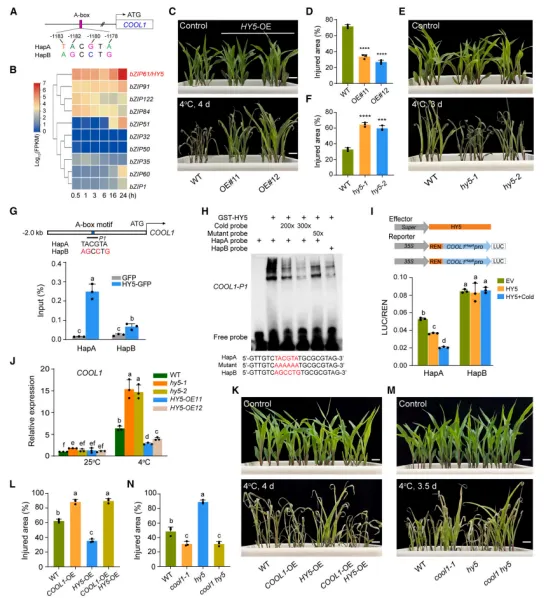
全基因组关联分析(GWAS)在玉米第3染色体上鉴定到一个显著关联峰(图1A)。进一步定位发现,该峰内的5个SNP均位于bHLH转录因子基因 COOL1 的启动子区(图1B)。通过重测序和连锁不平衡分析,确认了9个显著SNP,并根据它们将205份自交系分为HapA(耐冷)和HapB(冷敏感)两种单倍型。HapB型株系的相对叶损伤面积显著高于HapA型(图1D),且 COOL1 表达水平也更高(图1E)。构建近等基因系NIL‑COOL1^HapA 表现出更强的耐冷性和更低的 COOL1 表达(图1F–1H)。过表达 COOL1 降低耐冷性,而CRISPR敲除突变体 cool1 则显著提高耐冷性(图1I–1M)。结果表明,COOL1 是玉米苗期耐冷的负调控因子。序列分析发现,最强的四个SNP位于一个A-box(TACGTA)基序中,该基序是bZIP转录因子HY5的结合位点(图2A)。冷应激下 HY5 表达显著上调(图2B)。过表达 HY5 增强玉米耐冷性,而 hy5 突变体则对冷更敏感(图2C–2F)。ChIP‑qPCR和EMSA显示,HY5与 COOL1^HapA 启动子的结合能力显著强于与 COOL1^HapB 的结合(图2G–2H)。双荧光素酶报告实验表明,HY5能显著抑制HapA型启动子活性,尤其冷处理后更明显,但对HapB型启动子无显著影响(图2I)。遗传分析证实,HY5负调控 COOL1 表达,且 cool1 hy5 双突变体表型与 cool1 单突变体类似(图2J–2N)。综上,HY5通过抑制 COOL1 转录正向调控耐冷性。
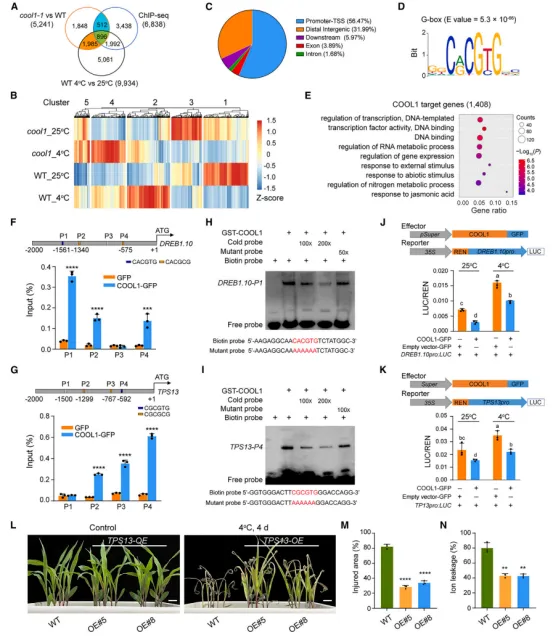
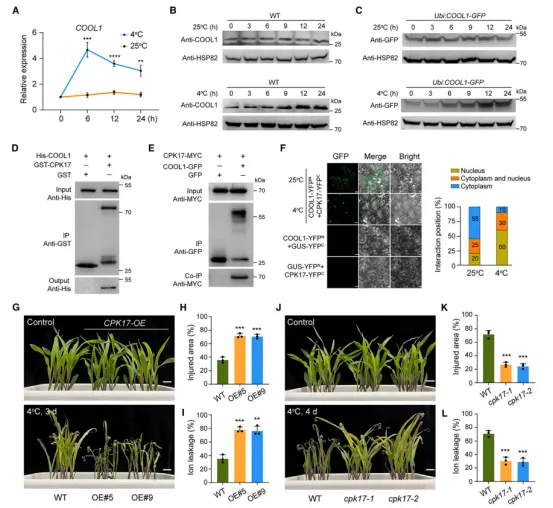
