NG|北京大学现代农业研究院构建花生T2T基因组,探究着丝粒区间的演化规律,明确了结构变异在花生驯化中的作用.
2026年4月“Nature Genetics”杂志在线发表了北京大学现代农业研究院刘晓芹团队联合多家单位的“Telomere-to-telomere genome assemblies and population resequencing of diploid and allotetraploid peanut varieties”研究论文,该研究论文组装了2个二倍体祖先种和4个代表品种的T2T基因组,并结合521份材料的表型数据,鉴定了调控油含量、种子大小等多个性状的功能基因和优异单倍型,为花生分子设计育种提供了重要的理论依据和基因资源。
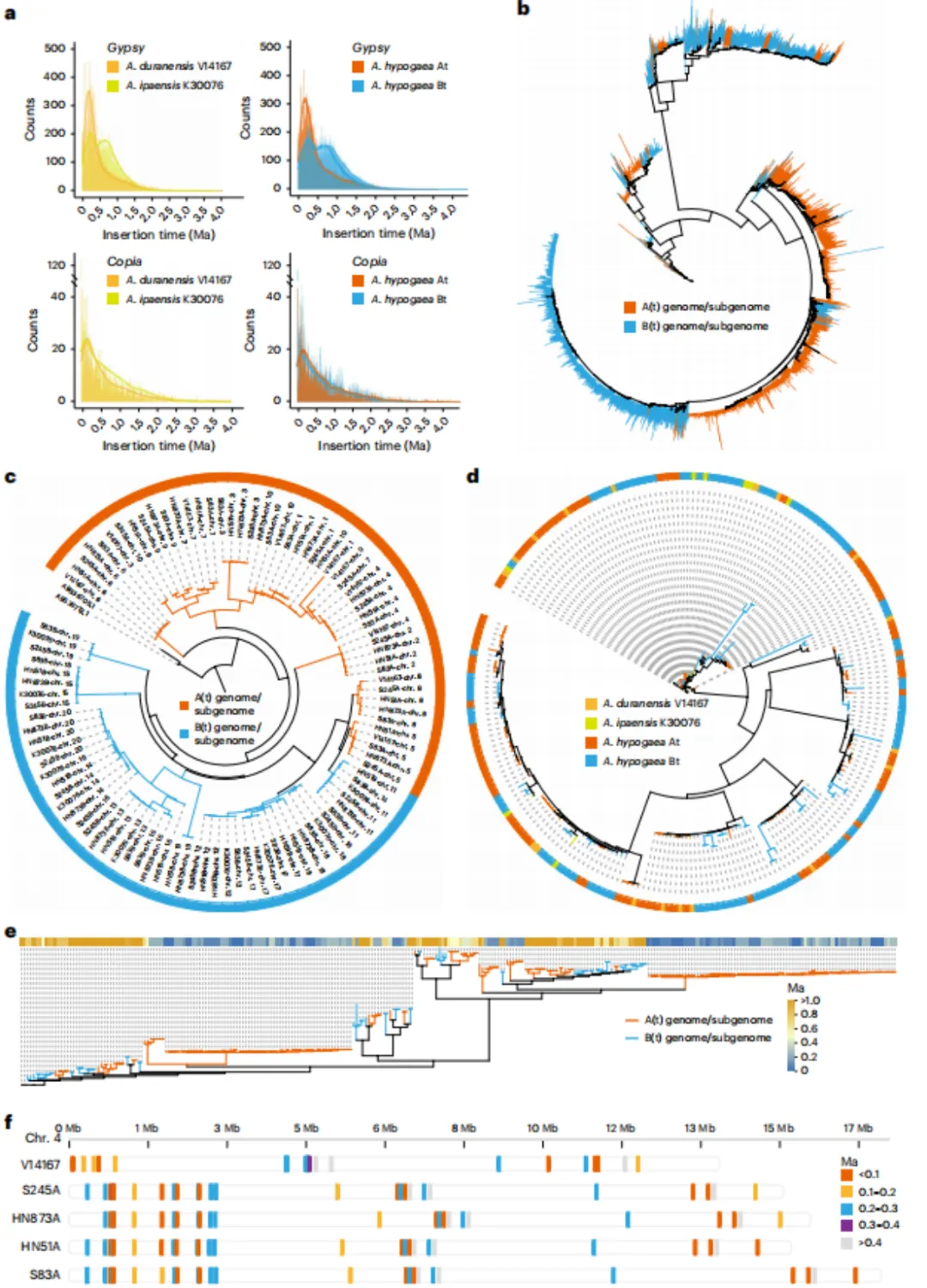
为了解析花生(A. hypogaea)的基因组结构与遗传多样性,研究人员对6个代表性种质(包括2个二倍体祖先和4个异源四倍体栽培种)进行了测序。这些材料表型多样,杂合率为0.14%–0.36%。