(点击页面左下角阅读原文,直达文献页面)
一、中文标题
降解者与非降解者间的互利共生稳定了天然生物聚合物降解群落的功能(Mutualism between degraders and nondegraders stabilizes the function of a natural biopolymer- degrading community)
二、发表单位及通讯作者
三、科学问题
本研究旨在探究天然木质纤维素降解微生物群落(EMSD5)中,非降解微生物(如大肠杆菌)对降解者(如梭菌)的具体贡献,以及群落内部如何调控降解产物的分配以限制非降解者的过度利用(“公地悲剧”),从而维持群落结构与功能的稳定。
四、发表时间
五、摘要
本研究以高效转化玉米芯为异丙醇的木质纤维素降解群落EMSD5为模型,揭示了降解者与非降解者间稳定的互利共生机制。研究发现,非降解者(如大肠杆菌)在群落初期占主导,通过快速消耗氧气创造厌氧环境,并提供关键生长因子生物素,从而扩展了严格厌氧降解者(如梭菌)的生态位。在扩展的生态位内,降解者通过两种互锁机制限制非降解者对降解产物的利用:1)资源分配:拉氏梭菌产生的低聚糖大部分不能被大肠杆菌直接利用,其中一部分可被丁酸梭菌进一步降解为单糖;2)酶分泌反馈调控:丁酸梭菌的糖苷酶分泌受胞外木糖浓度调节,共培养时分泌减少。基于此,研究构建了由表达异源木聚糖酶的大肠杆菌与丁酸梭菌组成的合成群落,成功实现了无需厌氧预处理的半纤维素至异丙醇的转化。本研究阐明了自然降解群落中稳定合作维持的生态机制,并为设计高效生物质转化合成群落提供了新策略。
六、研究背景
异养微生物群落的生物聚合物降解是全球碳生物地球化学循环的关键驱动力。在这些群落中,生物聚合物的降解能力通常分布在多个菌株之间,形成了复杂的代谢相互依赖关系,这种功能分布通过协同作用增强了群落的恢复力和效率。因此,维持不同物种间稳定的相互作用对于生物聚合物降解至关重要。
