Food Hydrocolloids| 新型产麦芽六糖淀粉酶与 4,6-α- 葡萄糖基转移酶协同酶法策略全面提升面包品质与消化特性
近日,中国农业大学团队在《Food Hydrocolloids》期刊上发表了题为《A synergistic enzymatic strategy for comprehensive improvement of bread quality and digestibility using a novel maltohexaose-forming amylase and a 4,6-α-glucanotransferase》的研究性论文(一区,IF:12.4)。该研究首次鉴定并在毕赤酵母中高效表达了来自噬珊瑚粘球菌的新型产麦芽六糖淀粉酶 CsMFA,探究了其与罗伊氏乳杆菌来源的 4,6-α- 葡萄糖基转移酶 LrN1 GtfB 联用对面包品质的协同调控作用,分析了酶处理对面包质构、抗老化特性及淀粉消化特性的影响,揭示了双酶体系可通过高效水解生淀粉、优化淀粉链长分布、提升支化度,使面包比容提升 32.6%、硬度降低 48.3%,4℃贮藏 4 天硬度降幅达 46.4%,同时显著降低快消化淀粉占比、提升慢消化淀粉含量,为清洁标签面包的品质综合优化与营养升级提供了高效的酶法协同策略。
面包作为全球消费最广泛的主食之一,是膳食能量、蛋白质与营养素的重要来源,但其产业发展长期面临两大核心瓶颈:一是贮藏过程中淀粉回生引发的面包老化问题,表现为风味流失、面包瓤硬化、适口性下降,严重缩短产品货架期;二是面包属于高血糖指数食品,精制小麦淀粉的快速消化易引发餐后血糖与胰岛素的剧烈波动,长期摄入会增加胰岛素抵抗、2 型糖尿病等代谢疾病的发病风险,因此,延缓面包老化、调控淀粉消化特性是提升面包感官品质与营养属性的核心研究方向。淀粉的酶法改性是契合食品行业清洁标签趋势的高效绿色手段,其中产麦芽低聚糖淀粉酶可通过水解淀粉生成麦芽低聚糖,增强体系保水性以抑制淀粉回生、延缓面包老化,但其对淀粉消化特性的调控效果十分有限;而 4,6-α- 葡萄糖基转移酶可通过转糖基作用在淀粉链上引入 α-1,6 糖苷键支链,通过空间位阻阻碍消化酶与底物的接触,有效提升淀粉抗消化性,但其对天然生淀粉的催化活性极低,单独应用甚至可能损害面包的质构品质。此外,目前已报道的产麦芽六糖淀粉酶最适温度普遍高于面包面团发酵的常规温度(27-38℃),野生菌株的酶表达水平低,难以满足工业化应用需求,且产麦芽六糖淀粉酶与 4,6-α- 葡萄糖基转移酶在面包体系中的协同应用,及其对面包品质、淀粉结构与消化特性的调控机制,此前尚未见相关报道。