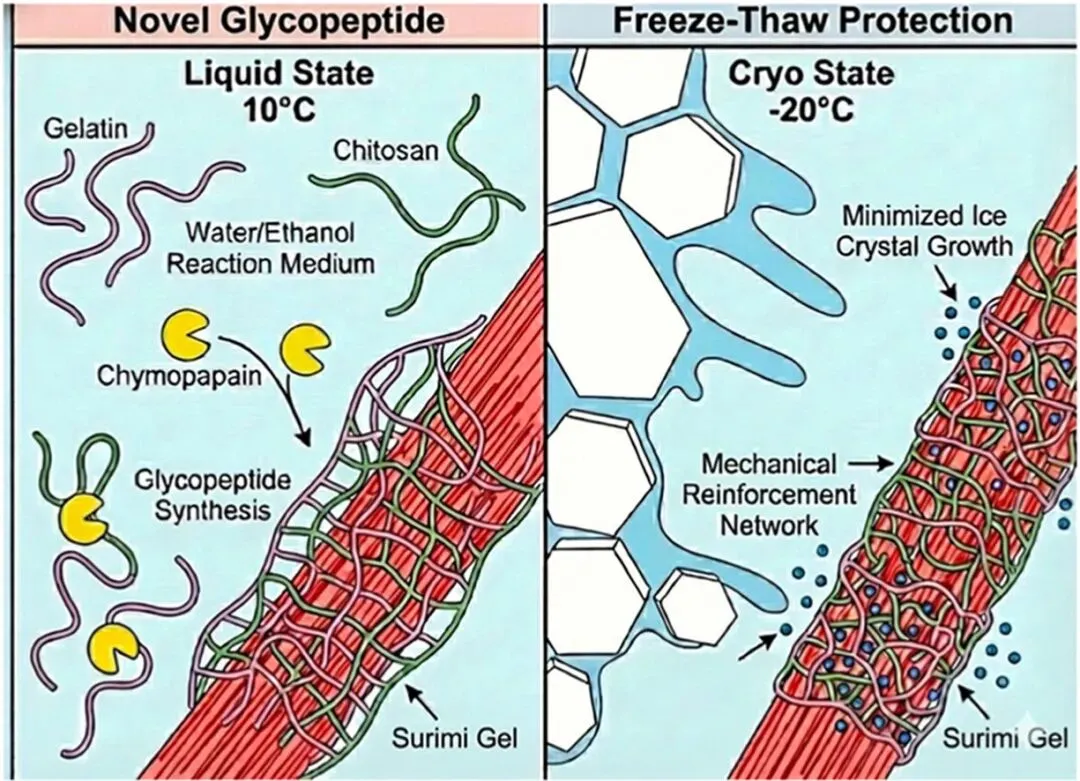
Food Hydrocolloids| 过冷态与液态下木瓜凝乳蛋白酶介导的鱼明胶糖基化修饰:结构编辑机制及在鱼糜凝胶中的相态依赖性抗冻作用
近日,安徽农业大学团队在《Food Hydrocolloids》期刊上发表了题为《Chymopapain-mediated glycosylation of fish gelatin in supercooled versus liquid states: Structural editing mechanisms and phase-dependent cryoprotection in surimi gels》的研究性论文(一区,IF:13.8)。该研究以木瓜凝乳蛋白酶催化鱼明胶与壳聚糖的糖基化接枝反应,系统对比了液态(10℃)、部分冻结(-5℃)与深冻(-20℃)三种热力学相态下的反应行为,结合分子对接与结构解析揭示了相态诱导的空间可及性调控机制,阐明了不同相态下酶对氨基酸残基的靶向选择性差异及产物结构重塑规律。研究将制备的共轭物以 1% 添加量应用于鱼糜凝胶,经 12 次冻融循环验证,不同相态合成的产物分别通过维持网络刚性、保护巯基基团或调控水分分布的差异化路径发挥抗冻保护作用,可将凝胶析水率降至约 12.5%,为冷冻食品高性能抗冻配料的精准创制提供了理论依据。
鱼明胶可规避疯牛病风险与宗教饮食限制,是哺乳动物水胶体的清洁标签替代原料,但冷水鱼源明胶天然存在脯氨酸、羟脯氨酸等亚氨基酸含量不足的缺陷,导致其热稳定性差、流变性能弱,且两亲性不足,难以稳定冷冻鱼糜等复杂多相体系以抵御界面失稳与冰晶重结晶。糖基化接枝是改善蛋白功能的有效策略,然而传统非酶美拉德反应条件严苛,易造成热敏性鱼胶原降解并生成有害的晚期糖基化终末产物;工业常用的微生物转谷氨酰胺酶更倾向催化蛋白同源自交联,且多糖链空间位阻大,难以实现高效异质接枝,易生成不溶性聚集体。