研究背景
粮食安全是全球面临的重大挑战。水稻作为世界上最重要的主食作物之一,其产量的提升在很大程度上取决于穗粒数(GNPP)。穗粒数是一个受遗传、环境以及植物能量供应共同影响的复杂性状。植物作为固着生长生物,必须在不断变化的能量状况(如光照强度、养分分布)下灵活调节自身的生长发育。
SnRK1(蔗糖非发酵1相关蛋白激酶1)复合物被公认为植物细胞内的“能量传感器”。当能量供应充足时,SnRK1活性受到抑制以促进生长;当能量匮乏时,它被激活,开启分解代谢(能量回收)并抑制合成代谢(能量消耗)。SnRK1是一个异源三聚体复合物,由一个催化亚基(α亚基)和两个调节亚基(β和γ亚基)组成。尽管在模式植物拟南芥中,SnRK1亚基的功能已有深入研究,但在作物,尤其是水稻中,能量感知如何与生殖器官发育(如穗粒数的形成)协调一致的分子基础仍未完全解析。

论文概要
山西农业大学王拴锁、孔照胜团队联合中国科学院遗传与发育生物学研究所傅向东团队在Molecular Plant发表了题为“Dual repression of OsSnRK1β1A by the deubiquitinase OsOTUB1 orchestrates energy metabolism and grain yield in rice”的论文。