基于此,本研究首次鉴定了来源于噬珊瑚粘球菌的新型产麦芽六糖淀粉酶 CsMFA,实现了其在毕赤酵母中的高效异源表达,系统解析了其酶学特性与水解规律,并将其与罗伊氏乳杆菌来源的高活性 4,6-α- 葡萄糖基转移酶 LrN1 GtfB 联用,探究了二者对面包质构、抗老化特性、淀粉消化特性的协同调控效应,同时以小麦淀粉为模型阐明了双酶体系修饰淀粉的分子机制,旨在为高品质、慢消化、长货架期面包的生产,提供一套高效、绿色的清洁标签酶法解决方案。
明确了新型产麦芽六糖淀粉酶 CsMFA 的酶学特性与工业化应用潜力
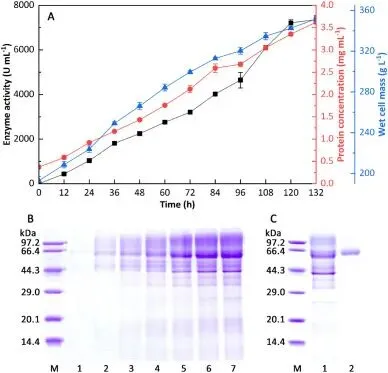
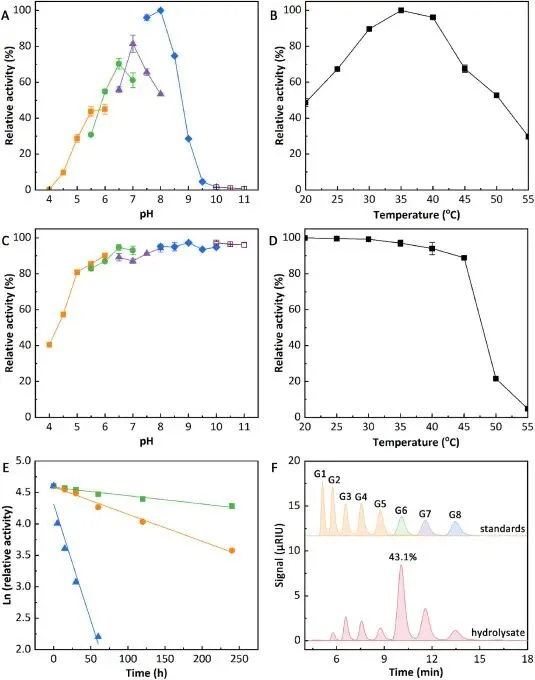
本研究成功在毕赤酵母中实现了 CsMFA 的高效异源表达,经 5 L 发酵罐高密度甲醇诱导 132 h 后,发酵上清液酶活可达 7362.2 U/mL,纯化后比酶活达 4001.9 U/mg,表达量与催化活性均远超已报道的同类产麦芽六糖淀粉酶。该酶最适 pH 为 8.0,在 pH 5.0-11.0 范围内保温 30 min 仍可保留 80% 以上的初始酶活,具备极宽的 pH 稳定范围;最适温度为 35℃,与面包面团发酵的温度条件高度匹配,20℃时仍能保持约 50% 的最大酶活,45℃保温 30 min 酶活保留率超 80%,解决了传统同类酶最适温度过高、面包发酵条件下催化效率低的行业痛点。CsMFA 水解可溶性淀粉的主产物为麦芽六糖,占比达 43.1%,采用 “内切型随机水解 + 外切型定向生成麦芽六糖” 的双模式水解机制,对麦芽六糖无水解活性,具备严格的底物特异性,为其在面包加工中的规模化应用奠定了坚实基础。
揭示了 CsMFA 与 LrN1 GtfB 对面包质构与感官品质的协同增效效应
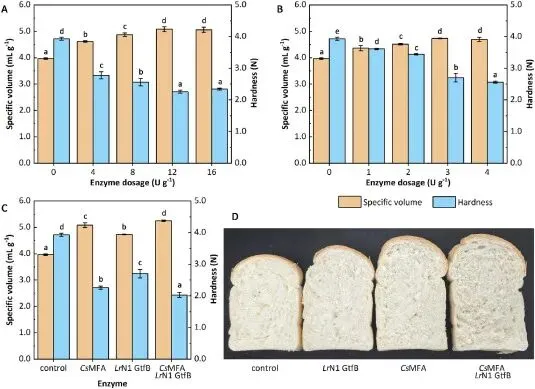
CsMFA 与 LrN1 GtfB 单独使用均可优化面包品质,二者联用则呈现显著的协同提升作用。在最优添加量下,12 U/g CsMFA 单独处理可使面包比容从对照组的 3.96 mL/g 提升至 5.08 mL/g,面包瓤硬度降低 42.5%;3 U/g LrN1 GtfB 单独处理可使面包比容提升 19.4%,硬度降低 31.3%;而双酶联用可使面包比容提升 32.6% 至 5.25 mL/g,硬度降低 48.3% 至 2.03 N,质构优化效果显著优于单一酶处理组与市售商业面包改良酶制剂。微观结构观察显示,双酶处理后的面包瓤气孔更大、分布更均匀,面团持气性与烘焙膨胀性显著提升。其协同机制源于两种酶的催化特性互补:CsMFA 水解生淀粉生成的麦芽低聚糖(尤其是 DP≥6)是 LrN1 GtfB 的优选底物,大幅提升了后者的转糖基催化效率;而 LrN1 GtfB 的转糖基作用进一步优化了面团的网络结构,增强了面团发酵与烘焙过程中的结构稳定性,最终实现了面包比容与质构品质的全面提升。
