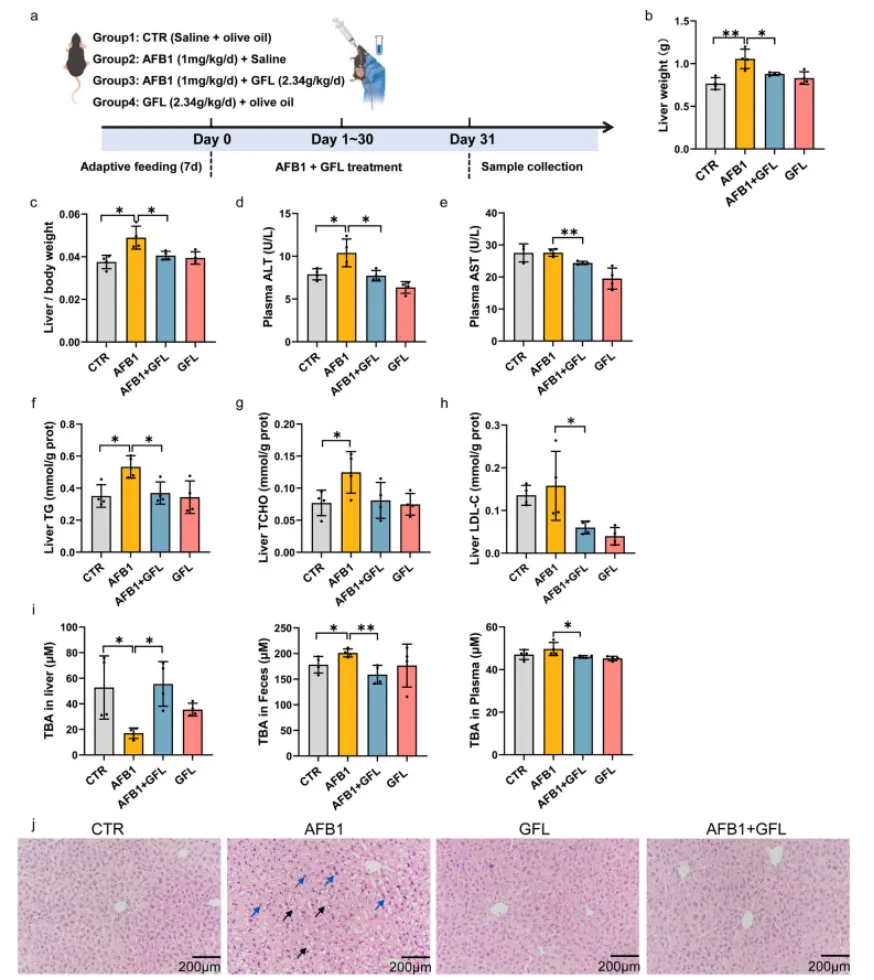
3.1 GFL缓解AFB1诱导的小鼠肝损伤
本研究将C57BL/6小鼠随机分为四组:对照组(CTR)、AFB1组(1 mg/kg/d AFB1)、GFL组(2.34 g/kg/d GFL)和AFB1+GFL组(2.34 g/kg/d GFL + 1 mg/kg/d AFB1),试验持续30天。结果显示,AFB1慢性暴露导致小鼠肝脏重量、肝脏指数显著升高,血浆ALT、AST等肝损伤核心标志物活性大幅上调,肝脏TG、TCHO、LDL-C等脂质代谢指标异常,肝组织出现肝细胞气球样变、核固缩、肝索排列紊乱、炎症细胞浸润等严重病理改变,同时伴随胆汁酸代谢紊乱。