低温酶法可在温和条件下实现接枝改性,但现有研究仅验证了单一低温条件的可行性,未厘清温度与溶剂物理相态的独立作用,不同冻结程度对酶的氨基酸位点选择性、产物分子结构及抗冻功能的调控机制尚未阐明,因此该研究系统探究三种热力学相态下木瓜凝乳蛋白酶介导的糖基化反应规律,旨在明确相态调控的结构编辑机制与抗冻构效关系。
相态通过空间可及性调控酶的位点选择性与产物结构
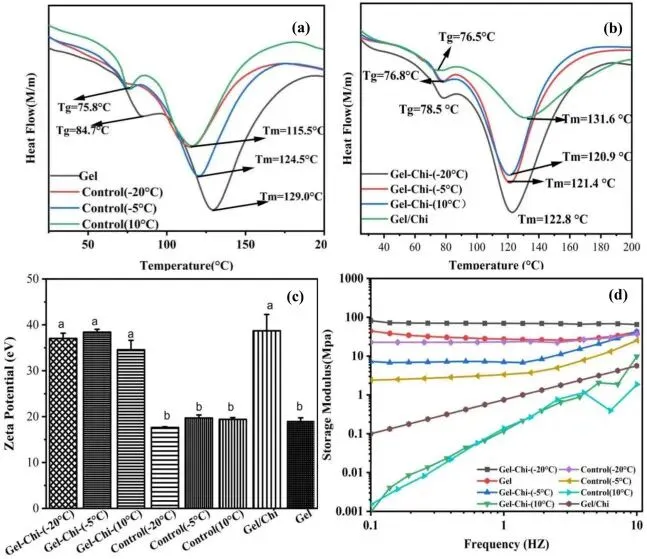
溶剂的物理相态是决定反应位点与产物架构的核心因素:-20℃深冻态基质刚性强、空间限制显著,酶被空间位阻 “过滤”,优先作用于无侧链的甘氨酸残基,产物三螺旋网络得到有效恢复,变性焓达峰值 160.9 J/g;-5℃部分冻结态兼具一定空间柔性,可实现对脯氨酸残基的靶向接枝;10℃液态下分子运动自由度最高,产物肽链伸展最充分,表面疏水性达约 240,接枝程度最高。
低温合成共轭物具备本征抗冻性与冻融结构稳定性
亚零下温度合成的共轭物因冷冻诱导的网络致密化而具备固有抗冻特性,可将自由水横向弛豫时间 T₂₃限制在约 400 ms;经反复冻融循环后,低温产物呈现独特的 “自适应收紧” 效应,水分向更紧密的毛细域迁移,网络未发生不可逆破损。相比之下,液态产物虽成胶能力较弱,但构建了以共价键为主、疏水作用为辅的稳定网络,氢键结合能更高。
鱼糜体系中呈现差异化的相态依赖性抗冻路径
以 1% 添加量应用于鱼糜凝胶并经 12 次冻融循环后,三类共轭物均可将析水率从 22.5% 降至约 12.5%,但作用机制存在显著差异:10℃液态产物作为力学增强剂,可嵌入肌原纤维网络维持刚性,同时通过空间屏蔽与化学抗氧化双重作用保护总巯基,抑制二硫键介导的肌动球蛋白聚集;-5℃产物作为水合动力学调节剂,可完全消除自由水组分,将固定化水弛豫时间 T₂₂缩短至 20 ms,从动力学上抑制冰晶生长。