阐明了双酶体系对面包老化的抑制效应与分子调控机制
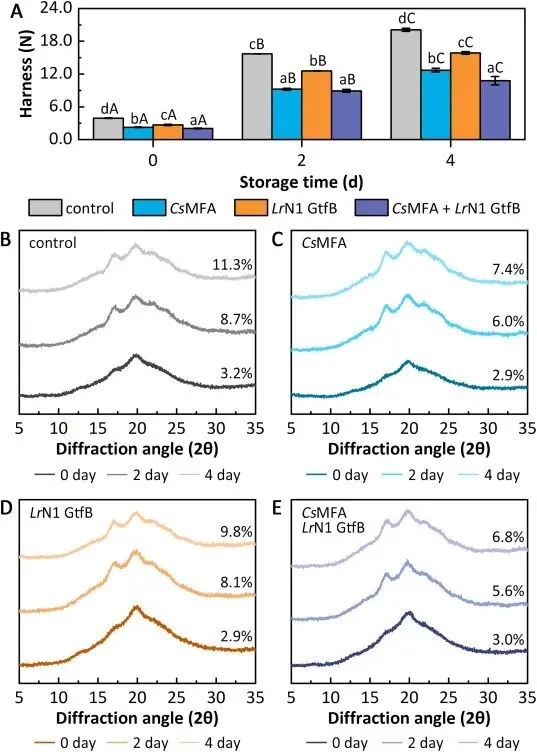
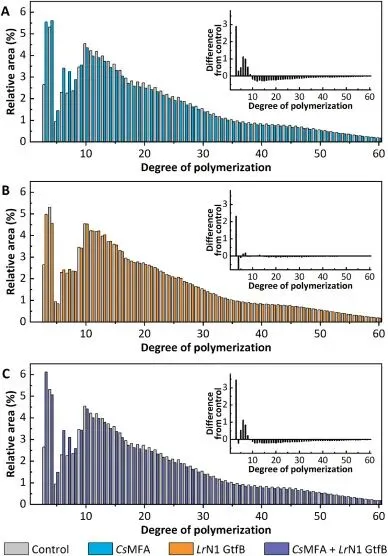
双酶联用可显著延缓面包贮藏过程中的老化进程,4℃冷藏 4 天后,对照组面包硬度从初始的 3.92 N 上升至 20.09 N,增幅达 5.1 倍;而双酶处理组面包贮藏 4 天后硬度仅为 10.77 N,较对照组降低 46.4%,同时面包瓤咀嚼性较对照组、CsMFA 单独处理组、LrN1 GtfB 单独处理组分别降低 41.1%、18.7% 和 29.9%,抗老化效果远超同类酶制剂。X 射线衍射结果显示,新鲜面包淀粉仅呈现 20° 处的 V 型结晶单峰,贮藏 4 天后对照组出现 17° 与 22° 处的 B 型结晶特征峰,相对结晶度升至 11.3%;而双酶处理组贮藏 4 天后相对结晶度仅为 6.8%,显著抑制了淀粉从 V 型向 B+V 混合型结晶的转变。链长分布分析进一步证实,双酶处理使面包淀粉中易引发回生的长链组分(DP 13-24、DP 25-36、DP>36)占比显著降低,而短链(DP 3-5)与中链(DP 6-12)占比分别提升 42.7% 和 6.9%,从根源上削弱了淀粉链重结晶的驱动力;同时 CsMFA 水解生成的麦芽低聚糖具备强保水性,可通过氢键结合阻碍淀粉链间的相互作用,最终实现了面包货架期的有效延长。
证实了双酶联用对面包淀粉消化特性的优化与营养升级效应
双酶体系可实现对面包淀粉消化特性的精准调控,显著提升面包的营养健康属性。对照组面包中快消化淀粉(RDS)占比高达 84.0%,慢消化淀粉(SDS)仅为 13.8%,抗性淀粉(RS)为 2.24%;CsMFA 单独处理可使 RDS 降至 80.6%,SDS 提升至 17.2%;LrN1 GtfB 单独处理可使 RDS 降至 77.9%,SDS 提升至 20.1%;而二者联用实现了最显著的调控效果,使 RDS 大幅降至 72.4%,SDS 占比显著提升至 25.6%,较对照组提升 85.5%。其协同调控机制分为两个维度:一方面,CsMFA 可将面包中的快消化长链淀粉水解为慢消化的麦芽低聚糖,直接降低 RDS 占比;另一方面,LrN1 GtfB 通过转糖基作用在淀粉链上引入大量 α-1,6 糖苷键支链,产生的空间位阻可显著阻碍 α- 淀粉酶、糖化酶等消化酶与淀粉底物的接触,进一步延缓淀粉水解速率。两种酶的协同作用,在不损害面包加工品质的前提下,实现了淀粉消化速率的有效调控,为低升糖指数烘焙食品的开发提供了可行路径。
