本研究使用的主要植物材料包括二穗短柄草(Brachypodium distachyon)野生型材料Bd21、四倍体小麦品种Kronos及其EMS突变体库、以及本氏烟草(N. benthamiana)。所有植株均在长日照(16小时光照/8小时黑暗)和精确温控(22℃/18℃)的生长箱中培养。为创制基因敲除突变体,通过Golden Gate法将CRISPR/Cas9导向RNA(gRNA)克隆至pYLCRISPR/Cas9PUbi载体。对于过表达株系,则将FKF1和SWN等基因的全长编码序列(CDS)克隆至pCUBI1390载体。所有二穗短柄草的遗传转化均通过农杆菌介导法完成。基因表达水平的检测采用实时荧光定量PCR(RT-qPCR)技术。首先使用Trizol试剂提取总RNA,经M-MLV逆转录酶合成cDNA后,在CFX96实时PCR系统上进行扩增。基因的相对表达量基于三个生物学重复,通过2-ΔΔCt法进行计算。所有实验数据以“均值±标准差”表示,并采用GraphPad Prism软件进行统计分析与作图。两组间的显著性差异通过学生氏t检验(Student's t-test)进行判断,多组间的比较则采用单因素方差分析(one-way ANOVA)及Tukey's多重比较检验。
2.2 蛋白质相互作用检测
为了鉴定与FKF1相互作用的蛋白,利用酵母双杂交(Y2H)系统,以FKF1为诱饵蛋白,筛选了二穗短柄草的cDNA文库。筛选出的候选互作关系及其他蛋白间的互作,通过靶向Y2H实验进行验证,即观察共转化酵母在四缺(-LTHA)筛选培养基上的生长状况。为在体外验证蛋白间的直接物理互作,进行了Pull-Down实验。将His标签和MBP标签的重组蛋白(如His-ELF3和MBP-FKF1)在大肠杆菌中表达纯化后,通过亲和树脂共孵育,并用免疫印迹(Immunoblot)检测被拉下的蛋白。为在植物活细胞中验证互作,采用了荧光素酶互补(LCI)和双分子荧光互补(BiFC)实验。将目标蛋白分别与nLUC/cLUC或nYFP/cYFP融合表达,通过检测荧光素酶活性或YFP荧光信号来判断蛋白互作。此外,通过免疫共沉淀(Co-IP)实验在二穗短柄草原生质体或本氏烟草叶片中对体内互作进行最终确认。即在共表达不同标签(如Flag和HA)的蛋白后,使用相应抗体进行免疫沉淀,再通过免疫印迹检测是否存在共沉淀的蛋白。