利用PacBio HiFi(平均深度116×)、Hi-C(112×)和ONT超长读长(70×)测序,结合Hifiasm组装,获得了端粒到端粒(T2T)水平的花生基因组,大小1.18–2.63 Gb,重叠群N50达85.94 Mb。基因组完整性高(BUSCO 98.67%,LAI 21.76,Q47.52),Illumina短读段比对率为99.76%。预测基因数为34,406–75,143,平均功能注释率98.75%,BUSCO注释完整性为98.88%。研究表明,转座元件(TEs)是花生基因组进化的关键驱动力,平均占各基因组的76.27%。其中,B亚基因组(77.77%)的TE含量高于A亚基因组(75.08%),且Gypsy元件占主导地位。LTR插入时间分析显示:A(t)基因组中Gypsy家族扩张约发生于0.20百万年前,而B(t)基因组则出现0.69和0.27百万年前两个扩张峰。不同Gypsy亚型(如Athila、Retand)在亚基因组间表现出差异化的扩张模式与插入时间。在着丝粒区域,Bt亚基因组长度显著减少(平均25.70 Mb),而At亚基因组略有增加(46.11 Mb),且CentO单体在A与B谱系间独立演化。CRM元件在亚基因组间呈穿插分布,Retand元件则在A(t)中有晚期爆发插入,而B(t)中缺失。综上,花生A和B亚基因组在TE动态与着丝粒重塑方面存在明显差异,反映了多倍化后不对称的基因组进化路径。