解析了双酶体系协同修饰小麦淀粉的构效关系与作用机制
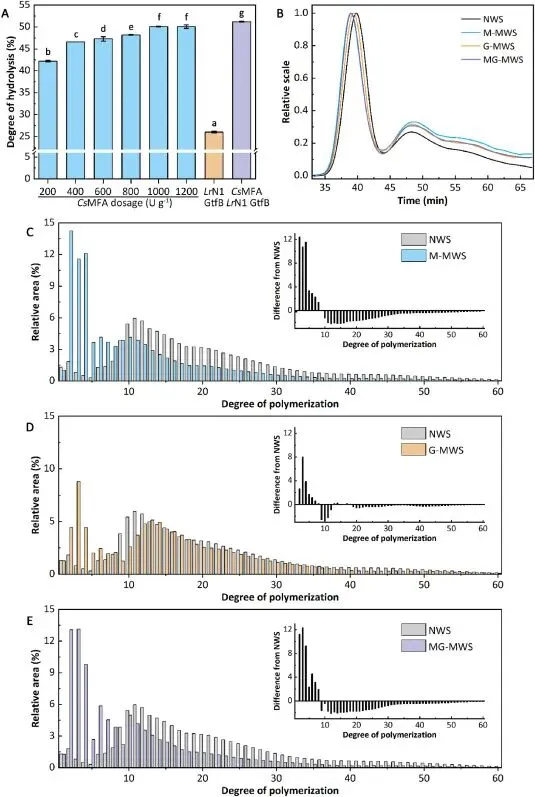
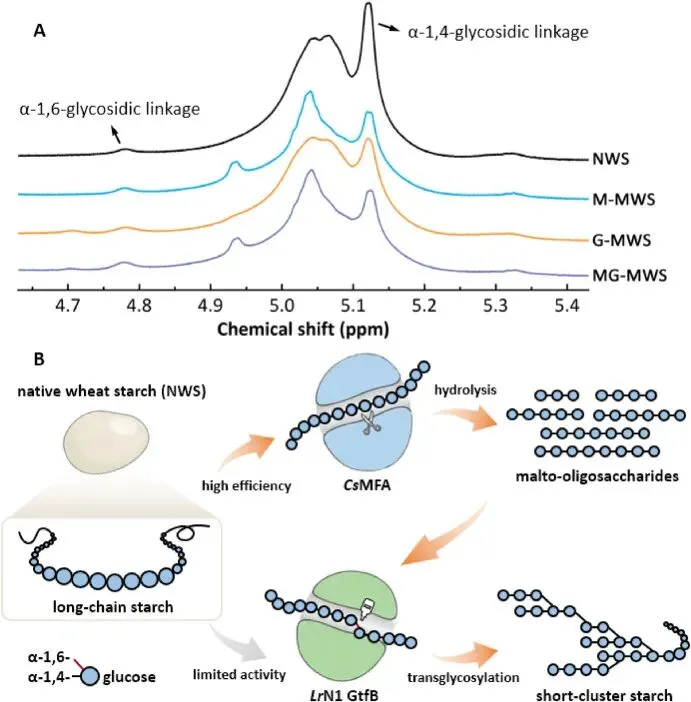
以小麦淀粉为模型的研究,从分子层面阐明了双酶体系协同提升面包品质的核心机制。CsMFA 对天然生淀粉展现出极强的水解能力,最高水解度可达 50.1%,显著弥补了 LrN1 GtfB 对生淀粉催化活性极低的短板,二者联用使淀粉水解度进一步提升至 51.2%。核磁共振结果显示,天然小麦淀粉的支化度仅为 4.6%,CsMFA 与 LrN1 GtfB 单独处理可使支化度分别提升至 6.8% 和 6.9%,而双酶联用使淀粉支化度大幅提升至 8.1%。链长分布分析进一步证实,两种酶具备功能互补性:CsMFA 通过内切水解作用,将长链淀粉降解为大量短链与中链麦芽低聚糖;而 LrN1 GtfB 优先利用这些中短链产物作为底物,通过转糖基反应引入 α-1,6 糖苷键,形成短簇状的支化淀粉结构。这种独特的淀粉结构,一方面通过减少长直链淀粉占比抑制淀粉回生,延缓面包老化;另一方面通过高支化度产生的空间位阻延缓消化酶的催化水解,调控淀粉消化特性;同时优化的链长分布还可改善面团的流变学特性,提升面团持气性与烘焙稳定性,最终实现了面包质构、货架期与营养特性的全面提升。
Fig. 1. High cell density fermentation and purification of CsMFA. (A) Time course of CsMFA expression in K. phaffii. Symbols ■, ●, and ▲ mean enzyme activity, protein concentration, and wet cell mass, respectively. (B) SDS-PAGE analysis of extracellular protein during high cell density fermentation. Lane M, low molecular weight standards; lanes 1–7, fermentation