该研究通过遗传筛选、生化解析及育种应用,揭示了由去泛素化酶OsOTUB1和SnRK1β亚基OsSnRK1β1A构成的双重抑制调节模块,阐明了其如何通过重塑碳、氮、脂代谢来协同控制能量稳态与穗发育的机制。该研究不仅为能量代谢与穗粒数之间的平衡提供了理论框架,还通过精准编辑启动子创制了高产水稻种质资源。
主要研究结果介绍
1. 穗粒数关键调控基因 NPT-S1/OsSnRK1β1A的克隆
研究团队此前通过研究理想株型基因NPT1(编码去泛素化酶OsOTUB1),发现npt1缺失突变体表现出显著增加的穗粒数。为了寻找其下游组分,研究者对npt1突变体进行了EMS诱变处理,筛选到了一个抑制子突变体npt-s1,它能够完全恢复(抵消)npt1穗粒数增加的表型,使其回到野生型中花11(ZH11)的水平(图1A-B)。
利用图位克隆方法,研究人员将候选基因定位在5号染色体一个15 kb的区间内,测序发现Os05g41220基因在npt-s1突变体中缺失了两个碱基,导致蛋白质翻译提前终止。该基因编码SnRK1复合物的β亚基,研究将其命名为OsSnRK1β1A。