而GFL灌胃处理可显著逆转上述异常,恢复肝脏生化指标与组织病理结构,有效改善慢性AFB1暴露诱导的肝损伤(图1)。
图1. GFL减轻AFB1诱导的肝损伤。
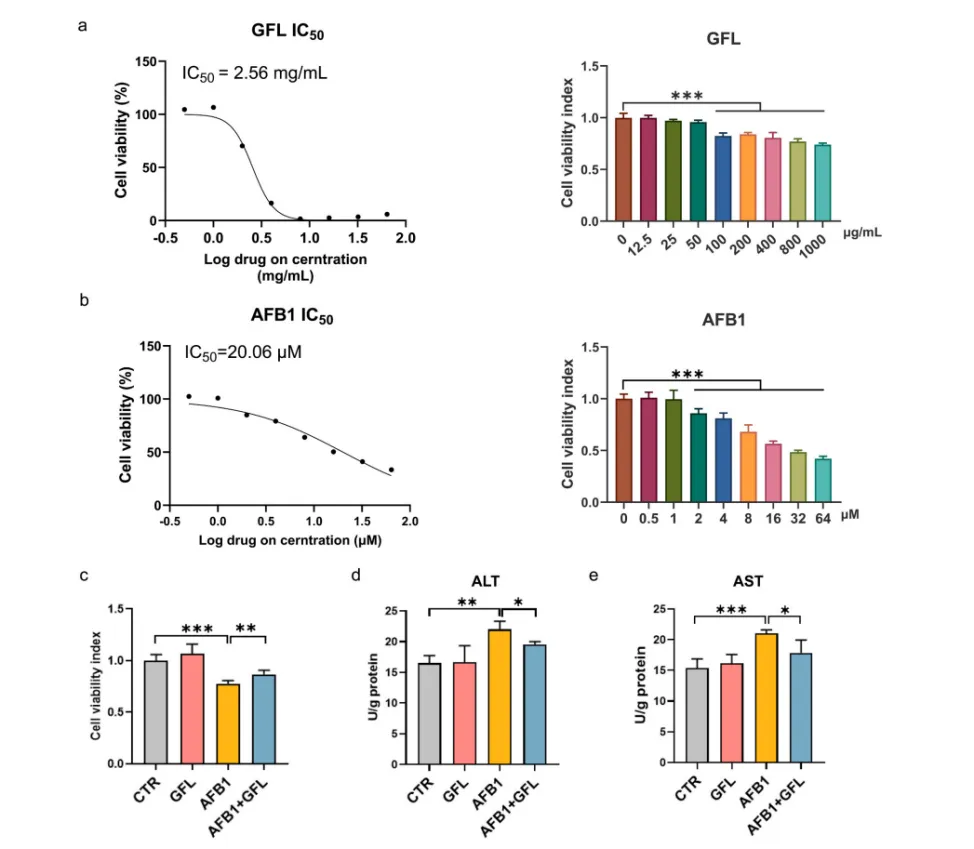
3.2 GFL减轻AFB1诱导的肝细胞毒性
为进一步验证GFL的保肝作用,本研究采用小鼠正常肝细胞AML12开展体外实验。结果显示,AFB1处理可显著降低肝细胞活力,上调细胞内ALT、AST活性,诱发明显的肝细胞毒性;而25 μg/mL GFL预处理可有效抑制AFB1诱导的肝细胞活力下降,降低细胞内转氨酶水平,在细胞层面直接证实了GFL对AFB1诱导肝细胞毒性的保护作用(图2)。
图2. GFL预处理可减轻AFB1诱导的肝细胞毒性。