生物聚合物降解通常始于专性降解者分泌胞外酶,将聚合物裂解为可运输的降解产物。这些产物作为“公共物品”,可被降解者和“非降解者”(即自身不产生降解酶的菌株)共同利用。这种动态容易引发“公地悲剧”,即非降解者过度利用公共物品而不贡献于集体降解努力,从而破坏群落稳定和生态系统功能。尽管存在这种挑战,合作性相互作用在生物聚合物降解群落中仍普遍存在,并对塑造微生物群落结构和进化轨迹至关重要。因此,理解这些相互作用如何被调控、以及群落如何缓解剥削行为,对于阐明自然群落组装和设计高效降解联合体至关重要。
现有研究多利用合成微生物群落探讨微生物如何管理在交换氨基酸、维生素等共有价值营养物质时的剥削行为。然而,生物聚合物降解群落内部的交叉喂养动态具有独特特征,例如,降解者分泌的胞外酶不能被其他微生物直接消耗,并在环境中持续释放降解产物,加剧了非降解者对降解者的剥削。特别是在高扩散速率的环境(如水生系统)中,绝大部分降解产物在吸收前就已扩散远离降解者。这表明,降解者必须采用严格的调控机制来控制酶分泌并防止被过度利用,但这些具体机制尚不明确。此外,尽管降解者塑造非降解者的组装,非降解者也通过交叉喂养等方式影响降解者的生长和行为,但非降解者是否还通过其他机制影响降解者的生态位,以及这些相互作用如何影响群落的整体效率和稳定性,仍知之甚少。
为应对这些挑战,本研究聚焦于地球上最丰富的生物聚合物——木质纤维素的降解,并以从堆肥中富集、能稳定高效转化未处理木质纤维素为异丙醇的天然群落EMSD5为研究对象,旨在揭示其内部降解者与非降解者间维持稳定合作的生态与分子机制。
七、研究结果
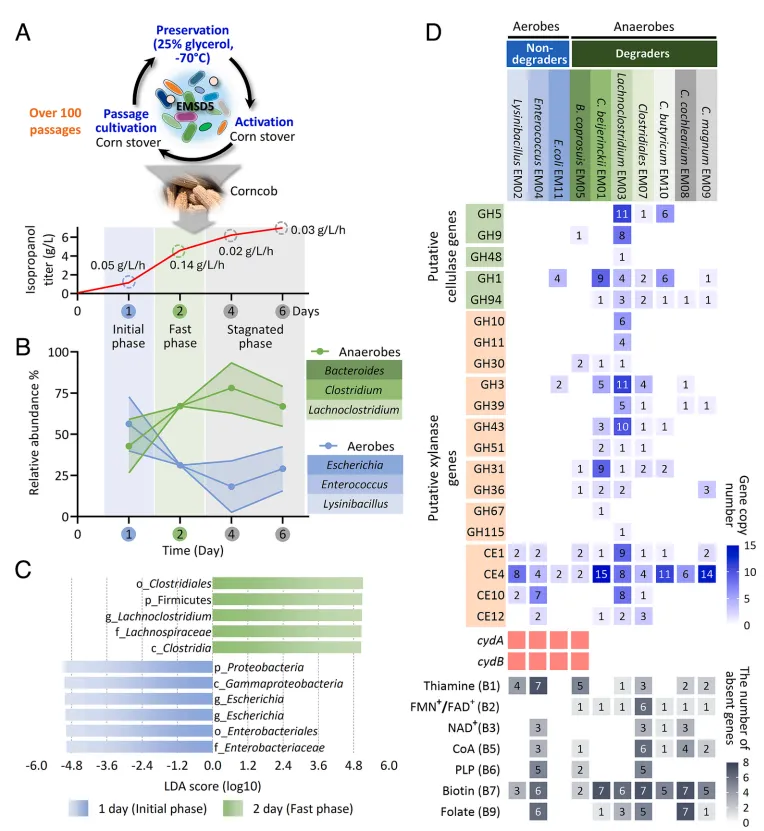
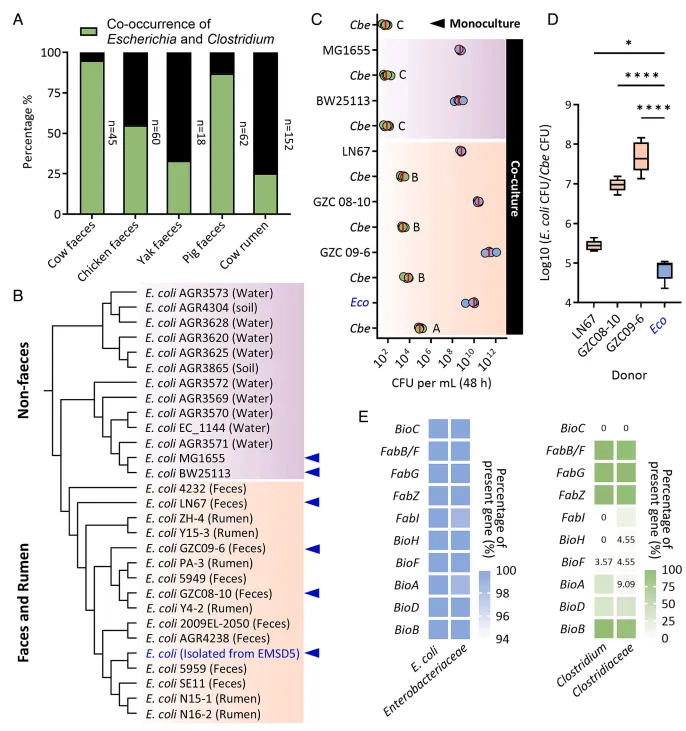