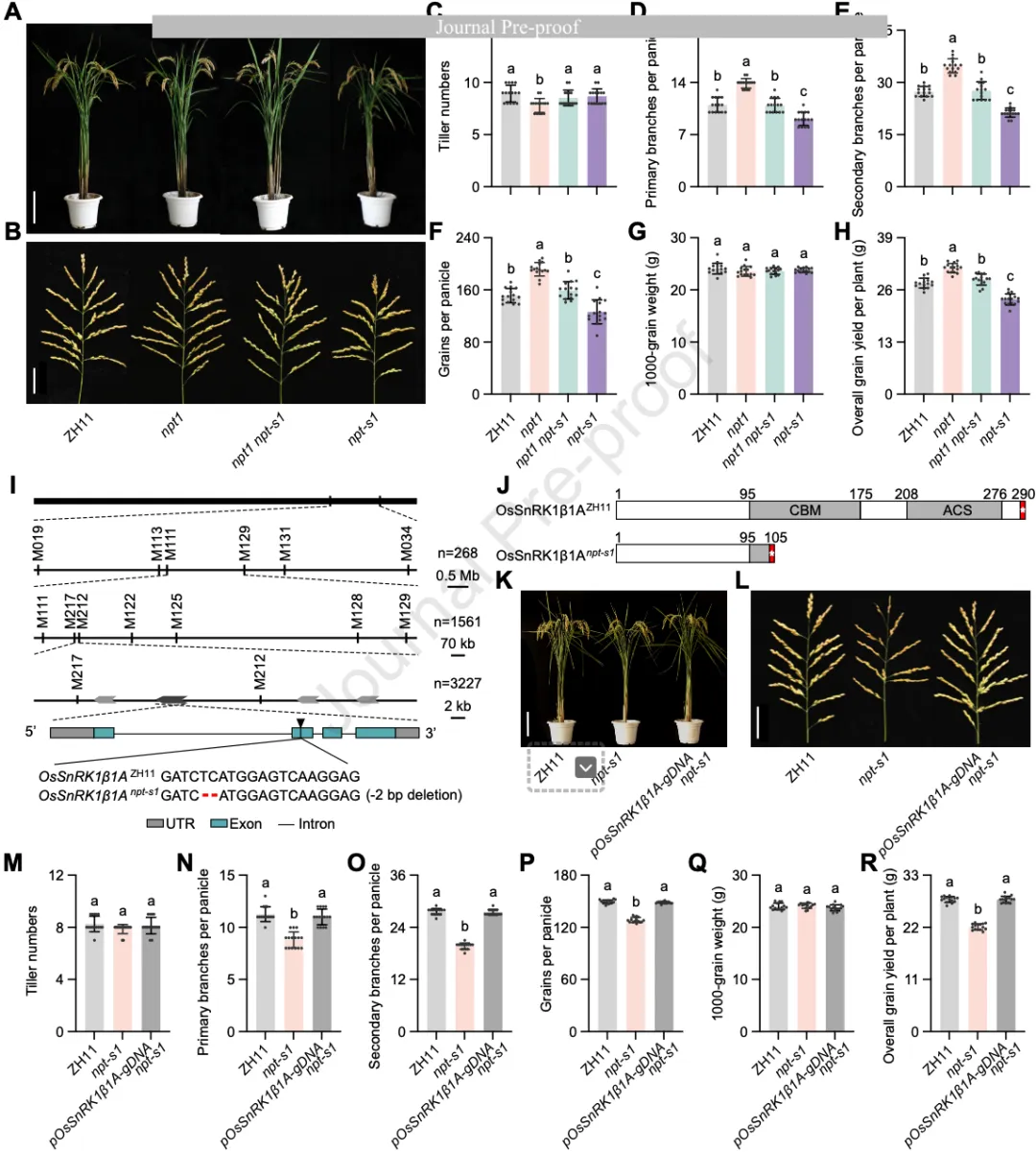 图1
图12. OsSnRK1β1A 正向调节穗粒数并抑制能量分解代谢
功能分析显示,ossnrk1β1a缺失突变体表现穗分枝和穗粒数显著减少(图2A-H)。相反,过表达OsSnRK1β1A的植株穗粒数和穗分枝明显增加,且单株产量较野生型提升了约8%(图2H)。
有趣的是,OsSnRK1β1A的功能与经典SnRK1α亚基在压力下的作用相反:它在正常生长条件下通过抑制能量分解代谢来促进穗部发育。转录组分析显示,OsSnRK1β1A能够显著抑制与分解代谢相关的基因(如脂质分解基因OsGDSL1、氨基酸分解基因OsDIN9等)的表达。在持续黑暗处理(能量胁迫)下,OsSnRK1β1A过表达品系表现出更严重的失绿和生物量损失,而突变体则表现出更强的耐受性(图2I-K),进一步确证了其在能量平衡中的核心地位。
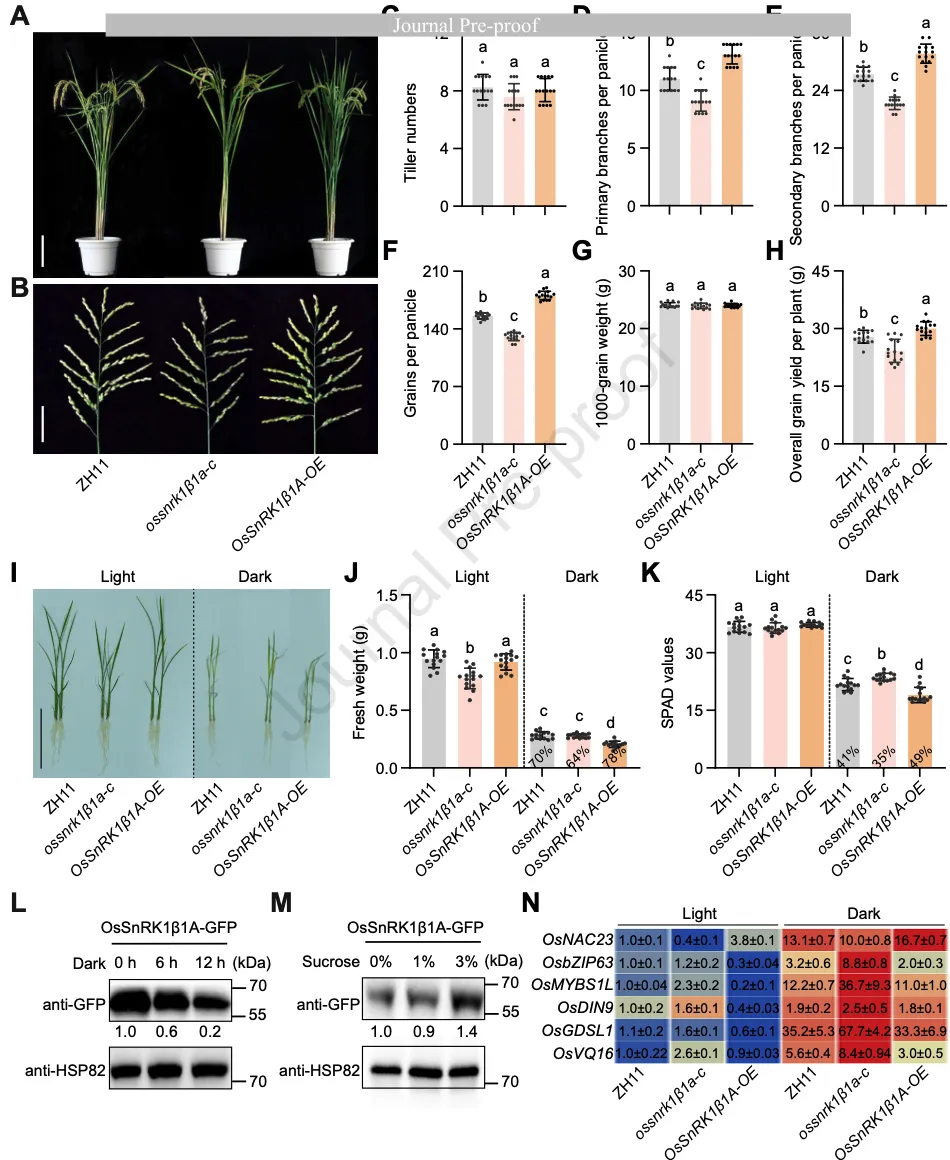 图2
图23. 肉豆蔻酰化介导的“胞质隔离”机制