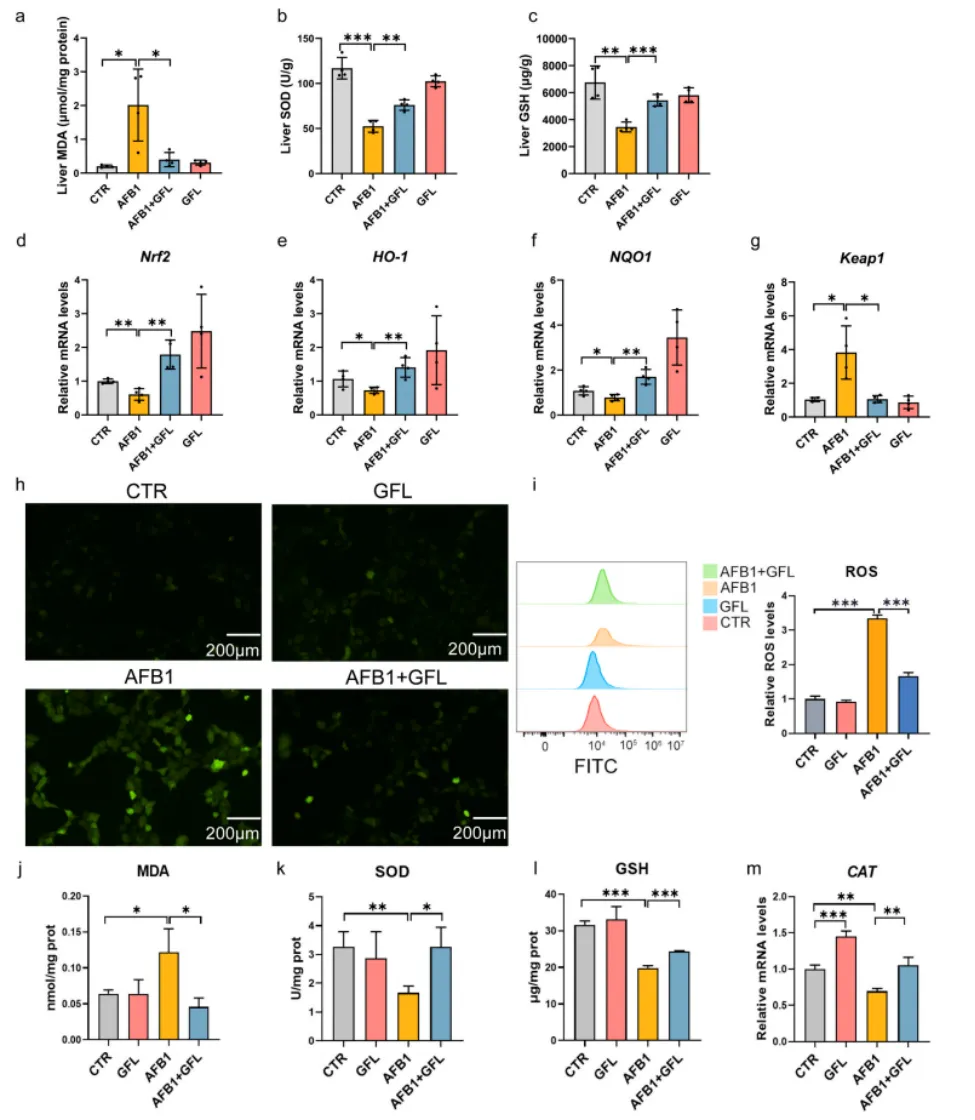
3.3 GFL通过激活Keap1/Nrf2通路减轻AFB1诱导的氧化应激
AFB1暴露显著升高了小鼠肝脏与AML12肝细胞中ROS、脂质过氧化终产物MDA的水平,同时降低了抗氧化酶SOD活性与内源性抗氧化物质GSH的含量,下调了抗氧化通路核心基因Nrf2、HO-1、NQO1的表达,上调了通路抑制因子Keap1的表达。而GFL处理可显著逆转上述氧化应激相关指标的异常,有效激活Keap1/Nrf2抗氧化信号通路,增强肝细胞的内源性抗氧化防御能力,抑制AFB1诱导的氧化应激损伤(图6)。
图6. GFL减轻AFB1诱导的肝细胞氧化应激。