2.3 蛋白质稳定性与染色质修饰分析
为研究FKF1对ELF3蛋白稳定性的调控,设计了多种蛋白降解实验。在无细胞降解(Cell-free Degradation)体系中,将纯化的His-ELF3蛋白与野生型或突变体植株的总蛋白提取物共孵育,通过免疫印迹检测ELF3的降解情况,并通过添加蛋白酶体抑制剂MG132来验证其是否依赖于26S蛋白酶体途径。为进一步确证FKF1的E3连接酶功能,进行了体外泛素化实验,检测FKF1是否能够催化ELF3的多聚泛素化修饰。最终,为探究ELF3调控下游基因转录的表观遗传机制,开展了染色质免疫沉淀(ChIP)实验。将交联后的植物组织进行染色质提取与超声破碎,随后使用特异性抗体(如抗H3K27me3抗体或标签蛋白抗体)富集与目标蛋白结合的染色质片段。通过对富集的DNA进行定量PCR(ChIP-qPCR),分析目标蛋白或组蛋白修饰在PPD1等基因启动子区域的富集水平,从而揭示其对基因表达的直接调控作用。
3.1 FKF1在二穗短柄草中促进开花
功能保守预测:基于与拟南芥同源蛋白的功能相似性,推测二穗短柄草中的FKF1同样调控开花。
过表达促开花:构建FKF1过表达株系,其在长日照条件下表现出显著的早花表型。
下游基因上调:在FKF1过表达株系中,关键开花基因PPD1和FT1的转录水平显著升高。
功能缺失晚花:利用CRISPR/Cas9技术创制fkf1功能缺失突变体,该突变体表现出明显的晚花表型。
下游基因下调:在fkf1突变体中,PPD1和FT1的表达受到显著抑制。
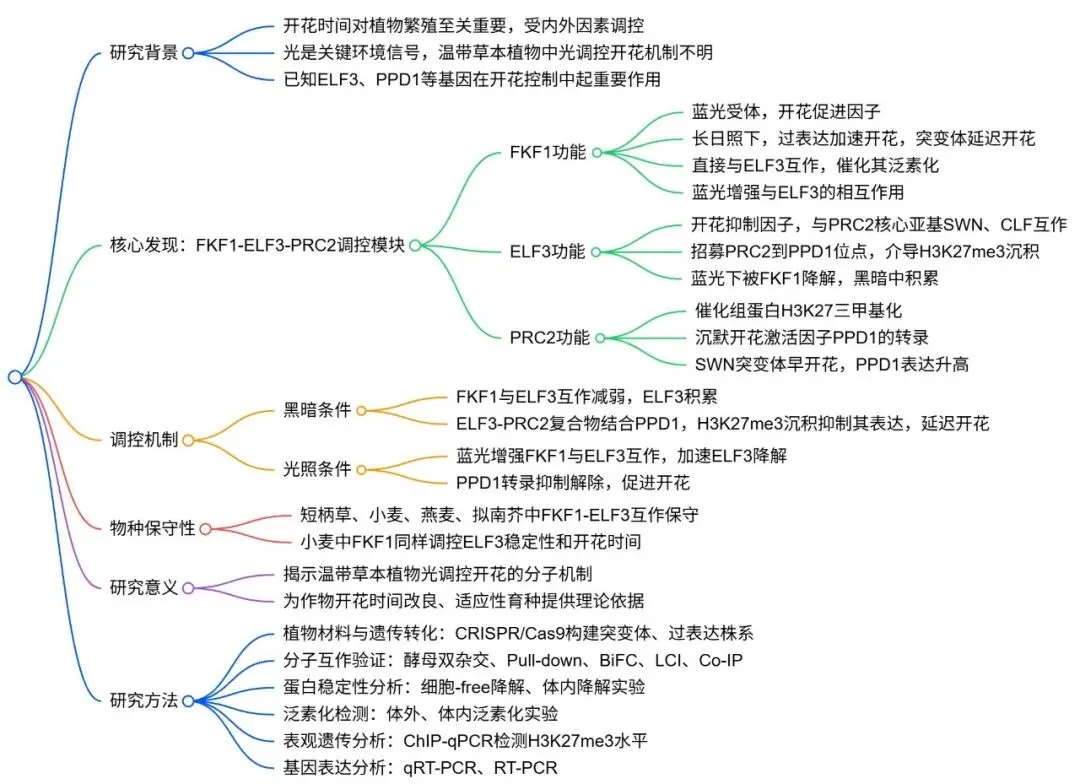
为了探究FKF1在二穗短柄草中的生物学功能,首先构建了FKF1的过表达转基因株系。在长日照条件下,这些过表达株系的开花时间显著早于野生型对照。对开花调控网络中的关键基因进行检测后发现,光周期途径基因PPD1及其下游的成花素基因FT1在FKF1过表达株系中均表现出转录水平的显著上调。与之相反,通过CRISPR/Cas9技术获得的fkf1功能缺失突变体则表现出显著的晚花表型,且其体内的PPD1和FT1基因转录水平受到明显抑制。这些结果共同表明,FKF1作为开花的正向调控因子,通过激活PPD1和FT1的表达来促进二穗短柄草的开花进程 (Fig. 1)。