supernatants withdrawn at 0, 24, 48, 72, 96, 120, and 132 h, respectively. (C) SDS-PAGE analysis of CsMFA purification. Lane M, low molecular weight standards; lane 1, crude enzyme; lane 2, purified CsMFA.
Fig. 2. Optimal pH (A), optimal temperature (B), pH stability (C), thermostability (D), thermal denaturation half-lives (E), and hydrolysis property (F) of CsMFA. The optimal pH and pH stability were measured by the standard method in different buffers at 35 and 25 °C, respectively. The buffers included citrate (■), MES (●), MOPS (▲), CHES (◆), and CAPS (□). The optimal temperature and thermostability were measured by the standard method in pH 8.0 at 20–55 °C. For thermal denaturation half-lives, enzyme activities were measured after incubation at 40 °C (■) (y = −0.0012x+ 4.5817, R2 = 0.9656), 45 °C (●) (y = −0.0043x+ 4.5872, R2 = 0.9903), and 50 °C (▲) (y = −0.0371x+ 4.3155, R2 = 0.9550), respectively. For hydrolysis property, 1.0% (w/v) soluble starch was hydrolyzed by CsMFA (1.0 U mL−1) at 30 °C and pH 8.0 for 12 h. The hydrolysate was analyzed by HPLC with glucose (G1) and G2-G8 as standards.