生化实验(Y2H, BiFC, Co-IP)证明,OsSnRK1β1A能与所有三个催化亚基(α1, α2, α3)相互作用。机制研究发现,OsSnRK1β1A并不直接改变α亚基的磷酸化状态,而是通过一种“空间限制策略”发挥作用(图3A-G)。
OsSnRK1β1A蛋白具有一个保守的N-末端肉豆蔻酰化位点(Gly2)。研究发现,正常的OsSnRK1β1A通过肉豆蔻酰化作用将α亚基(如OsSnRK1α1)锚定在细胞质中形成点状结构,从而限制其进入细胞核。在ossnrk1β1a突变体中,α亚基向核内转移,激活了分解代谢基因。若将Gly2突变为Ala(G2A)导致肉豆蔻酰化失活,OsSnRK1β1A将失去将α亚基滞留在胞质的能力,进而导致穗发育表型与野生型无异(图3H-N)。这一发现揭示了植物通过蛋白质的翻译后修饰来动态分配能量传感器的亚细胞分布,从而耦合能量状态与发育输出。
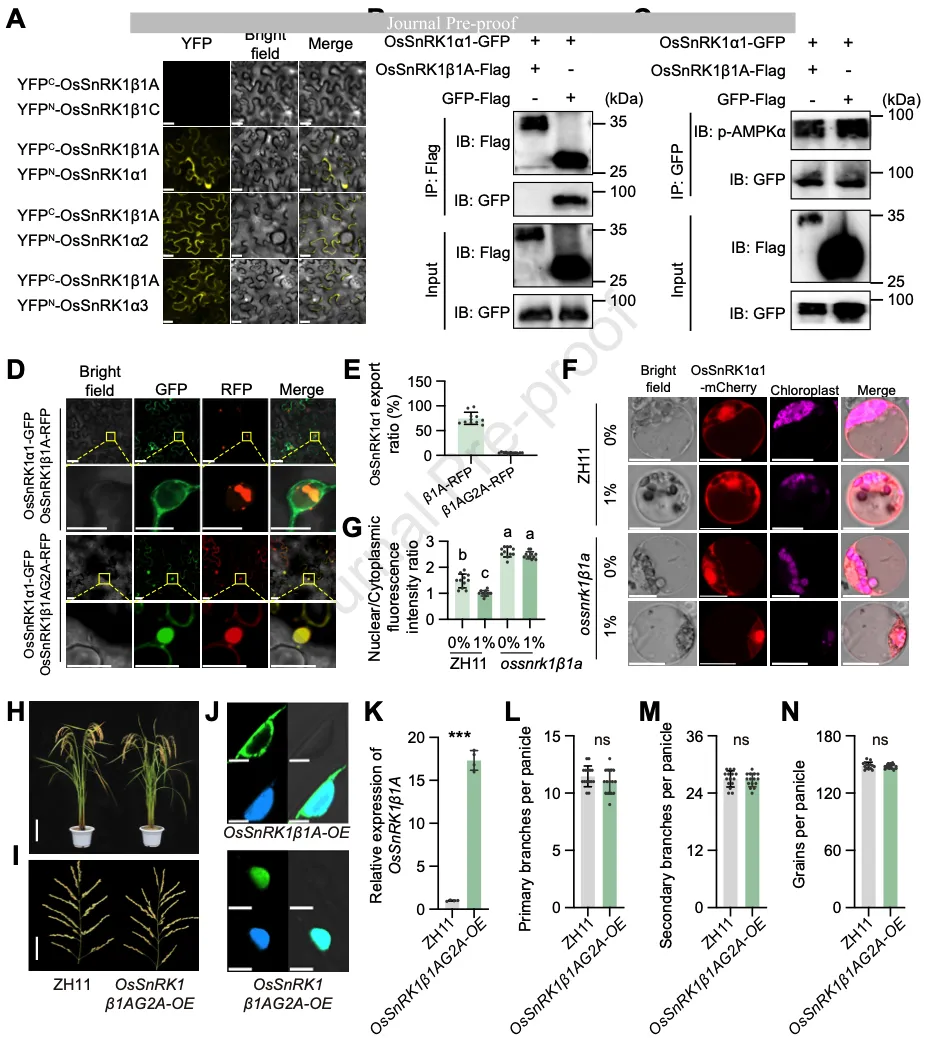 图3
图34. 去泛素化酶 OsOTUB1 对 OsSnRK1β1A 的双重抑制
研究进一步揭示了穗粒数调控因子OsOTUB1是如何负向调节OsSnRK1β1A的。实验证明,OsOTUB1对OsSnRK1β1A具有双重抑制效应(图6):