图1. FKF1在二穗短柄草中作为开花激活因子。 (A) 野生型(WT)和两个独立的FKF1过表达(OE-1和OE-2)株系在长日照条件下的开花表型。(B) WT、OE-1和OE-2株系的抽穗时间。数据为均值±标准差(n=10)。(C) 在长日照条件下,于不同时间点采集的WT、OE-1和OE-2叶片中PPD1的相对表达水平。数据为均值±标准差(n=3)。(D) 在长日照条件下,于不同时间点采集的WT、OE-1和OE-2叶片中FT1的相对表达水平。数据为均值±标准差(n=3)。(E) WT和两个独立的fkf1突变体(fkf1-1和fkf1-2)在长日照条件下的开花表型。(F) WT、fkf1-1和fkf1-2株系的抽穗时间。数据为均值±标准差(n=10)。(G) 在长日照条件下,于不同时间点采集的WT、fkf1-1和fkf1-2叶片中PPD1的相对表达水平。数据为均值±标准差(n=3)。(H) 在长日照条件下,于不同时间点采集的WT、fkf1-1和fkf1-2叶片中FT1的相对表达水平。数据为均值±标准差(n=3)。图中星号表示根据双尾t检验,转基因株系与WT之间存在统计学上的显著差异(*p < 0.01)。
3.2 FKF1与ELF3发生物理相互作用
筛选互作靶标:通过酵母双杂交(Y2H)筛选,鉴定出ELF3是FKF1的潜在互作蛋白。
酵母体系验证:在Y2H系统中,确认了FKF1与ELF3之间存在直接的物理相互作用。
植物细胞验证:通过烟草叶片中的分裂荧光素酶(Split-LUC)互补实验,证实了二者在植物细胞内的相互作用。
原生质体验证:在二穗短柄草原生质体中进行的免疫共沉淀(Co-IP)实验,进一步确认了FKF1和ELF3蛋白在体内形成复合体。
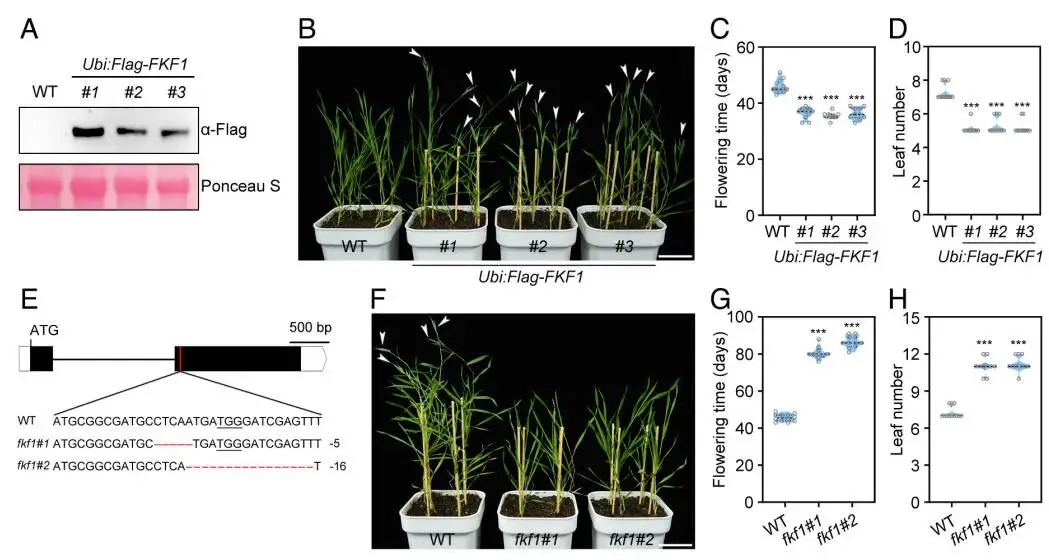
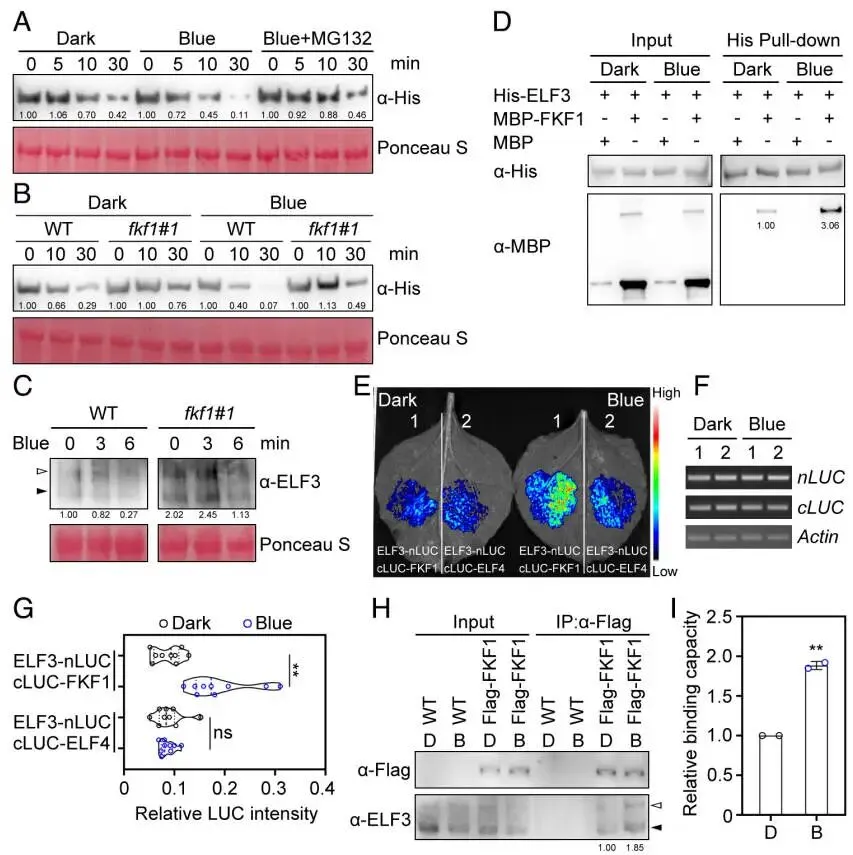