结果1、好氧非降解者在木质纤维素生物转化初期占据主导 通过分析EMSD5在6天异丙醇生产过程中的群落动态,发现其可分为初始期、快速期和停滞期。好氧菌(如大肠杆菌属、赖氨酸芽孢杆菌属、肠球菌属)在初始期(第1天)占据主导,其中大肠杆菌相对丰度高达39.2%。随后,厌氧降解者(如拉氏梭菌属、梭菌属、拟杆菌属)的相对丰度从第1天的49.9%上升至第4天的81.4%。宏基因组分析表明,负责纤维素和半纤维素降解的糖苷水解酶基因主要由厌氧菌(尤其是拉氏梭菌和梭菌)编码,而仅丁酸梭菌EM01拥有完整的异丙醇生产途径,确认好氧菌主要作为非降解者发挥作用。
图1 菌群分析
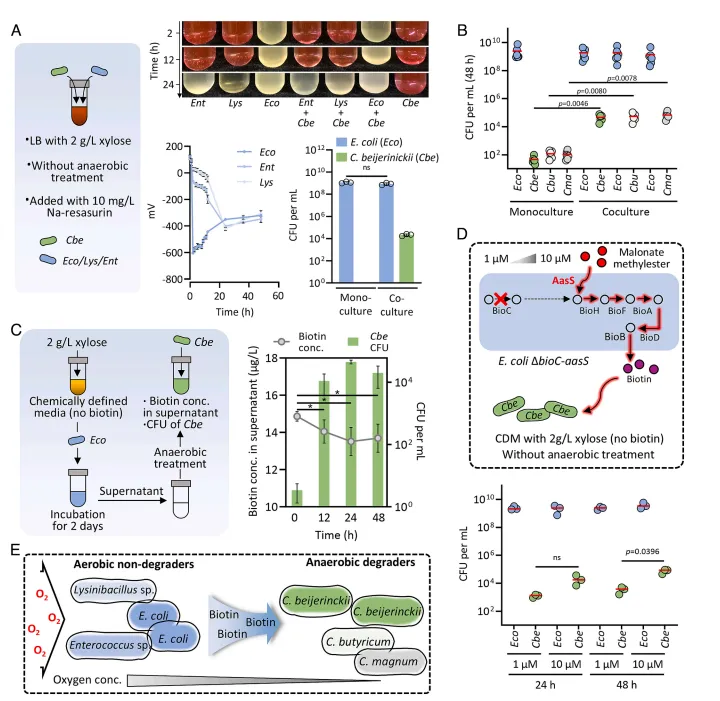
结果2、好氧非降解者通过提供厌氧条件和生物素扩展梭菌的生态位 鉴于梭菌的严格厌氧性且培养基未经厌氧处理,研究假设部分EMSD5成员通过创造厌氧生态位来促进梭菌生长。生物信息学分析证实好氧菌(如大肠杆菌EM11)拥有耗氧关键基因。实验表明,大肠杆菌与丁酸梭菌共培养可在2小时内快速耗尽氧气(刃天青褪色,氧化还原电位降至-300 mV以下),使丁酸梭菌成功生长;而其他好氧菌耗氧较慢。此外,梭菌缺乏完整的生物素合成途径,而大肠杆菌EM11则拥有完整的维生素合成通路。生长实验证实生物素是梭菌生长的限制性营养因子。在缺乏生物素的限定培养基中,与大肠杆菌共培养可使梭菌的菌落形成单位增加100倍。利用生物素合成缺陷(ΔbioC)并引入异源酶以利用丙二酸甲酯的大肠杆菌突变体进行共培养,进一步证实了大肠杆菌通过提供生物素支持梭菌生长。
图2 生态位分析
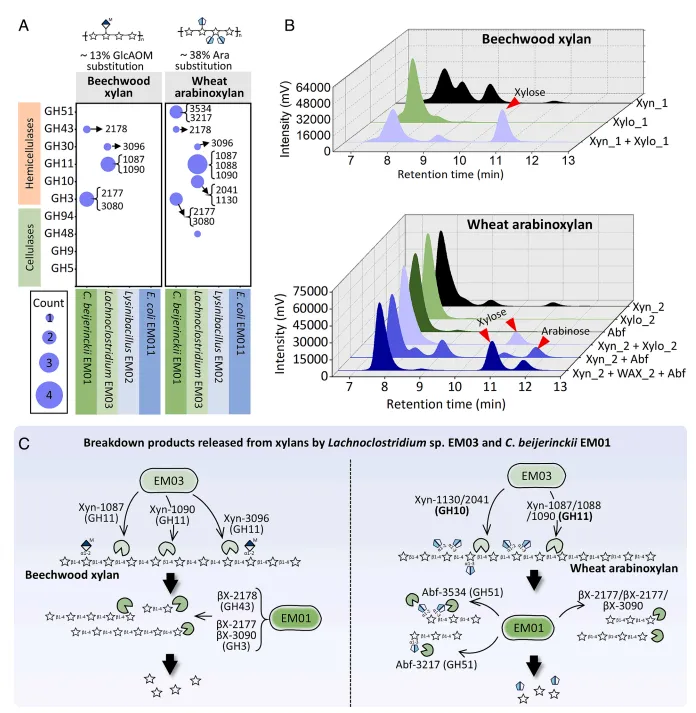
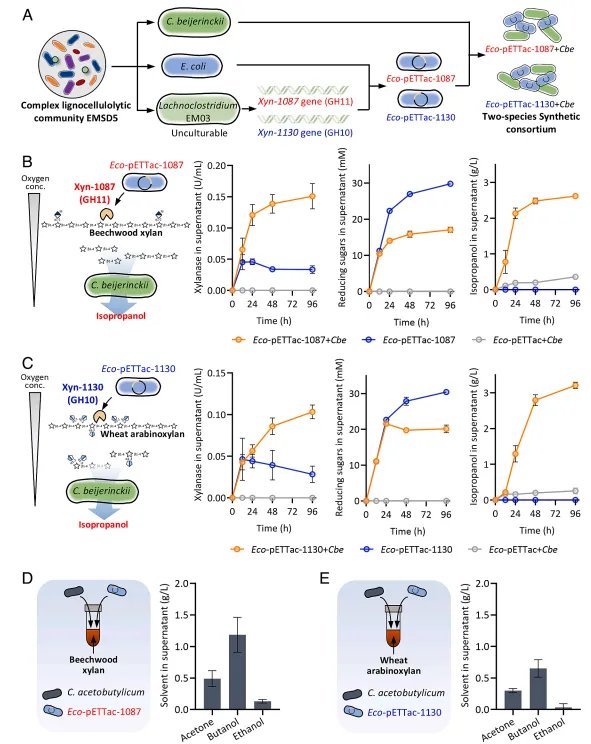