- 竞争性抑制: OsOTUB1与催化亚基OsSnRK1α1结合OsSnRK1β1A的同一个结构域(ACS域)。因此,当OsOTUB1含量增加时,它会竞争性地从OsSnRK1β1A-OsSnRK1α1复合物中解离出α亚基,使其重新入核。
- 泛素链编辑驱动的稳定性调节: 这是一个非经典的调节模式。OsSnRK1β1A同时具有K48和K63连接的泛素化。OsOTUB1作为去泛素化酶,并不直接稳定底物,而是特异性地切除OsSnRK1β1A上的K63连接泛素链。由于K63链在该通路中起保护作用,K63链的切除导致OsSnRK1β1A更容易受到K48泛素化修饰,从而触发26S蛋白酶体介导的降解(图6G)。
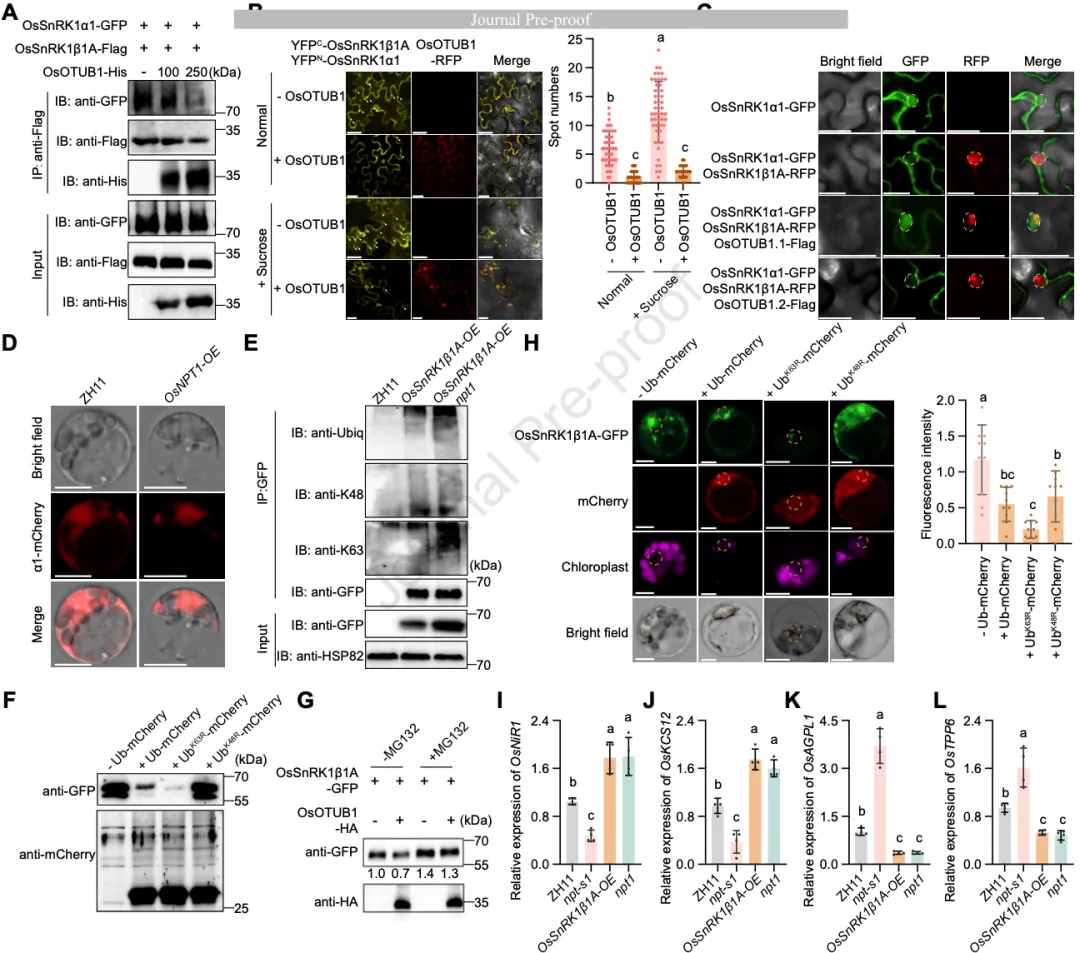 图6
图6能量状态动态调节这一轴线:在黑暗(能量低)下,OsOTUB1水平升高,加速OsSnRK1β1A的降解并释放α亚基进入核内,开启节能模式;而在充足光照或高糖环境下,OsOTUB1受到抑制,OsSnRK1β1A积累并将α亚基滞留在胞质,优先保障产量形成(图5E-H)。
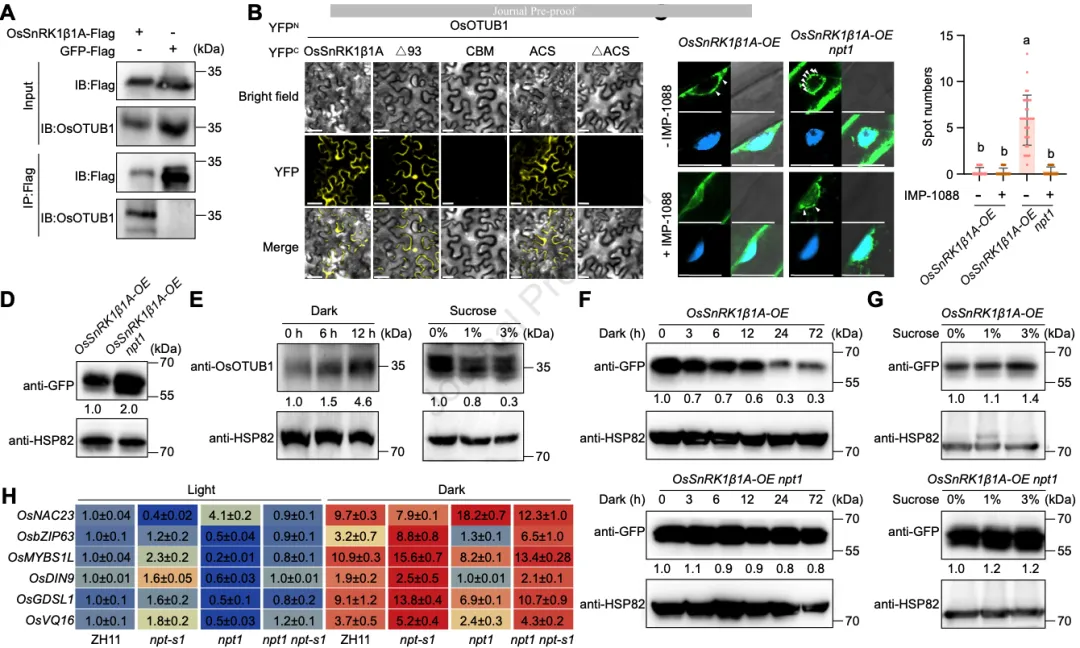 图5
图55. 优异单倍型挖掘与启动子精准编辑育种