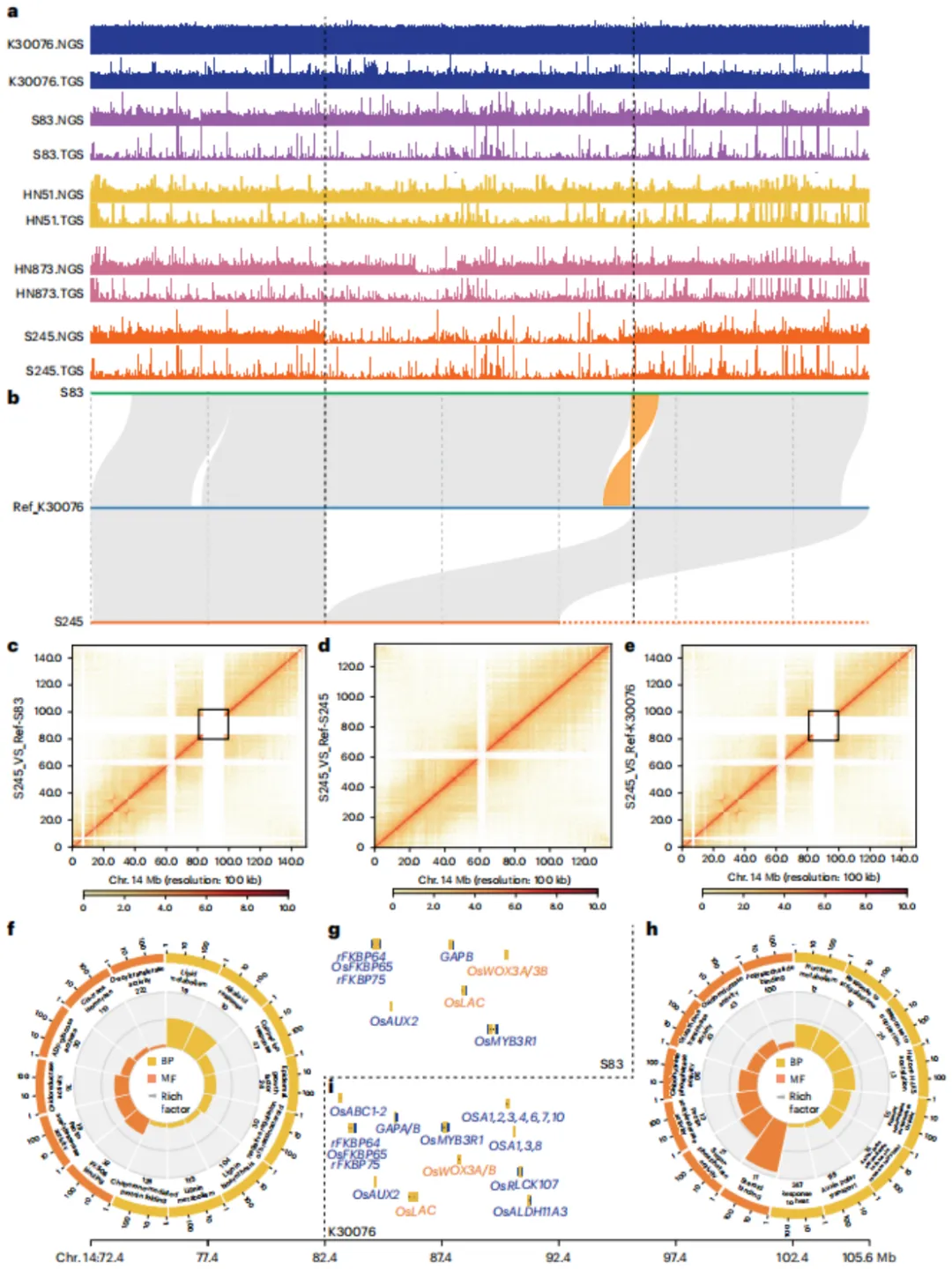
利用T2T基因组与软件工具,在四个四倍体花生栽培种与两个二倍体基因组间共鉴定出大量结构变异(SV),包括插入、缺失、拷贝数变异、易位及倒位。其中,At亚基因组变异数量远多于Bt亚基因组。约45%的SV基因与转座子相关。栽培种间比较显示SV数量存在差异,且SV基因与类黄酮、油菜素内酯及脂肪酸代谢等通路显著相关。进一步发现,A(t)亚基因组中存在保守的染色体重排,而B(t)中特异性重排较少。与地方品种HN873相比,其他品种含有更多特有SV。特别地,在chr.14上发现一个约13.23 Mb的插入片段,存在于所有野生型花生(var. hirsuta)及部分栽培种(var. hypogaea)中,其中包含与植株结构、种子发育及环境响应相关的基因(如OsLAC同源基因、WOX3A/B),可能解释了二者表型差异。
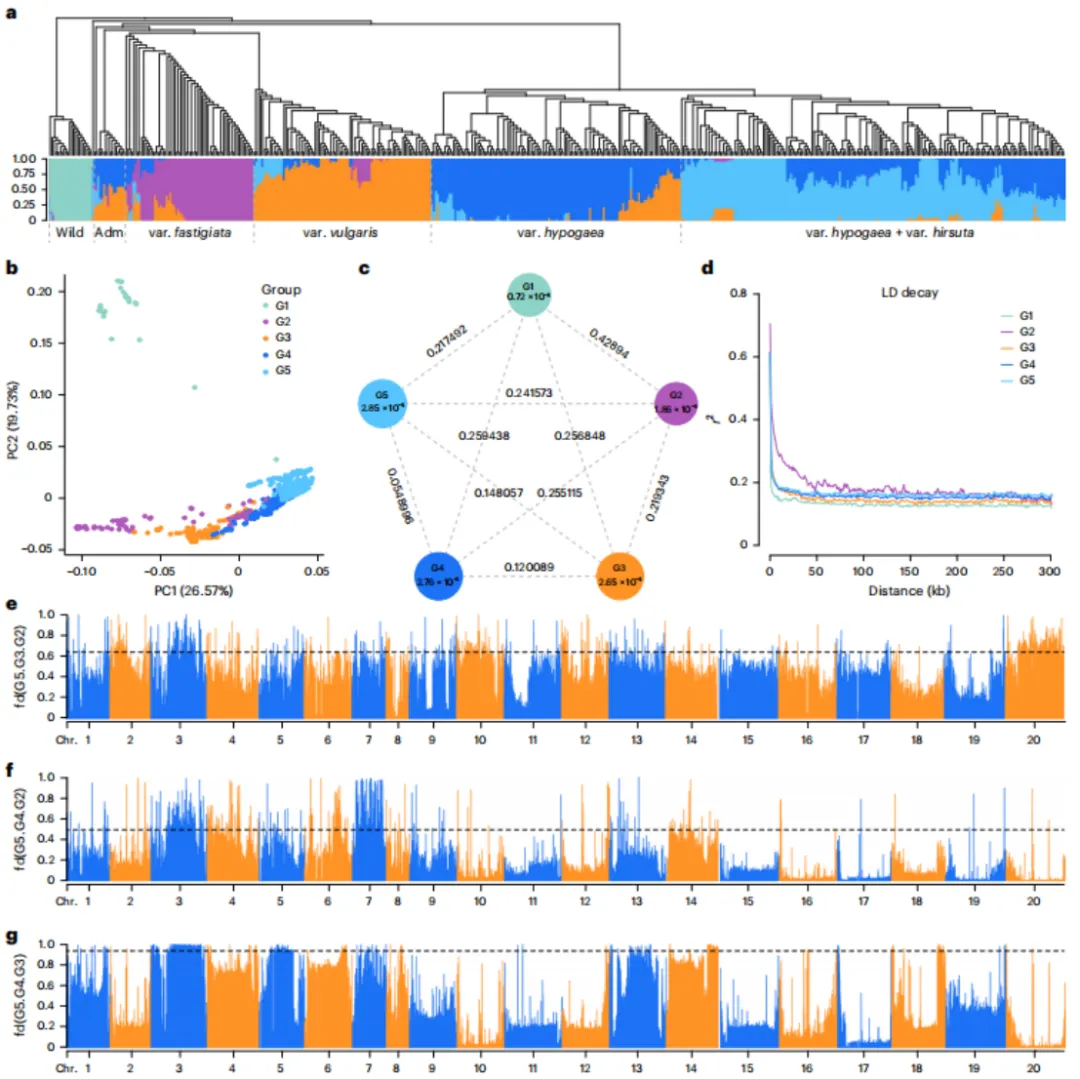