反应遵循相态过滤驱动的转糖基化催化机制
分子对接结果证实,相态通过空间位阻实现底物的 “选择性过滤”:甘氨酸富集序列可深入酶活性中心结合,脯氨酸富集序列仅能在酶表面锚定。反应本质为木瓜凝乳蛋白酶催化的逆水解转糖基化反应,由乙醇 - 水共溶剂体系降低水活度驱动;低温条件下,冰结晶引发的 “冷冻浓缩效应” 可大幅提升未冻液相中底物与酶的局部浓度,补偿低温带来的动力学衰减,最终实现位点可控的分子结构编辑。
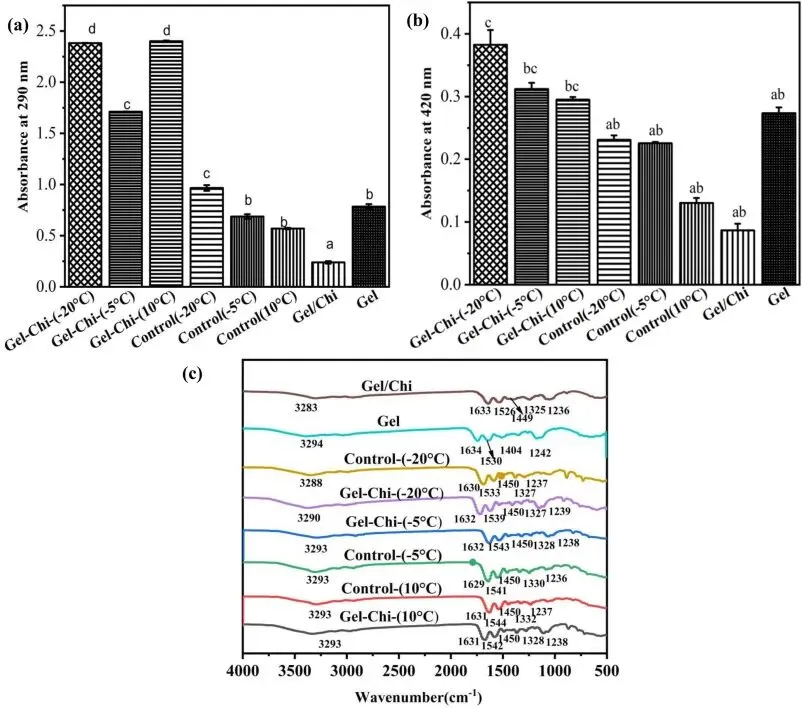
Fig. 1. UV-Vis and FTIR spectra of glycosylated gelatin, control group, gelatin, and gelatin/chitosan mixture.
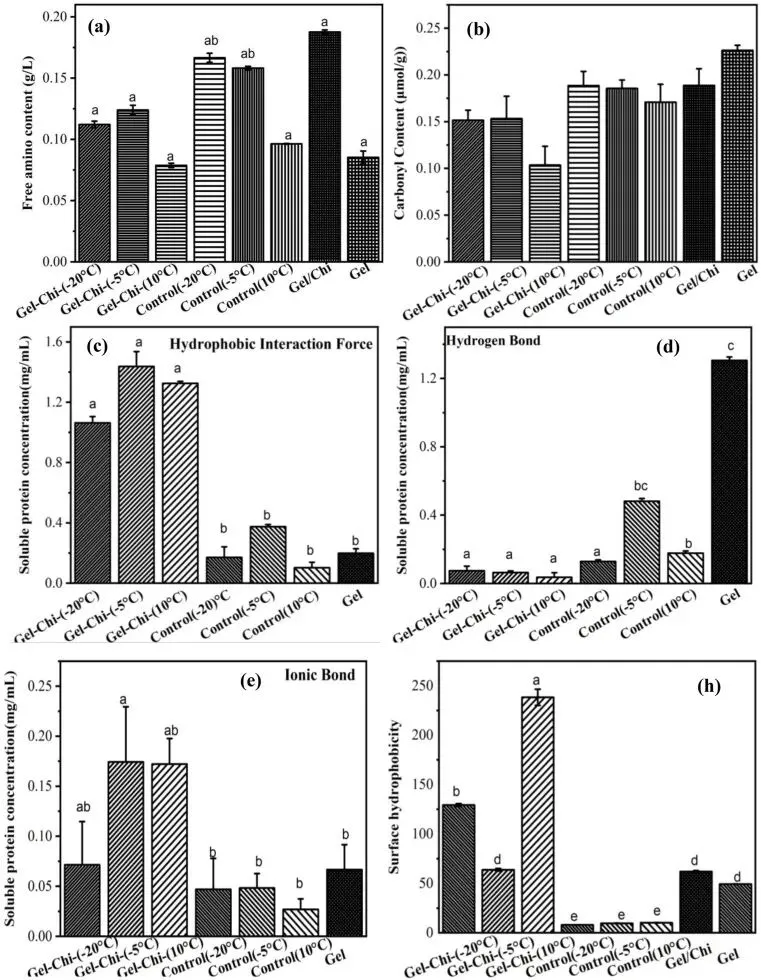
Fig. 2. Free amino group content (a), carbonyl group content (b), intermolecular forces (c–e), and surface hydrophobicity (f) of glycosylated gelatin, control group, gelatin, and gelatin/chitosan mixture.
Fig. 3. DSC thermograms (a–b), zeta potential (c), and storage modulus (d) of glycosylated gelatin, control group, gelatin, and gelatin/chitosan mixture.