Fig. 3. Effects of enzymatic treatments on bread quality. (A) Specific volume and hardness of bread treated with increasing dosage (0–16 U g−1) of CsMFA. (B) Specific volume and hardness of bread treated with increasing dosage (0–4 U g−1) of LrN1 GtfB. (C) Specific volume and hardness of bread treated with the synergistic combination of CsMFA (12 U g−1) and LrN1 GtfB (3 U g−1). The control received no enzyme. All measurements were performed after baking and cooling at 25 °C for 2 h. Different lowercase letters (a-e) indicate statistically significant differences (p <0.05) in specific volume or hardness among treatments. (D) Representative cross-sectional images of breads from each treatment group.
Fig. 4. Effects of enzymatic treatments on bread staling. (A) Hardness evolution of bread stored at 4 °C up to 4 days. Treatments included control (no enzyme), CsMFA (12 U g−1), LrN1 GtfB (3 U g−1), and the combination of both enzymes. Different lowercase letters (a-d) indicate statistically significant differences (p <0.05) among treatments at the same storage time; uppercase letters (A–C) denote significant differences across storage times within the same treatment. (B-E) XRD patterns of bread stored at 4 °C for 4 days. (B) Control, (C) CsMFA-treated. (D) LrN1 GtfB-treated. (E) combined-enzyme treatment. Relative crystallinity values are shown on the right of each profile.
Fig. 5. Chain length distribution of starch isolated from bread. Panels A, B, and C compare the CLD of starch from the control bread with that from bread treated with CsMFA (12 U g−1), LrN1 GtfB (3 U g−1), and a combination of CsMFA and LrN1 GtfB, respectively. The inset in each graph represents the difference plots, calculated by subtracting the control CLD profile from the corresponding enzyme-treated profile.
Fig. 6. Characterization of modified wheat starch. (A) DH of NWS treated with CsMFA (200–1200 U g−1), LrN1 GtfB (100 U g−1), or their combination (CsMFA 1000 U g−1 and LrN1 GtfB 100 U g−1). Reactions were conducted at 35 °C and pH 7.0 for 12 h. Different lowercase letters (a-g) indicate statistically significant differences (p <0.05) among treatments. (B) Molecular weight distribution of NWS and MWS. M-MWS, G-MWS, and MG-MWS represent starch treated with CsMFA (1000 U g−1), LrN1 GtfB, and the combination of both enzymes, respectively. (C-E) CLD analysis of M-MWS (C), G-MWS (D), and MG-MWS(E). The inset in each graph represents the difference plots, calculated by subtracting the CLD profile of NWS from that of each corresponding MWS.
Fig. 7. Structural modifications of wheat starch by CsMFA and LrN1 GtfB. (A) 1H NMR spectra of NWS and MWS. M-MWS, G-MWS, and MG-MWS were prepared by incubating NWS at 35 °C (pH 7.0) for 12 h with CsMFA (1000 U g−1), LrN1 GtfB (100 U g−1), and their combination (CsMFA 1000 U g−1 and LrN1 GtfB 100 U g−1), respectively. (B) Schematic illustration of the proposed synergistic mechanism of CsMFA and LrN1 GtfB.
https://doi.org/10.1016/j.foodhyd.2026.112838
免责声明:「原创」仅代表原创编译,水平有限,仅供学术交流,本平台不主张原文的版权,如有侵权,请联系删除。文献解读或作者简历如有疏漏之处,我们深表歉意,请作者团队及时联系《食探未来》主编(微信号:shitanweilai8077),我们会在第一时间进行修改或撤稿重发,感谢您的谅解!




 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