结果3、降解者(拉氏梭菌与丁酸梭菌)通过互利互作释放半纤维素降解产物 胞外蛋白质组学分析鉴定出拉氏梭菌EM03和丁酸梭菌EM01是EMSD5降解玉米芯及其组分(如山毛榉木聚糖、小麦阿拉伯木聚糖)的主要胞外糖苷水解酶生产者。针对不同木聚糖,两者分泌互补的酶系:EM03主要分泌木聚糖酶(GH10, GH11)将木聚糖降解为低聚糖;EM01则分泌β-木糖苷酶(GH3, GH43)和α-阿拉伯呋喃糖苷酶(GH51)将低聚糖进一步水解为单糖(木糖、阿拉伯糖)。体外酶组合实验证实,两者酶系协同作用能显著提高还原糖及单糖的产量,揭示了在木聚糖降解中存在的互利互作链。
图3 菌株交互
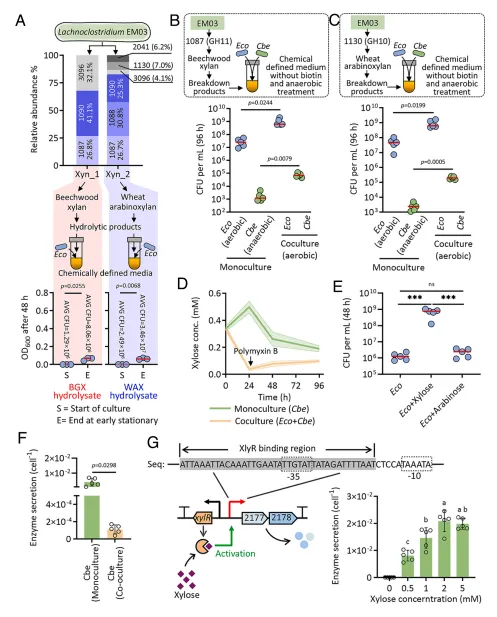
结果4、降解产物可利用性受资源分配和酶反馈调控的双重限制 尽管EM03产生大量低聚糖,但研究发现大肠杆菌无法直接利用木聚糖底物,且对其降解产生的低聚糖利用有限。然而,当大肠杆菌与丁酸梭菌共培养时,其生长得到促进。进一步研究发现,只有由EM03特定木聚糖酶(如GH11 1087, GH10 1130)产生的低聚糖能被丁酸梭菌有效利用并转化为木糖,后者随后被大肠杆菌消耗。这表明存在资源分配:EM03产生的低聚糖种类多样,但仅特定种类能被下游的丁酸梭菌利用。
此外,研究发现了酶分泌的反馈调控机制。虽然丁酸梭菌与大肠杆菌共培养时总β-木糖苷酶活性更高,但单位细胞的酶分泌量在共培养条件下更低。遗传分析发现,丁酸梭菌的β-木糖苷酶基因上游存在木糖阻遏蛋白XylR。当胞外木糖浓度高时,XylR从启动子解离,促进酶基因转录和分泌;当木糖被快速消耗(如被共培养的大肠杆菌利用)导致浓度降低时,酶分泌受到抑制。这种调控机制减少了在资源有限条件下不必要的酶合成,从而限制了单糖对快速生长的非降解者(大肠杆菌)的可利用性。
图4 资源分配
