高于油桐。其中,柚皮素、(−)-表没食子儿茶素、花旗松素、查耳酮柚皮素和儿茶素的倍数变化超过了5(表1)。图2. 千年桐(M)与油桐(F)根部木质部的代谢组学分析。(a) 269种代谢物在尖孢镰刀菌油桐专化型侵染四个阶段(0, 1, 2, 3)的热图。(b) 269种差异表达代谢物前20条KEGG通路的散点图。黄酮类生物合成通路的p值最为显著,已用红色框标出。13种黄酮类化合物富集于黄酮类生物合成通路。(c) 黄酮类生物合成通路中13种黄酮类化合物含量的倍数变化。M:千年桐;F:油桐。柚皮素、(−)-表没食子儿茶素、花旗松素、查耳酮柚皮素和儿茶素的倍数变化 > 5。M0和F0:接种后0天;M1和F1:接种后2天;M2和F2:接种后5天;M3和F3:接种后9天。(3) 儿茶素、(−)-表儿茶素和(−)-表没食子儿茶素在体外对Fof-1表现出抑制作用针对上述代谢组学分析中鉴定出的13种黄酮类化合物,我们在体外评估了它们在不同浓度下对Fof-1的抗菌活性。与溶剂对照(二甲基亚砜,DMSO)和空白对照相比,仅有三种植物质((−)-表没食子儿茶素、儿茶素和(−)-表儿茶素)能抑制Fof-1的生长,其中(−)-表没食子儿茶素的抗菌活性最强(图3a)。我们进一步测试了不同浓度下的抑制效果。如图3b所示,在0.125和0.25 mg/mL浓度下,(−)-表儿茶素的抑制效果最为明显。此外,当浓度超过0.5 mg/mL时,(−)-表没食子儿茶素表现出最强的抑制效果。当浓度为4 mg/mL时,(−)-表没食子儿茶素的抑制率达到最大值54.7%。图3. (−)-表没食子儿茶素、儿茶素、(−)-表儿茶素及溶剂对照(二甲基亚砜,DMSO)在体外不同浓度下对尖孢镰刀菌油桐专化型的生长抑制情况 (a) 及抑制率 (%) (b)。(4) 千年桐中上调基因在黄酮类生物合成途径中显著富集基于11,837个转录本的FPKM值,分别对千年桐中M1、M2、M3与M0时期,以及油桐中F1、F2、F3与F0时期进行两两比较(设定倍数变化≥2且p值≤0.05)。在千年桐中鉴定出3523个差异表达基因(DEGs),在油桐中鉴定出7976个DEGs。通过绘制维恩图,分析了千年桐(M1 vs. M0, M2 vs. M0, M3 vs. M0)和油桐(F1 vs. F0, F2 vs. F0, F3 vs. F0)三组比较中上调DEGs的重叠与特异性。我们发现,千年桐有三组共有的上调DEGs为483个,而油桐共有1956个(图4a,b)。对于抗病的千年桐,KEGG富集分析显示,这483个上调DEGs显著富集于以下通路:代谢途径、谷胱甘肽代谢、氰基氨基酸代谢、光合作用、多种植物次生代谢产物的生物合成、淀粉和蔗糖代谢、花生四烯酸代谢、次生代谢产物生物合成、色氨酸代谢、二萜类生物合成、异黄酮生物合成、半乳糖代谢、亚油酸代谢、黄酮类生物合成、ABC转运蛋白、硫胺素代谢和油菜素甾醇生物合成(p < 0.05)(图4c,表S2)。在油桐中,1956个上调DEGs富集的KEGG通路则包括mRNA监视途径、蛋白质输出以及其他类型的O-聚糖生物合成等(p < 0.05)(图4d,表S3)。图4. 千年桐(M)与油桐(F)根部木质部的转录组学分析。(a) M1 vs. M0、M2 vs. M0和M3 vs. M0组中上调基因的维恩图。(b) F1 vs. F0、F2 vs. F0和F3 vs. F0组中上调基因的维恩图。(c) 千年桐中483个上调基因前20条KEGG通路的散点图。(d) 油桐中1956个上调基因前20条KEGG通路的散点图。(e) 尖孢镰刀菌油桐专化型侵染期间,黄酮类生物合成途径中八个上调基因的热图。右侧柱状图显示平均FPKM值。M0和F0:接种后0天;M1和F1:接种后2天;M2和F2:接种后5天;M3和F3:接种后9天。与代谢组学结果一致,上调的DEGs在千年桐中也富集于黄酮类生物合成途径(图4c)。然而,该通路在油桐中并未富集(图4d)。黄酮类生物合成途径中八个上调DEGs的信息列于表2,包括花色素还原酶(ANR)、查尔酮-黄烷酮异构酶(CHI)、咖啡酰辅酶A-O-甲基转移酶(CCoAMT)、细胞色素P450 75B1(CYP75B1)、类SRG1蛋白和霜霉病抗性蛋白。基于FPKM值绘制的这八个上调DEGs在千年桐和油桐中的热图如图4e所示。这些结果表明,在Fof-1侵染期间,千年桐黄酮类生物合成途径中的基因与代谢物之间存在关联。(5) 转录组与代谢组联合分析揭示黄酮类生物合成途径中基因与代谢物的协同表达我们的结果表明,Fof-1侵染触发了千年桐的直接防御反应,包括次生代谢产物的产生和黄酮类生物合成途径中基因表达的诱导。