研究者分析了266份水稻资源的序列变异,鉴定出了两个主要单倍型(Hap1和Hap2)。其中Hap2(主要存在于籼稻和澳洲稻中)具有更高的穗粒数和更多的二级分枝(图7A-D)。通过构建近等基因系,证明Hap2的优势源于其编码区的氨基酸差异。
为了进一步挖掘该基因的增产潜力,研究团队利用CRISPR/Cas9技术针对OsSnRK1β1A启动子中的负调控元件(CAREOSREP1和CCAAT-box)进行了精准编辑。结果显示,启动子编辑品系(pro2和pro5)在不改变植株基本构型(如株高、分蘖数)的前提下,显著提高了OsSnRK1β1A的转录水平,进而显著增加了水稻的穗粒数和穗分枝,展现了极佳的分子育种应用前景(图7L-Q)。
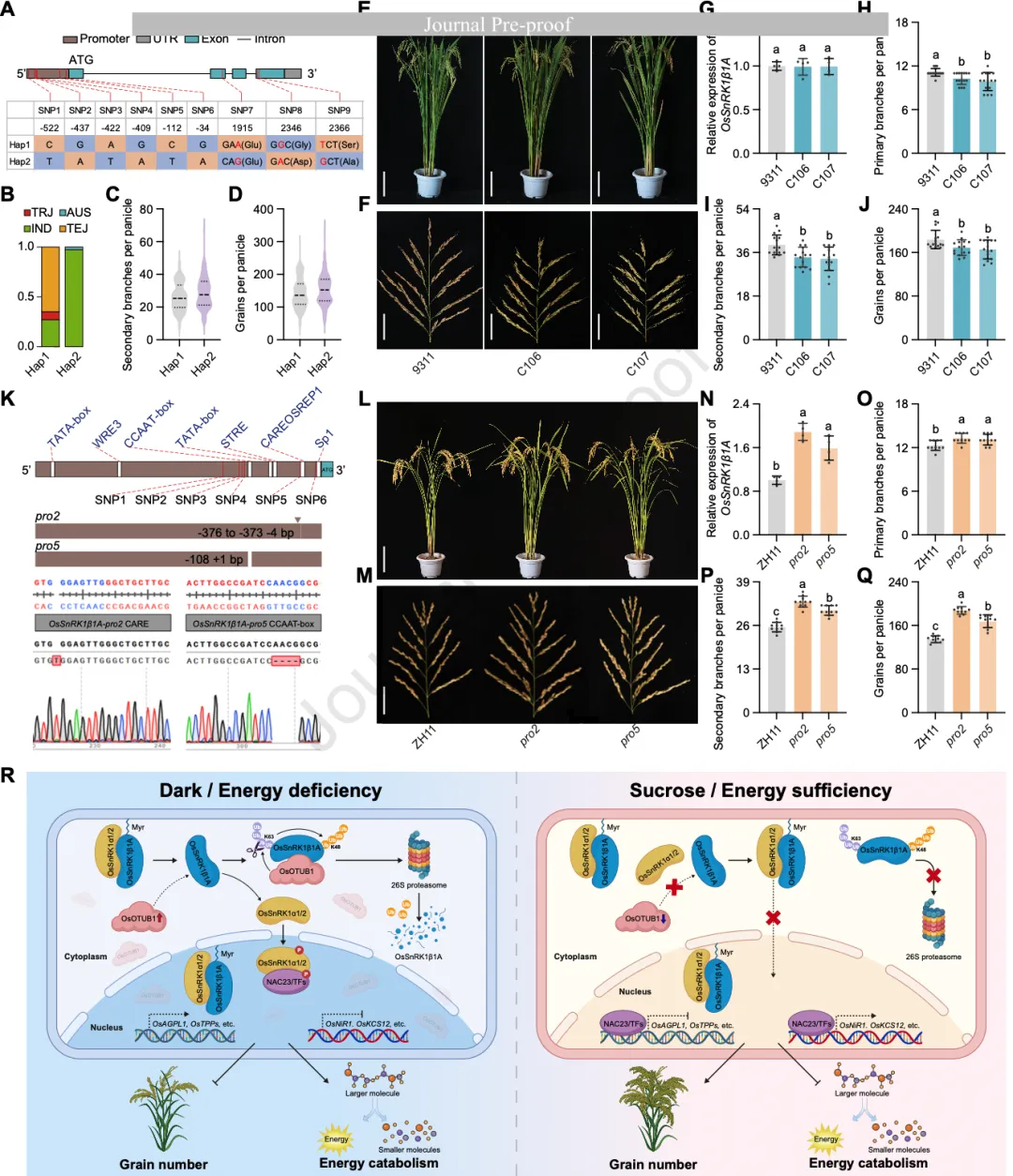 图7
图7全文总结与展望
本研究构建了一个精细的水稻穗粒数调控网络:OsOTUB1-OsSnRK1β1A-OsSnRK1α1/2。
- OsSnRK1β1A是一个正向调控穗发育的中心枢纽,它通过N-末端肉豆蔻酰化将催化亚基(SnRK1α)拦截在胞质中,从而抑制分解代谢,将能量资源导向生殖生长。
- 上游的OsOTUB1作为“控制杆”,通过竞争性结合和非典型去泛素化修饰(泛素链编辑)双重抑制OsSnRK1β1A的活性。
- 该通路敏锐地响应植物的能量状况,实现了“能量感知-分解代谢-穗部发育”的高度集成。
这一发现不仅在理论上重新定义了SnRK1复合物在作物生殖发育中的调控逻辑,还提供了一个通用的高产育种策略:即通过编辑启动子负调控元件来精准上调关键代谢调节基因。这种策略相较于引入自然等位基因,具有更强的背景适应性和操作灵活性。
研究团队与资助
该研究主要由山西农业大学农学院、省部共建有机旱作农业国家重点实验室(筹)和中国科学院遗传与发育生物学研究所合作完成。山西农业大学Ludan Zhang为论文第一作者。山西农业大学孔照胜教授、中科院遗传发育所傅向东研究员以及山西农业大学王拴锁教授为论文通讯作者。
该研究得到了国家自然科学基金、山西省“后稷实验室”科研项目、山西省基础研究计划以及中国科学院种子创新重点实验室等项目的资助。
DOI链接:https://doi.org/10.1016/j.molp.2026.06.007
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