基于521份全球花生种质的重测序数据(平均深度16.76×,覆盖度96.71%),鉴定出约1.01亿个高质量SNP。群体结构分析将这些材料划分为5个遗传类群(G1野生种,G2栽培种变种fastigiata+混合,G3变种vulgaris,G4变种hypogaea,G5变种hypogaea+hirsuta),其中G4与G5遗传差异最小。栽培花生的两个主要类型(fastigiata与hypogaea)约在9400年前分化,后续亚群分化发生在7200–8900年前。选择区域分析显示,B亚基因组受选择区域(平均4.98 Mb)显著大于A亚基因组(2.47 Mb),表明不对称演化。基因渗入主要发生在A亚基因组,尤其G2–G3和G4–G5之间。四倍体中保留了大量来自二倍体的SNP(FST、XP-CLR及渗入区域分别为22.48%、51.97%和49.19%)。GO/KEGG分析鉴定出与脂肪酸合成、类黄酮代谢及营养代谢相关的选择基因。
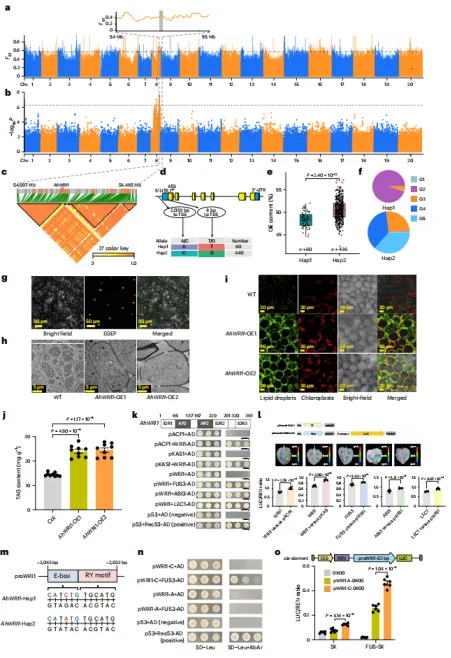
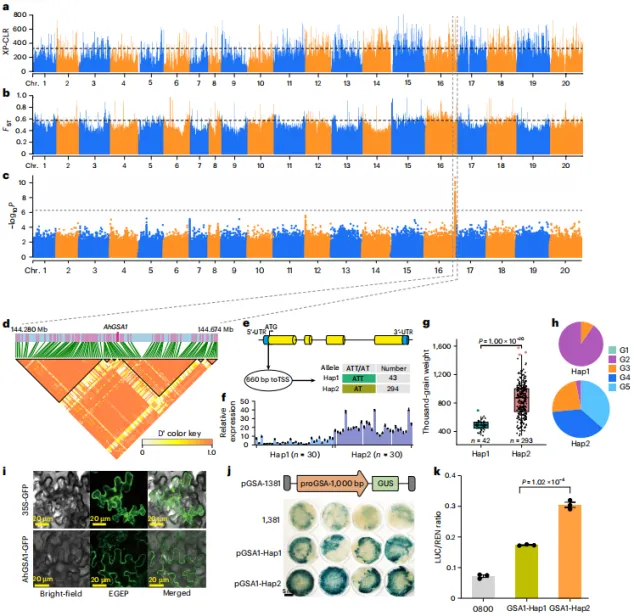