图2 COOL1 启动子自然变异影响HY5的结合亲和力RNA-seq和ChIP-seq联合分析鉴定出1,408个COOL1直接靶基因(图3A)。GO富集显示这些基因参与温度应答、转录调控等过程(图3E)。COOL1结合基序为G-box(图3D)。ChIP-seq发现COOL1结合到 DREB1.10 和 TPS13 的启动子区(图3F–3G),EMSA证实直接结合(图3H–3I)。双荧光素酶实验显示COOL1抑制这两个基因的启动子活性(图3J–3K)。过表达 TPS13 提高玉米耐冷性(图3L–3N)。因此,COOL1通过抑制 DREB1 和 TPS 等正调控因子的表达来负调控耐冷性。冷处理诱导 COOL1 转录及蛋白积累(图4A–4C)。通过LCMS/MS鉴定到COOL1互作蛋白CPK17,酵母双杂交、pulldown和CoIP证实二者互作(图4D–4E)。BiFC显示互作在常温下主要发生于细胞质,冷处理后向细胞核转移(图4F)。过表达 CPK17 降低耐冷性,而 cpk17 突变体则增强耐冷性(图4G–4L)。表明CPK17是耐冷性的负调控因子。
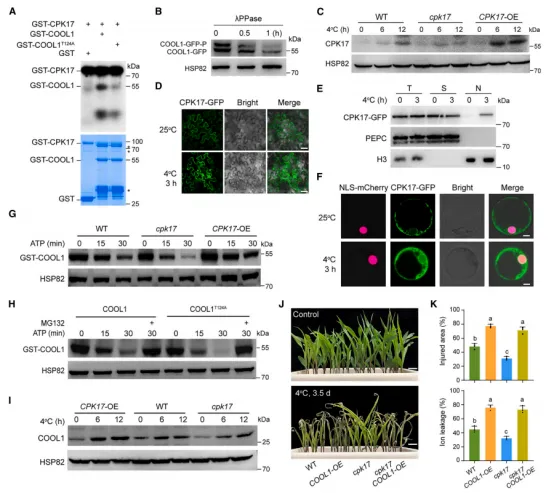
图4 CPK17 与COOL1互作并降低玉米耐冷性体外磷酸化实验显示CPK17可直接磷酸化COOL1,且Thr124为关键位点(图5A)。体内去磷酸化处理证实COOL1被磷酸化(图5B)。冷激活的CPK17激酶活性在 CPK17OE 中增强(图5C)。亚细胞定位显示,冷处理后CPK17从细胞质转入细胞核(图5D–5F)。细胞无细胞降解实验表明,CPK17介导的磷酸化减缓COOL1降解,而T124A突变体降解更快且被MG132抑制(图5G–5H)。免疫印迹进一步证实CPK17稳定COOL1蛋白(图5I)。遗传互作分析显示,cpk17 能部分恢复 COOL1OE 的冷敏感表型(图5J–5K)。说明CPK34通过磷酸化稳定COOL1,从而负调控耐冷性。