为了揭示FKF1调控开花的分子机制,采用酵母双杂交(Y2H)文库筛选的方法来鉴定其互作蛋白。结果表明,FKF1能够与开花抑制因子ELF3发生相互作用。这一互作关系通过靶向Y2H实验得到了进一步确认。随后,通过在烟草叶片中进行的分裂荧光素酶(Split-LUC)互补实验,证明了FKF1与ELF3在植物活细胞内确实存在物理互作。为了在二穗短柄草自身系统中验证此结论,在二穗短柄草原生质体中进行了免疫共沉淀(Co-IP)实验。结果显示,当FKF1-FLAG与ELF3-HA共表达时,FKF1-FLAG能够被ELF3-HA特异性地沉淀下来,反之亦然。综合这些来自不同实验体系的证据,可以确定FKF1与ELF3在体内外均能发生直接的物理相互作用 (Fig. 2)。
图2. FKF1与ELF3发生物理相互作用。 (A) 通过酵母双杂交文库筛选鉴定出ELF3是FKF1的互作蛋白。将含有pGBKT7-FKF1的酵母细胞与含有pGADT7-cDNA文库的酵母细胞进行交配,并在筛选培养基上进行筛选。(B) 在SD/-Leu/-Trp/-His/-Ade培养基上进行的靶向酵母双杂交实验,验证了FKF1与ELF3的相互作用。pGBKT7-53与pGADT7-T的互作用作阳性对照。(C) 在本氏烟草叶片中进行的分裂荧光素酶(Split-LUC)互补实验,显示了FKF1与ELF3的相互作用。将所示组合的质粒共转化至烟草叶片中。热图代表荧光信号强度。(D) 在二穗短柄草原生质体中进行的免疫共沉淀(Co-IP)实验,验证了FKF1与ELF3的相互作用。将35S:FKF1-FLAG与35S:ELF3-HA共转化至原生质体中,使用抗FLAG或抗HA琼脂糖凝胶珠进行免疫沉淀。使用抗FLAG和抗HA抗体进行蛋白质印迹分析。
3.3 FKF1介导ELF3的光依赖性降解
调控蛋白丰度:ELF3蛋白在FKF1过表达株系中积累减少,在fkf1突变体中积累增多。
介导体外降解:在无细胞降解实验中,FKF1能够直接导致ELF3蛋白的降解。
发挥泛素化功能:体外泛素化实验证明,FKF1可作为E3连接酶催化ELF3的泛素化修饰。
蓝光增强互作:Y2H和Split-LUC实验均表明,蓝光能够显著增强FKF1与ELF3的相互作用。
蓝光加速降解:无细胞降解实验显示,蓝光处理能显著加速FKF1对ELF3的降解过程。
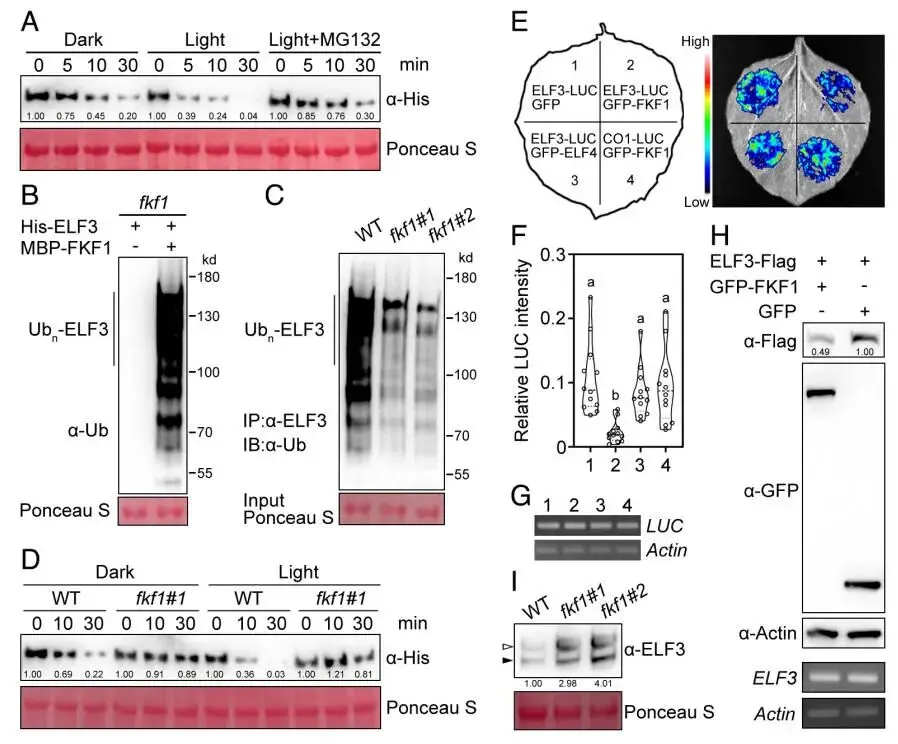