通过GWAS定位到花生第8染色体上一个与含油量相关的200 kb区域,并鉴定出候选基因AhWRI1。该基因启动区存在A→C变异,第一外显子存在非同义SNP(T→G,精氨酸→甲硫氨酸)。AhWRI1在种子发育期优势表达,其中Hap2单倍型(FPKM=24.81)比Hap1型(FPKM=16.30)具有更高的表达量和含油量(54.10% vs. 48.41%)。亚细胞定位证实其为转录因子。在油菜和大豆中过表达AhWRI1可促进脂滴积累和脂肪酸含量升高。进一步发现,AhLEC1、AhFUS3、AhABI3等种子发育关键调控因子结合于AhWRI1启动子,其中启动子区的A→C突变位于E-box/RY基序内,该突变破坏与AhFUS3的结合,但显著增强启动子活性,从而导致AhWRI1表达差异和不同品种间含油量的变异。同时克隆了含ATT或AT突变的1000 bp启动子序列,并证实其能驱动GUS报告基因表达,其中AT启动子的活性显著强于ATT。亚细胞定位显示AhGSA1蛋白定位于细胞质膜。双荧光素酶实验进一步确认,AT启动子活性极显著高于ATT(P = 1.02×10⁻⁴)。这些结果强调了启动子区域插入/缺失变异对AhGSA1转录调控的重要性。
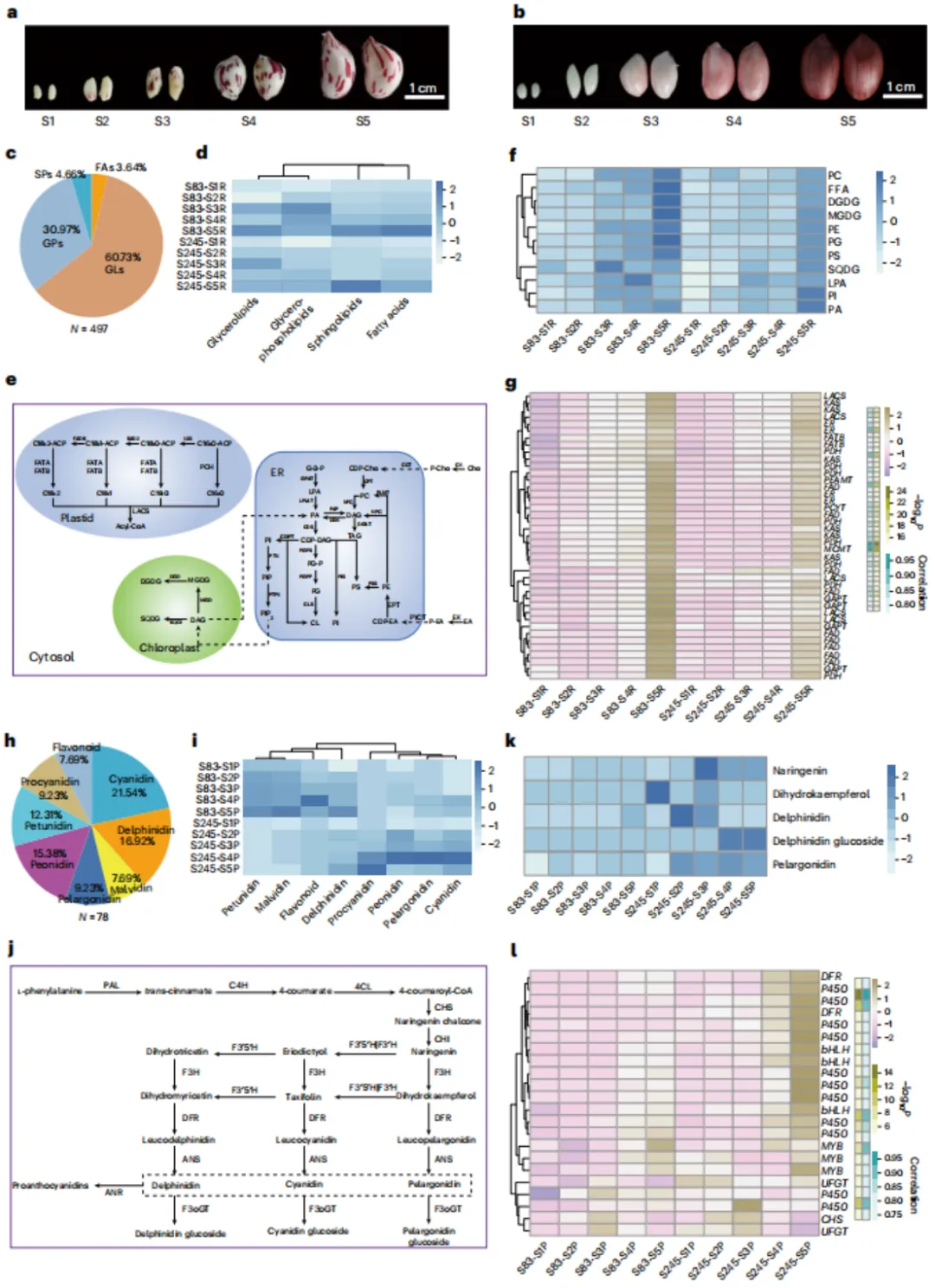
通过对两个花生品种(S83和S245)在五个发育阶段的转录组与代谢组联合分析,系统揭示了种子发育过程中脂质和花青素代谢特征。共鉴定出497种脂质代谢物,其中S83在发育后期脂质含量显著高于S245。通过WGCNA分析,筛选出与脂质代谢高度相关的基因模块,关键基因包括脂肪酸去饱和酶和酮脂酰-ACP合酶。同时鉴定出78种花青素代谢物,S83富含矮牵牛素等,S245则富含原花青素等,差异可能影响种皮颜色。进一步分析发现,MYB和bHLH转录因子表达与花青素积累趋势一致。本研究识别出调控花生油脂含量和种皮颜色的关键基因,为高油品种选育及种子颜色改良提供了重要候选基因与理论依据。
该文章创新点
(1)构建了两个二倍体与四个四倍体花生品种的T2T完整基因组组装。
(2)通过群体遗传揭示了育种过程中亚基因组间的不对称选择事件,并挖掘了重要性状关键基因。
(3)整合转录组与代谢组分析,明确了种子发育过程中的代谢通路。
https://doi.org/10.1038/s41588-026-02577-z



 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