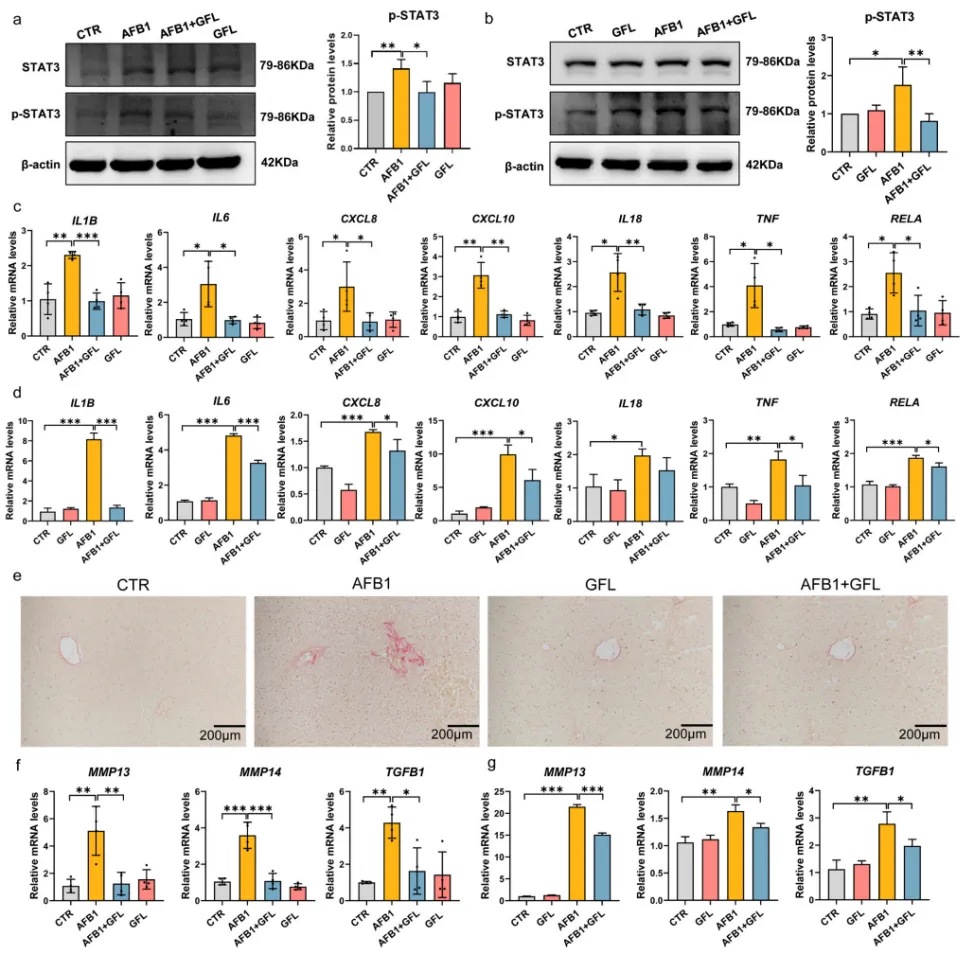
3.4 GFL通过抑制IL6/STAT3通路减轻AFB1诱导的肝脏炎症与纤维化
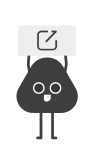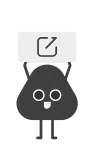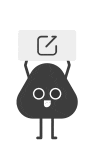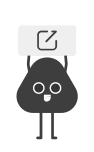
AFB1暴露显著激活了小鼠肝脏与AML12肝细胞中IL6/STAT3炎症信号通路,大幅上调IL1β、IL6、TNF-α等促炎因子的mRNA与蛋白表达,同时升高了MMP13、TGFB1等纤维化相关基因的水平,诱发肝脏纤维化病变。