鉴于FKF1是一个E3泛素连接酶,且与ELF3互作,推测FKF1可能通过蛋白酶体途径调控ELF3的稳定性。蛋白质印迹分析显示,与野生型相比,ELF3蛋白在FKF1过表达株系中的丰度显著降低,而在fkf1突变体中则显著升高。体外无细胞降解实验进一步证实,FKF1的存在能够直接促进ELF3的降解,且这种降解可被蛋白酶体抑制剂MG132所阻断。体外泛素化实验的结果清晰地表明,FKF1能够对ELF3进行泛素化修饰。更有趣的是,光照条件对这一过程有显著影响。酵母双杂交和分裂荧光素酶实验均显示,蓝光能够特异性地增强FKF1与ELF3的相互作用,而红光或黑暗则无此效应。与此一致,在无细胞降解体系中,蓝光处理也显著加速了FKF1介导的ELF3蛋白降解。这些结果共同揭示了FKF1通过介导ELF3的泛素化和降解来调控其蛋白稳定性,且这一过程受到蓝光的正向调控 (Fig. 3)。
图3. FKF1介导ELF3的光依赖性降解。 (A) 在长日照条件下,于傍晚时间点(ZT15)采集的WT、OE-1和fkf1-1叶片中ELF3蛋白的丰度。使用抗ELF3抗体进行蛋白质印迹分析。肌动蛋白(Actin)作为上样对照。(B) FKF1介导的ELF3体外无细胞降解。将纯化的MBP-ELF3蛋白与含有GST或GST-FKF1的总蛋白提取物在有或无蛋白酶体抑制剂MG132的条件下共孵育。使用抗MBP抗体检测MBP-ELF3的蛋白水平。CBB(考马斯亮蓝)染色显示上样量均等。(C) FKF1在体外对ELF3进行泛素化。将纯化的MBP-ELF3和GST-FKF1蛋白与E1、E2和泛素(Ub)在反应缓冲液中孵育。使用抗MBP抗体检测泛素化的MBP-ELF3。(D) 蓝光增强FKF1与ELF3的相互作用。将共同转化了pGBKT7-FKF1和pGADT7-ELF3的酵母细胞在黑暗、白光、蓝光或红光下孵育3天。(E) Split-LUC实验显示蓝光增强FKF1与ELF3的相互作用。将所示质粒组合共转化至烟草叶片,并在不同光照条件下处理24小时。(F) 蓝光加速FKF1介导的ELF3降解。将纯化的MBP-ELF3蛋白与含有GST-FKF1的总蛋白提取物在黑暗或蓝光下共孵育。
3.4 ELF3与PRC2核心亚基相互作用