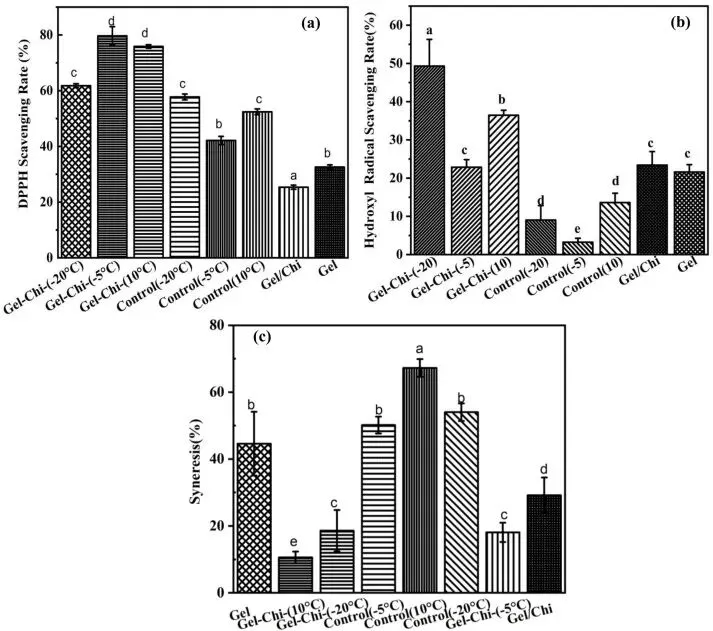
Fig. 4. DPPH radical scavenging activity (a), hydroxyl radical scavenging activity (b), and syneresis (c) of glycosylated gelatin, control group, gelatin, and gelatin/chitosan mixture.
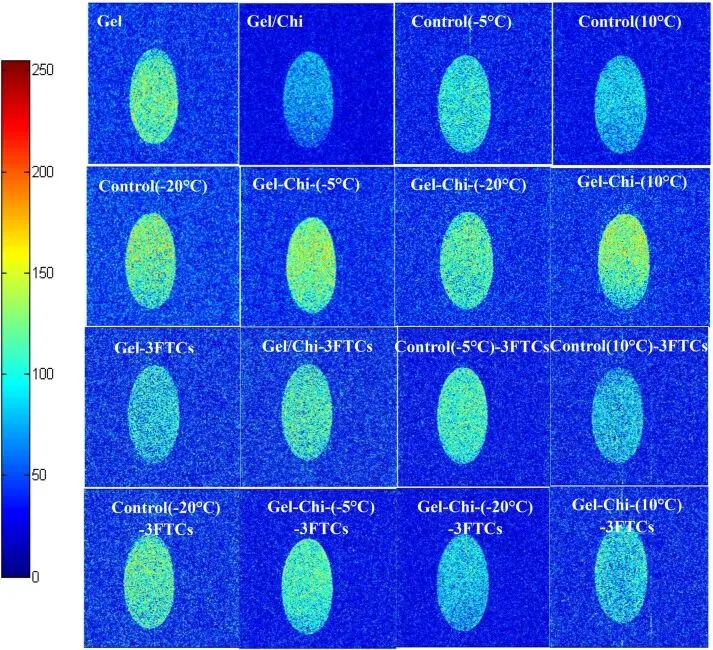
Fig. 5. Water distribution of glycosylated gelatin, control group, gelatin, and gelatin/chitosan mixture before and after freeze-thaw cycles.
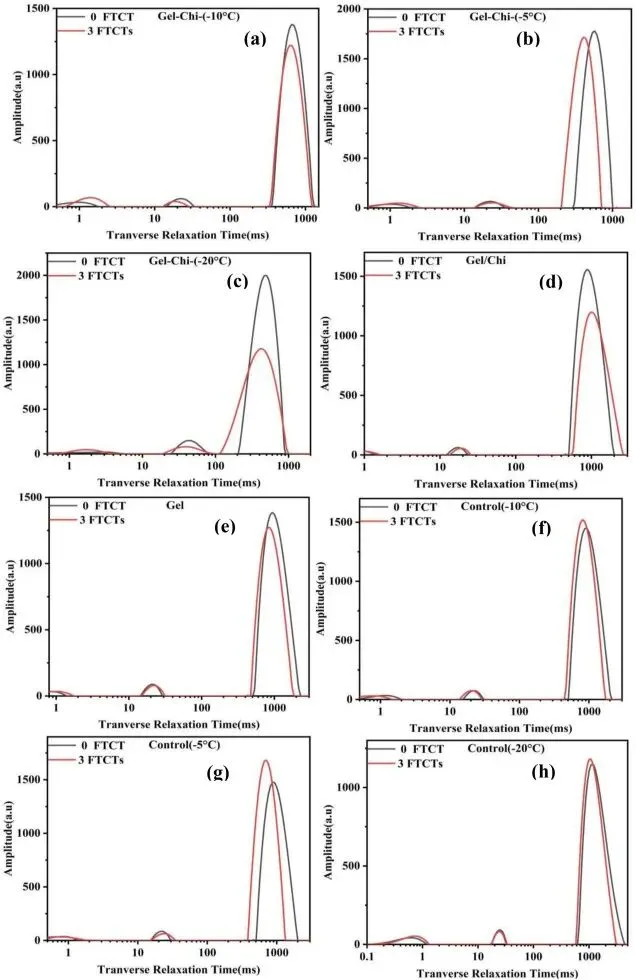
Fig. 6. LF-NMR proton density images of glycosylated gelatin, control group, gelatin, and gelatin/chitosan mixture before and after freeze-thaw cycles.