而GFL处理可显著抑制STAT3的磷酸化激活,下调下游促炎因子与纤维化相关基因的表达,有效减轻肝脏炎症浸润与纤维化程度,证实GFL可通过抑制IL6/STAT3通路发挥抗炎抗纤维化作用(图7)。
图7. GFL抑制AFB1诱导的肝脏炎症和纤维化。
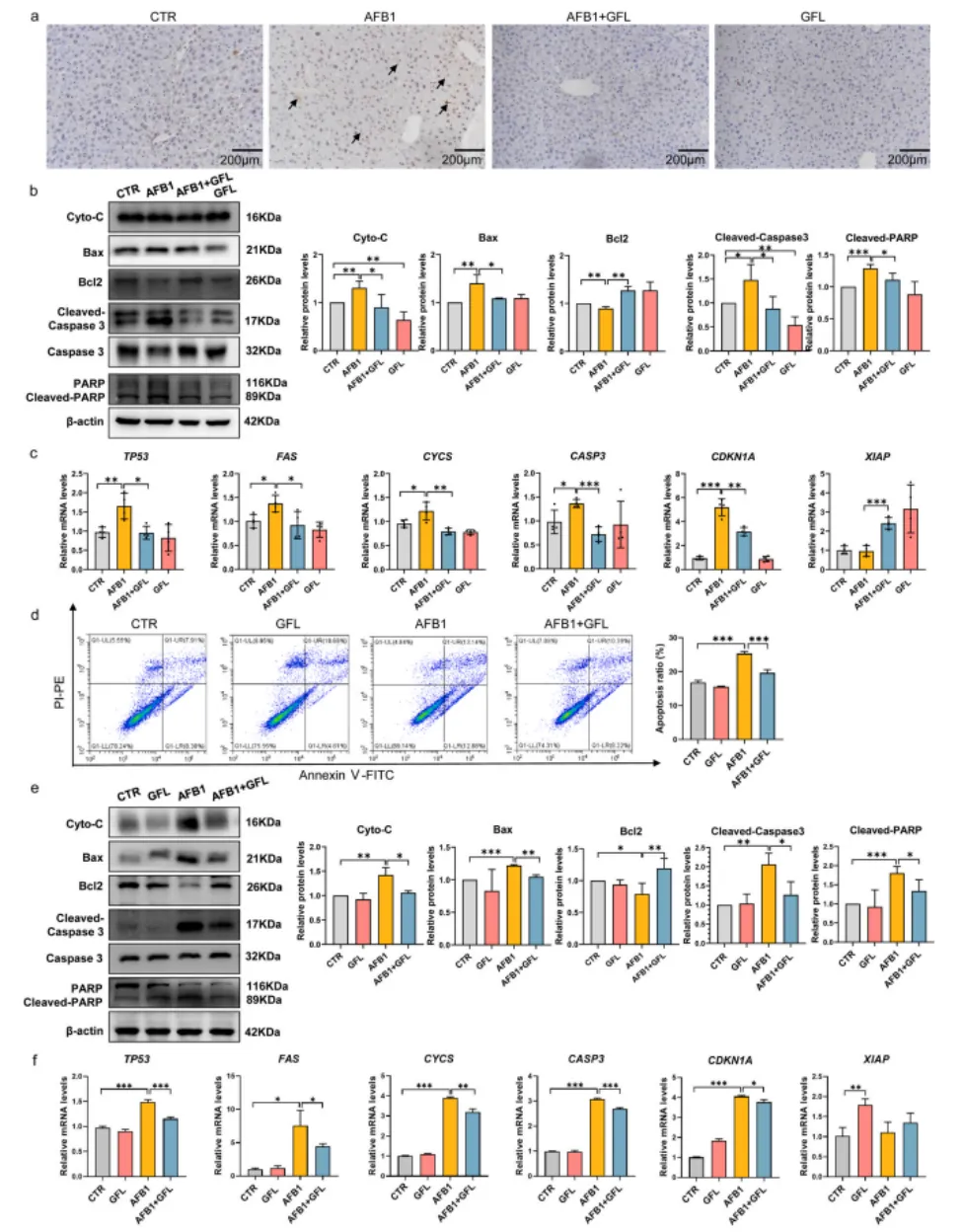
3.5 GFL抑制AFB1诱导的肝细胞凋亡
TUNEL染色与流式细胞术结果显示,AFB1暴露显著增加了小鼠肝脏组织与AML12肝细胞的凋亡率,同时上调了促凋亡蛋白Bax、Cyto-C、Cleaved-Caspase3、Cleaved-PARP的表达,下调了抗凋亡蛋白Bcl2的水平。