探究下游机制:为阐明ELF3抑制PPD1转录的机制,推测其可能与表观遗传调控复合体PRC2相关。
筛选PRC2互作:通过Y2H实验,系统性地检测了ELF3与PRC2复合体各核心亚基的相互作用。
鉴定互作亚基:Y2H结果显示,ELF3特异性地与PRC2的两个催化亚基SWN和CLF发生相互作用。
体内互作验证:在二穗短柄草原生质体中进行的Co-IP实验,证实了ELF3在体内确实能够与SWN和CLF形成蛋白质复合体。
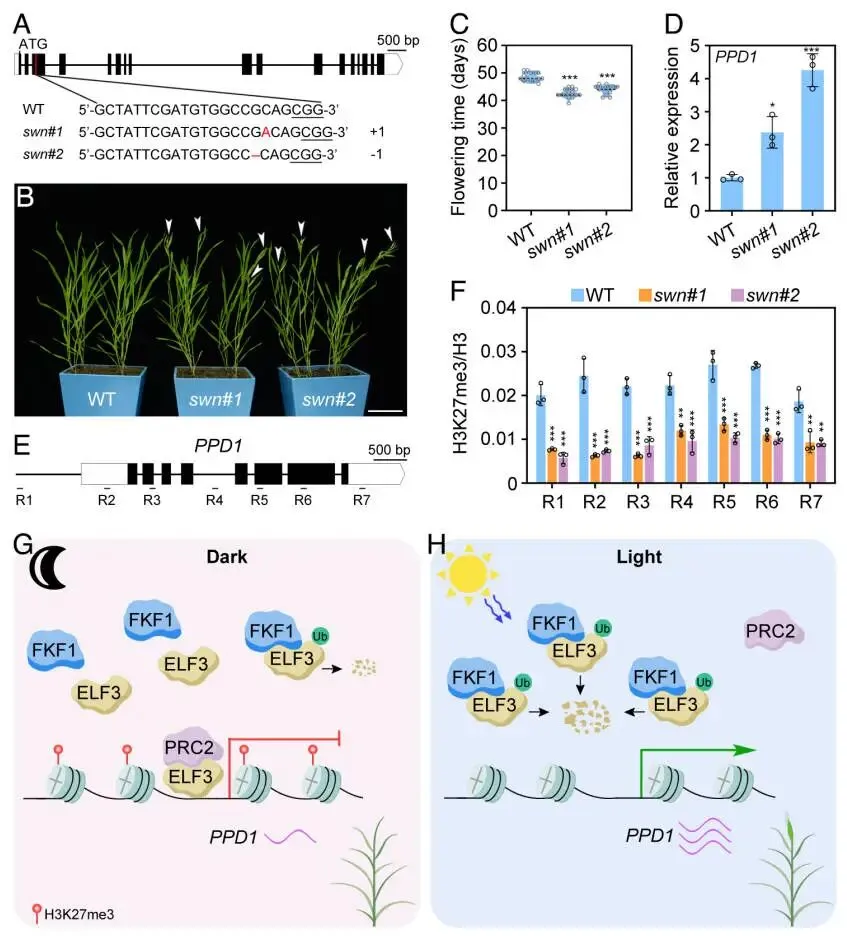
为了深入探究ELF3如何抑制PPD1的转录,提出了一个假设,即ELF3可能通过招募多梳抑制复合体2(PRC2)来实现这一功能。为了验证该假设,首先通过酵母双杂交(Y2H)系统地测试了ELF3与二穗短柄草中PRC2复合体所有核心亚基的相互作用可能性。结果显示,ELF3能够与PRC2的两个E(z)同源催化亚基SWN和CLF发生特异性相互作用,但与其他亚基(如FIE、MSI1、EMF2a和EMF2b)则没有互作。随后,在二穗短柄草原生质体中进行的免疫共沉淀(Co-IP)实验进一步验证了这些发现。结果表明,ELF3-HA能够被SWN-FLAG或CLF-FLAG特异性地沉淀下来。这些结果共同证明,ELF3在体内能够与PRC2的核心催化亚基SWN和CLF发生物理相互作用 (Fig. 4)。
图4. ELF3与PRC2的核心亚基相互作用。 (A) 酵母双杂交实验显示ELF3与PRC2的两个核心亚基SWN和CLF相互作用。将所示质粒组合共转化至酵母细胞中,并在SD/-Leu/-Trp/-His/-Ade培养基上进行筛选。(B) 在二穗短柄草原生质体中进行的Co-IP实验,验证了ELF3与SWN的相互作用。(C) 在二穗短柄草原生质体中进行的Co-IP实验,验证了ELF3与CLF的相互作用。将所示质粒组合共转化至原生质体中,使用抗FLAG琼脂糖凝胶珠进行免疫沉淀。使用抗FLAG和抗HA抗体进行蛋白质印迹分析。
3.5 ELF3将PRC2招募至PPD1基因座
分析染色质结合:通过染色质免疫沉淀(ChIP-qPCR)技术,检测ELF3是否直接结合PPD1的启动子区域。
确认ELF3结合:ChIP-qPCR结果显示,ELF3蛋白在PPD1基因的启动子区域有显著富集。