更具体地说,在千年桐中,包括(−)-表儿茶素、儿茶素、花旗松素、查耳酮柚皮素、二氢杨梅素、杨梅素和柚皮素在内的13种黄酮类物质的丰度在M2阶段急剧增加,并在M3阶段保持高水平。然而,这些黄酮类物质在油桐中仅在F3阶段才增加。与黄酮含量增加的趋势相似,在千年桐中,黄酮类生物合成途径中的八个基因在Fof-1侵染后表达也上调。其中,编码四个关键代谢酶的基因,即ANR(A008279, A008266)、CHI(A005627)和CYP75B1(A021985),参与了上述13种黄酮类物质的生物合成(图5a)。图5. 13种黄酮类化合物与黄酮类生物合成途径中四个基因的关联分析。(a) 黄酮类生物合成途径总结。热图显示了尖孢镰刀菌油桐专化型侵染期间13种黄酮类化合物和四个基因的丰度。表没食子儿茶素、儿茶素和(−)-表儿茶素用蓝色字体标出。两个ANR基因用红色字体标出。(b) 黄酮类生物合成途径中13种黄酮类化合物与两个ANR基因的相关性分析。*p < 0.05, **p < 0.01, ***p < 0.001。(c) ANR(A008279)基因在千年桐(M)和油桐(F)中响应侵染的逆转录定量PCR验证。M0和F0:接种后0天;M1和F1:接种后2天;M2和F2:接种后5天;M3和F3:接种后9天。基于对13种黄酮类化合物的抗菌实验,我们发现(−)-表没食子儿茶素、儿茶素和(−)-表儿茶素对Fof-1的生长有明显抑制作用(图3a)。如图5a所示,ANR(基因A008279和A008266)直接参与(−)-表儿茶素和(−)-表没食子儿茶素的生物合成。因此,选择A008279和A008266进行进一步分析和功能验证。相关性分析表明,A008279和A008266的表达与大多数黄酮类化合物(如儿茶素、柚皮素、(−)-表儿茶素、查耳酮柚皮素、(−)-表没食子儿茶素、根皮苷、二氢杨梅素、根皮素和杨梅素)的丰度呈正相关(图5b)。我们利用NCBI数据库对两个ANR基因的编码核苷酸序列进行了保守结构域搜索。结果显示,基因ANR A008266的保守结构域不完整,因此我们选择基因ANR A008279来验证其潜在的抗病功能。通过RT-qPCR分析检测了基因ANR(A008279)的表达。在Fof-1侵染的所有阶段,ANR(A008279)在千年桐中均上调(图5c),但在油桐中,该基因在F2阶段下调,在F3阶段才上调(图5c)。(6) ANR基因与黄酮类生物合成途径中其他差异表达基因及代谢物的基因网络根据千年桐和油桐中11,837个转录本的FPKM值以及1921种次生代谢物的含量值,我们使用R软件包的加权基因共表达网络分析(WGCNA)方法,构建了权重截断值 > 0.40的基因与代谢物调控网络。分析显示,ANR(A008279)在黄酮类生物合成、苯丙素生物合成、花青素生物合成和异黄酮生物合成等途径中,与另外39个边界基因和44个边界代谢物存在极高的连接性(图6a,深岩蓝色模块)。使用Cytoscape软件展示的共表达网络如图6b所示。值得注意的是,在黄酮类生物合成途径的13种化合物中,有9种被鉴定为边界代谢物,并在图6b中用红色字体标出。这九种黄酮类物质是:儿茶素(n289T142, n289T156)、(−)-表儿茶素(p291T156)、(−)-表没食子儿茶素(n305T131, n305T54, n305T91)、二氢杨梅素(n319T177, p321T177)、木犀草素(p287T158, p287T214)、杨梅素(p319T186)、杨梅苷(n463T186)、根皮苷(n435T210)和柚皮素(p273T214)。我们推测,ANR(A008279)基因可能与参与黄酮类生物合成的其他基因共表达,从而产生不同的黄酮类物质以抵御枯萎病。图6. ANR基因与黄酮类生物合成途径中其他差异表达基因(DEGs)和代谢物的加权基因共表达网络分析(WGCNA)。(a) 通过平均连锁层次聚类获得的基因树状图。深岩蓝色模块代表黄酮类生物合成途径中的DEGs和代谢物模块。(b) ANR(A008279)与39个边界基因和44个边界代谢物之间的基因网络。上述黄酮类生物合成途径中13种化合物里的9种以红色字体标出。(7) 过表达VmANR增加千年桐中黄酮类化合物的产生为探究千年桐中ANR基因(VmANR)表达水平的升高是否与黄酮含量相关,我们扩增了VmANR的开放阅读框(ORF)并将其插入过表达载体。