结果5、大肠杆菌对梭菌的互利作用在木质纤维素分解环境中普遍存在 对337个微生物群落的调查显示,大肠杆菌与梭菌在多种动物粪便及瘤胃等木质纤维素分解环境中频繁共存(共存率25.2%-95.6%)。系统发育分析表明,来自粪便环境的菌株(包括EMSD5中的大肠杆菌)比实验室模式菌株(如MG1655)更能有效促进丁酸梭菌生长,且EMSD5来源的菌株效率最高。基因组分析表明,肠杆菌科普遍拥有完整的生物素合成通路,而梭菌科则普遍存在基因缺失,提示两者间可能存在广泛的生物素交叉喂养。
图5 菌群进一步分析
结果6、利用梭菌与大肠杆菌的互利共生设计半纤维素生物转化合成群落 基于上述机理,研究构建了合成群落:将来自未培养拉氏梭菌的关键交叉喂养木聚糖酶基因(xyn-1087和xyn-1130)在大肠杆菌中异源表达,然后与丁酸梭菌共培养。该合成群落成功地将山毛榉木聚糖和小麦阿拉伯木聚糖分别转化为2.62 g/L和3.21 g/L的异丙醇,且无需厌氧预处理。优化接种比例表明该互利共生体系具有鲁棒性。该框架还可拓展至其他产溶剂梭菌(如丙酮丁醇梭菌),用于生产丙酮-丁醇-乙醇,证明了其广泛适用性。
图6 合成群落生产化学品
八、讨论
本研究揭示了天然木质纤维素降解群落EMSD5中,降解者与非降解者间多层次互利共生维持群落稳定的生态机制。创新性地发现,非降解者不仅是被动的受益者,更是生态位的积极开拓者,通过创造厌氧条件和提供必需维生素(生物素)为降解者铺平道路。同时,降解者通过酶底物特异性导致的资源分配和响应底物浓度的酶分泌反馈调控这两大互锁机制,有效限制了公共产物(单糖)向非降解者的流动,从而缓解了“公地悲剧”。这些发现为理解复杂自然微生物群落的稳定组装与功能维持提供了实证。基于此生态原理构建的合成群落,成功实现了半纤维素到化学品的高效转化,证明了将生态学见解应用于合成生物学设计的可行性,为开发更稳定、高效的生物质转化系统提供了新范式。