检测PRC2结合:进一步检测PRC2核心亚基SWN和CLF在PPD1启动子区域的富集情况。
确认ELF3依赖性:在elf3突变体中,SWN和CLF在PPD1启动子上的富集消失,证明其结合依赖于ELF3的存在。
分析组蛋白修饰:检测PPD1基因座上抑制性组蛋白标记H3K27me3的水平。
关联基因表达:H3K27me3的水平在野生型中较高,而在elf3和fkf1突变体中显著降低,与PPD1的转录激活状态呈负相关。
基于ELF3与PRC2亚基的互作,推测ELF3可能通过将PRC2招募至PPD1基因座来抑制其表达。为了验证这一点,进行了染色质免疫沉淀后定量PCR(ChIP-qPCR)实验。结果显示,在野生型植物中,ELF3蛋白在PPD1的启动子区域表现出显著富集。同样,PRC2的核心亚基SWN和CLF也在该区域富集。然而,在elf3功能缺失突变体中,SWN和CLF在PPD1启动子上的富集现象完全消失,这有力地证明了PRC2是被ELF3招募至PPD1基因座的。与此一致,对组蛋白修饰的检测发现,在野生型植物中,PPD1基因座上存在高水平的抑制性标记H3K27me3。而在elf3突变体中,该标记的水平显著下降,导致PPD1表达上调。值得注意的是,在fkf1突变体中,由于ELF3蛋白积累增加,其在PPD1启动子上的富集也相应增加,H3K27me3水平也随之升高,从而抑制了PPD1的表达。这些结果共同揭示了一个清晰的调控链条:ELF3作为引导因子,将PRC2复合体招募至PPD1基因座,通过催化H3K27me3修饰来抑制其转录 (Fig. 5)。
图5. ELF3将PRC2招募至PPD1基因座。 (A) PPD1基因的示意图。黑色框代表外显子,灰色框代表内含子,白色框代表非翻译区(UTR)。P1、P2和P3指示了用于ChIP-qPCR分析的区域。(B) 在WT和35S:ELF3-HA转基因株系中进行的ChIP-qPCR分析,显示了ELF3在PPD1启动子上的富集情况。(C) 在WT和elf3-1突变体中,使用抗SWN抗体进行的ChIP-qPCR分析,显示了SWN在PPD1启动子上的富集情况。(D) 在WT和elf3-1突变体中,使用抗CLF抗体进行的ChIP-qPCR分析,显示了CLF在PPD1启动子上的富集情况。