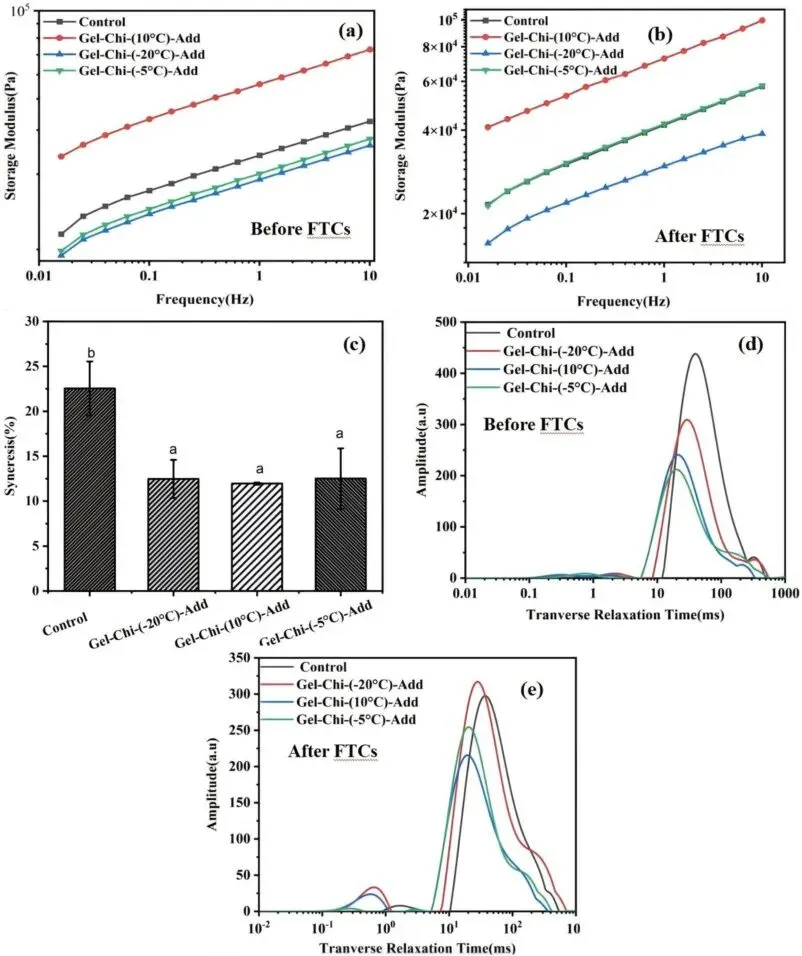
Fig. 7. Frequency sweep (a, b), syneresis (c), and LF-NMR parameters (d–e) of surimi gels fortified with 1% Gel-Chi synthesized at different temperatures (10°C, −5°C, and −20°C) before and after 12 freeze-thaw cycles (FTC).
Fig. 8. Visual appearance and microstructure (SEM) of surimi gels fortified with 1% Gel-Chi synthesized at different temperatures (10°C, −5°C, and −20°C) before and after 12 freeze-thaw cycles (FTC).
https://doi.org/10.1016/j.foodhyd.2026.113025
免责声明:「原创」仅代表原创编译,水平有限,仅供学术交流,本平台不主张原文的版权,如有侵权,请联系删除。文献解读或作者简历如有疏漏之处,我们深表歉意,请作者团队及时联系《食探未来》主编(微信号:shitanweilai8077),我们会在第一时间进行修改或撤稿重发,感谢您的谅解!