而GFL处理可显著减少肝细胞凋亡,逆转凋亡相关蛋白与基因的异常表达,通过抑制线粒体依赖的凋亡通路,减轻AFB1诱导的肝细胞凋亡,进而缓解肝损伤(图8)。
图8. GFL抑制AFB1诱导的肝细胞凋亡。
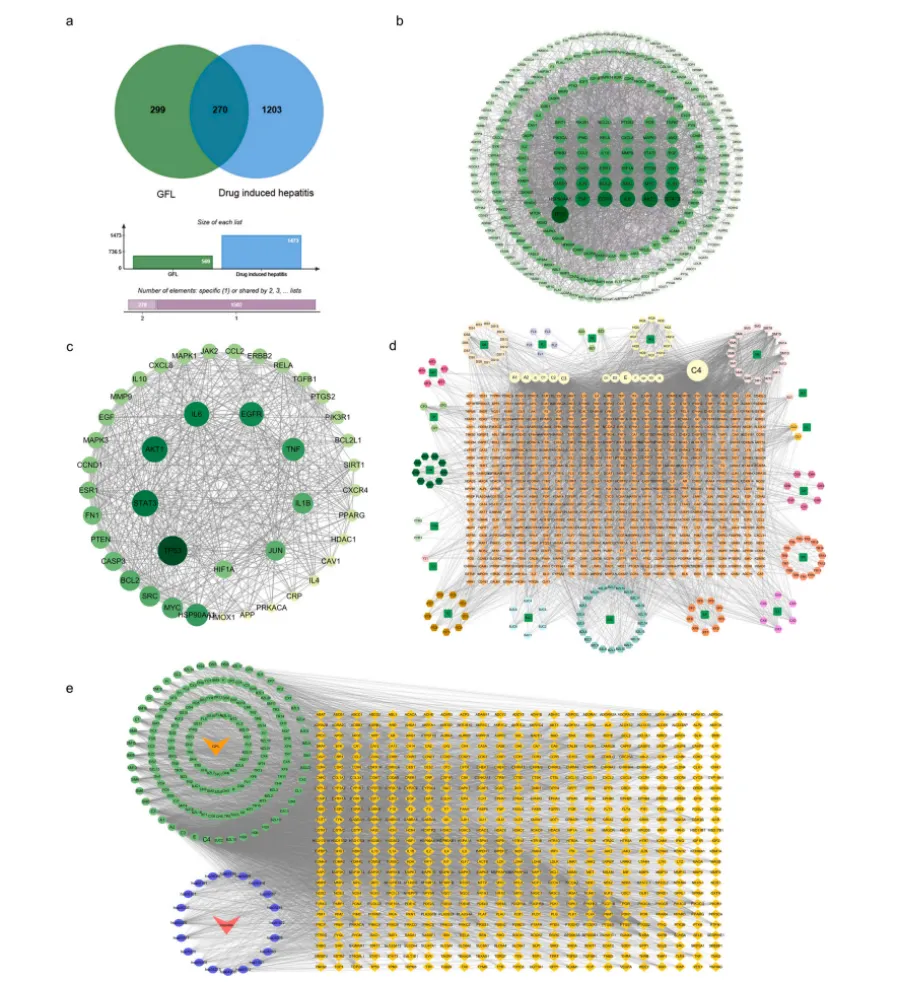
3.6 多组学与分子实验验证GFL保肝作用的核心靶点与活性成分
通过网络药理学分析,本研究筛选得到GFL与药物性肝炎的270个共同作用靶点,其中42个核心靶点主要富集于氧化应激、炎症反应、细胞凋亡、组织纤维化相关通路,同时鉴定出GFL中排名前5的核心活性成分为槲皮素(QCT)、β-谷甾醇、山柰酚、木犀草素、豆甾醇(图3)。
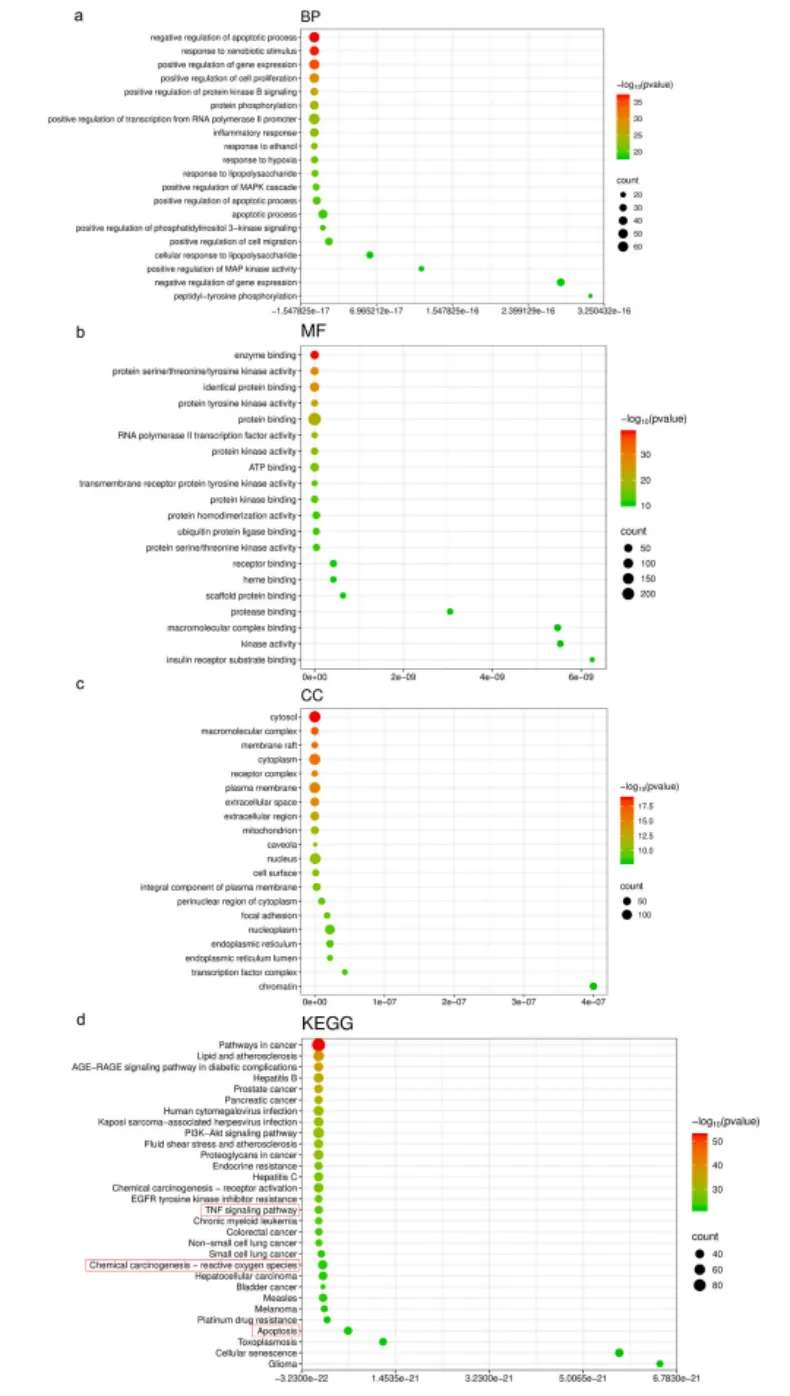