(E) 在WT、elf3-1和fkf1-1中,使用抗H3K27me3抗体进行的ChIP-qPCR分析,显示了H3K27me3在PPD1启动子上的富集情况。(F) 在WT、elf3-1和fkf1-1中PPD1的相对表达水平。数据为均值±标准差(n=3)。星号表示根据双尾t检验,与WT相比存在统计学上的显著差异(*p < 0.01)。
揭示了完整的信号调控链条: 本研究不仅发现了单个的互作关系,而是完整地描绘了一条从环境信号感知到核心开花基因表达调控的完整分子路径。具体而言,该路径为:蓝光激活的FKF1蛋白通过泛素化途径降解开花抑制因子ELF3,解除ELF3对下游基因的抑制,最终激活PPD1和FT1的表达,从而促进开花。这一环环相扣的证据链,系统性地揭示了FKF1-ELF3-PPD1/FT1模块在二穗短柄草开花调控中的核心作用。
阐明了光依赖的核心分子机制: 研究的突出创新点在于明确了“光”这一环境信号如何被整合进蛋白调控网络。研究证实,蓝光能够显著增强FKF1与ELF3之间的物理相互作用,并加速由FKF1介导的ELF3蛋白降解过程。这为理解植物如何精确感知光周期变化,并将其转化为蛋白稳定性的改变,最终控制开花时间提供了直接的分子机制证据。
建立了开花调控与表观遗传的直接联系: 本研究创新性地将光周期调控途径与表观遗传抑制机制直接联系起来。研究发现,开花抑制因子ELF3扮演了“桥梁”的角色,它能够直接与表观遗传抑制复合物PRC2的核心催化亚基(SWN和CLF)相互作用,并将其特异性地招募至关键开花基因PPD1的启动子区域。
验证了招募介导的基因沉默模型: 基于上述发现,研究通过ChIP-qPCR等实验进一步证实,ELF3对PRC2的招募是导致PPD1基因座上产生抑制性组蛋白修饰(H3K27me3)并最终抑制其转录的直接原因。这一“招募-修饰-沉默”模型的建立,深刻揭示了ELF3作为转录抑制因子的具体工作方式,为理解基因表达的表观遗传调控提供了新的范例。
审核|林蘖亙
阅读科研文献,荟萃前沿进展
欢迎关注、转发、投稿、点赞





 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