由于千年桐是缺乏高效转化系统的木本植物,本研究采用发根农杆菌介导的毛状根诱导系统来检测VmANR在千年桐中的功能。以含有空载体的转基因毛状根作为对照,分别用对照载体或过表达(OE)载体VmANR(OE-VmANR)对千年桐根茎进行农杆菌转化。随后将幼苗在相同条件下培养40天,在对照组和OE-VmANR组中均诱导出了毛状根。我们通过RT-qPCR验证筛选出对照组和OE-VmANR组的转基因阳性毛状根。使用高效液相色谱-质谱(HPLC-MS)系统检测了WGCNA中提到的九种黄酮类物质,即儿茶素、(−)-表儿茶素、(−)-表没食子儿茶素、二氢杨梅素、木犀草素、杨梅素、杨梅苷、根皮苷和柚皮素的含量。培养40天后,OE-VmANR组中目的基因的表达量比对照组提高了约35倍(图7a)。同样地,HPLC-MS结果显示,与对照组相比,OE-VmANR转基因毛状根中六种黄酮类化合物(儿茶素、(−)-表儿茶素、(−)-表没食子儿茶素、杨梅苷、根皮苷和柚皮素)的含量增加了6.5至12.1倍。其中,二氢杨梅素和木犀草素的含量分别提高了约63.5倍和19.8倍(图7b,表S4)。值得注意的是,在OE-VmANR转基因毛状根和对照组中均未检测到杨梅素(图7b)。因此,在转基因毛状根中引入的OE-VmANR激活了黄酮类生物合成途径的代谢通量,从而增强了千年桐中黄酮类物质的生成。图7. 过表达OE-VmANR转基因毛状根中ANR基因表达的逆转录定量PCR(RT-qPCR)验证(a)及九种黄酮类化合物含量的检测(b)。病毒诱导基因沉默VIGS-VmANR叶片中ANR基因表达的RT-qPCR验证(c)及九种黄酮类化合物含量的检测(d)。显著差异,**p < 0.01。(8) 沉默VmANR降低千年桐中黄酮类化合物的产生为深入探究VmANR在九种黄酮类化合物生物合成中的作用,我们采用病毒诱导基因沉默(VIGS)方法,在千年桐中暂时性沉默其表达。将含有pTRV2-VmANR载体的根癌农杆菌导入千年桐叶片。18天后,检查千年桐的生长状态并进行各项分析。RT-qPCR分析证实,VIGS植株叶片中VmANR的表达被成功下调,而在对照植株中其表达保持不变(图7c)。此外,与对照植株相比,VIGS植株中儿茶素和杨梅苷的含量分别降低了9.5倍和1.5倍(图7d,表S5)。值得注意的是,在VIGS-VmANR植株叶片和对照植株叶片中,均未检测到五种黄酮类物质((−)-表儿茶素、二氢杨梅素、(−)-表没食子儿茶素、木犀草素、杨梅素)的含量(图7d)。这可能是因为千年桐的根组织含有更丰富种类和更高浓度的黄酮类物质。ANR是原花青素生物合成中的关键酶。原花青素的合成是黄酮类生物合成的一部分。在拟南芥中,已报道BANYULS具有花色素还原酶的功能,能在体外催化花色素转化为黄烷-3-醇。蒺藜苜蓿和拟南芥中ANR的功能已在体外进行研究。ANR能将矢车菊素主要转化为(−)-表儿茶素,副产物为儿茶素;并能将飞燕草素等量转化为(−)-没食子儿茶素和(−)-表没食子儿茶素。为进一步探索VmANR在调控黄酮类代谢物积累中的功能,我们扩增了VmANR的ORF,并将其克隆到表达载体pET-32a中,从而在VmANR的N端添加了一个6×His标签,并在大肠杆菌BL21中表达。我们纯化了His-VmANR融合蛋白,并在体外以商品化的矢车菊素和飞燕草素为底物测试了其活性。使用HPLC分析,我们检测到His-VmANR催化反应中的产物,包括(−)-表儿茶素、儿茶素、(−)-没食子儿茶素和(−)-表没食子儿茶素。HPLC数据显示,千年桐中存在一种新的ANR催化反应,即能够以矢车菊素为底物催化生成(−)-表没食子儿茶素(图8,表S6)。然而,当底物为飞燕草素时,未检测到上述四种产物中的任何一种。图8. VmANR在体外对矢车菊素的酶活性测定。His标签蛋白用作阴性对照。本研究发现,与感病的油桐相比,抗病的千年桐其根部木质部提取物对Fof-1的生长表现出强烈的抑制作用。通过转录组与代谢组的联合分析,我们筛选出在千年桐受Fof-1侵染后含量增加的13种黄酮类化合物,以及黄酮类生物合成途径中表达上调的八个基因。过表达VmANR基因提高了多种黄酮类化合物的含量,其中儿茶素、(−)-表儿茶素和(−)-表没食子儿茶素对Fof-1的生长具有明显的抑制作用。上述可能的作用机制总结于图9。本研究确定了VmANR是一个关键基因,它通过促进抗菌类黄酮的生物合成来塑造对Fof-1侵染的抗性,从而为培育抗镰刀菌的油桐树种提供了策略。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