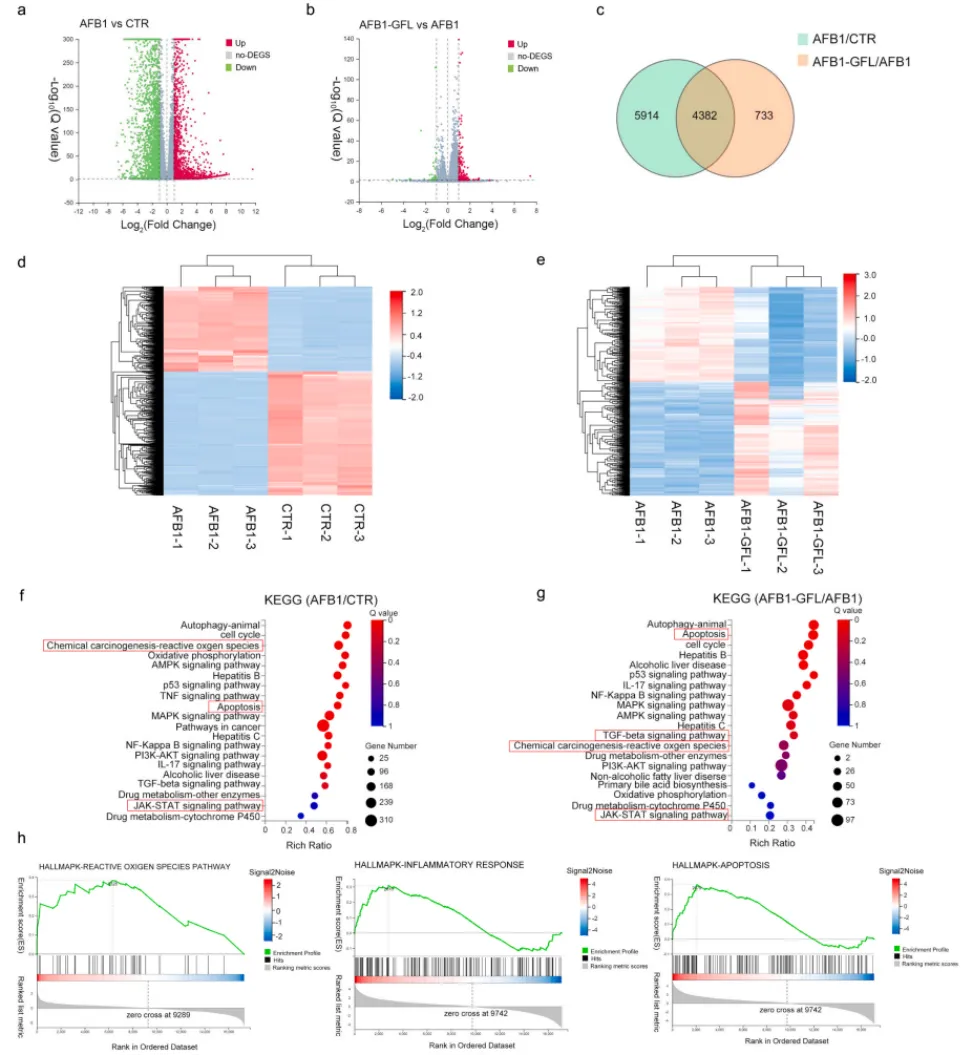
转录组测序结果进一步证实,AFB1处理导致肝细胞上万条基因表达异常,而GFL预处理可显著逆转这些差异基因的表达模式,其调控的核心通路集中于氧化应激、炎症反应与细胞凋亡(图4,图5),与网络药理学预测结果高度一致。
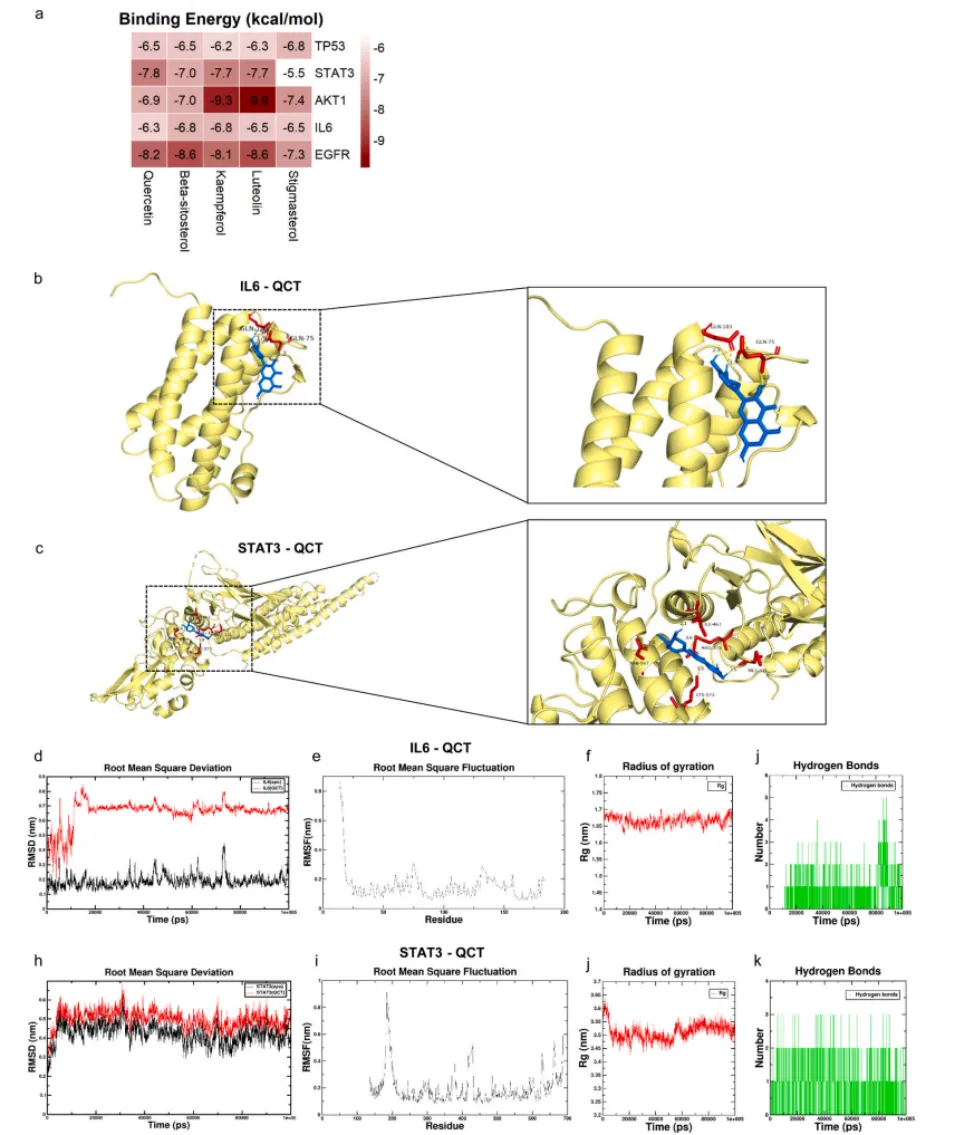
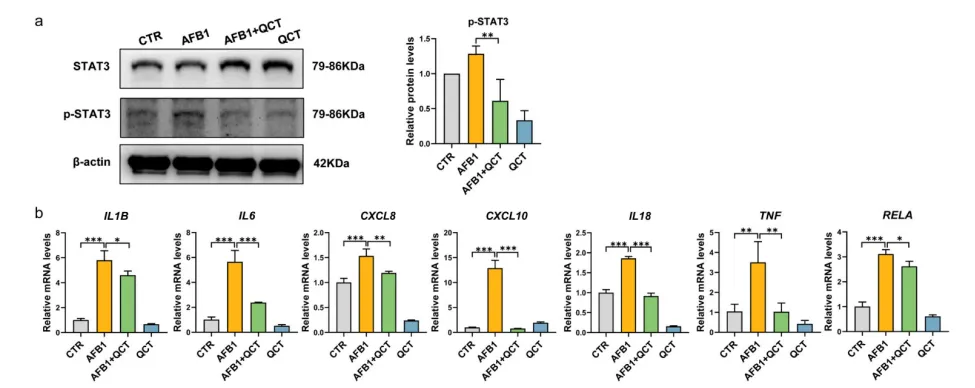
分子对接与100ns分子动力学模拟结果显示,GFL的核心活性成分槲皮素可与IL6、STAT3蛋白形成稳定的氢键结合,结合能分别达-6.3 kcal/mol与-7.8 kcal/mol;体外细胞实验进一步证实,槲皮素可显著抑制AFB1诱导的IL6/STAT3通路激活与下游促炎因子基因的上调,是GFL发挥保肝作用的关键物质基础(图9,图10)。
图3. GFL和AFB1诱导的毒性肝炎共同靶点的网络药理学分析。
图4. GFL和AFB1诱导的毒性肝炎的GO及KEGG通路富集分析。
图5. GFL处理前后AML12细胞的转录组分析。
图9. QCT与IL6和STAT3的分子对接及分子动力学模拟。
图10. QCT抑制AFB1诱导的AML12细胞中IL6/STAT3信号通路的激活。








 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