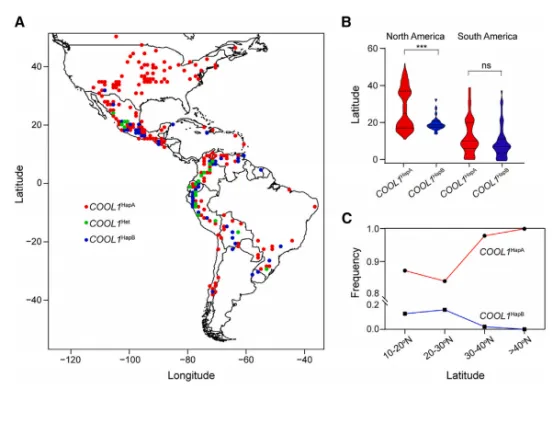
图5 CPK17在冷胁迫下入核,通过磷酸化稳定COOL1对59份大刍草测序发现,约60%的 parviglumis 和47%的 mexicana 携带耐冷型 COOL1^HapA(图6A–6B)。对1008份美洲玉米地方种的基因分型显示,COOL1^HapA 的频率随纬度升高而显著增加,在美国几乎固定;而 COOL1^HapB 主要分布于低纬度地区(图6C)。这表明 COOL1^HapA 在玉米向北方高纬度地区的适应过程中发挥了重要作用。
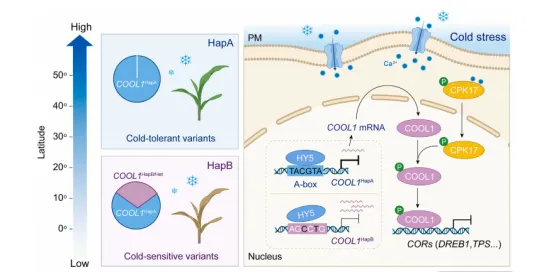
图6 COOL1^HapA 等位基因主要分布于北半球高纬度地区该模型概括了本研究的核心机制:冷胁迫激活CPK17并使其转位至细胞核,CPK17磷酸化COOL1的Thr124位点,增强其蛋白稳定性;稳定的COOL1结合到下游靶基因(如 DREB1、TPS)的启动子并抑制其表达,从而负调控玉米耐冷性。同时,COOL1 启动子区的自然变异(HapA vs HapB)影响了HY5的结合效率:HY5更高效地结合HapA型启动子,抑制 COOL1 转录,导致HapA株系中COOL1表达量低、耐冷性强。群体分布图显示,HapA等位基因在北方高纬度地区占据主导,为玉米适应寒冷气候提供了遗传基础。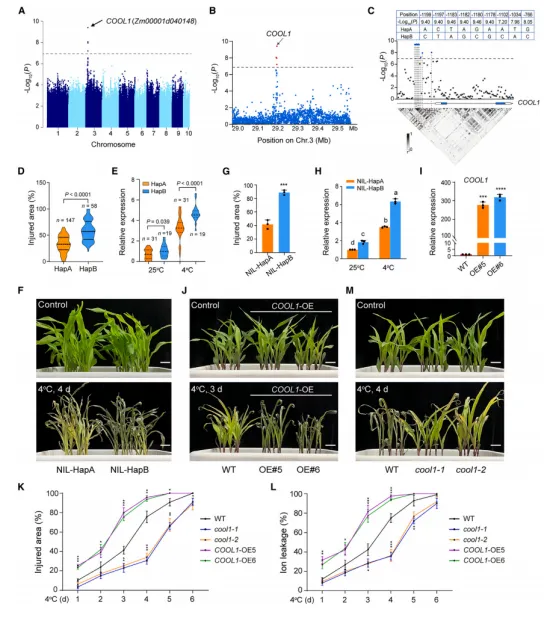
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
