Micro Res | 云南农业大学:跨界合成菌群通过调控根际微生物组与植物防御控制柑橘黄龙病及根腐病
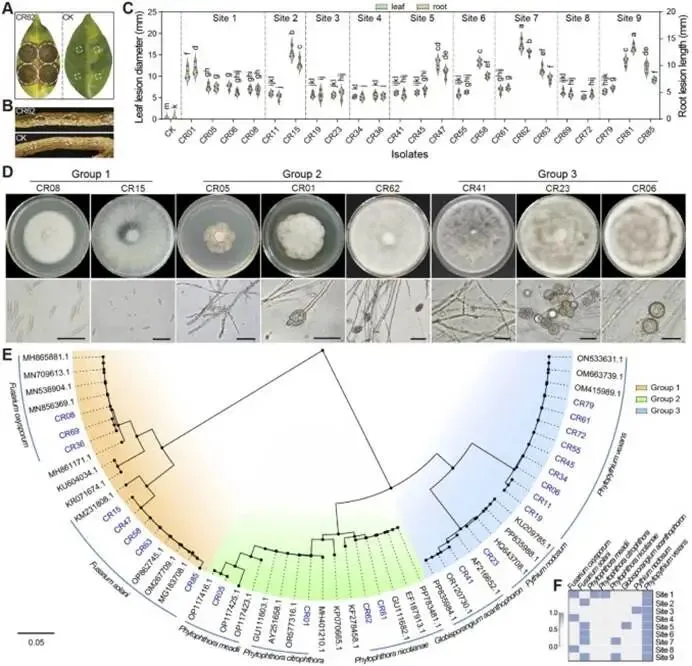
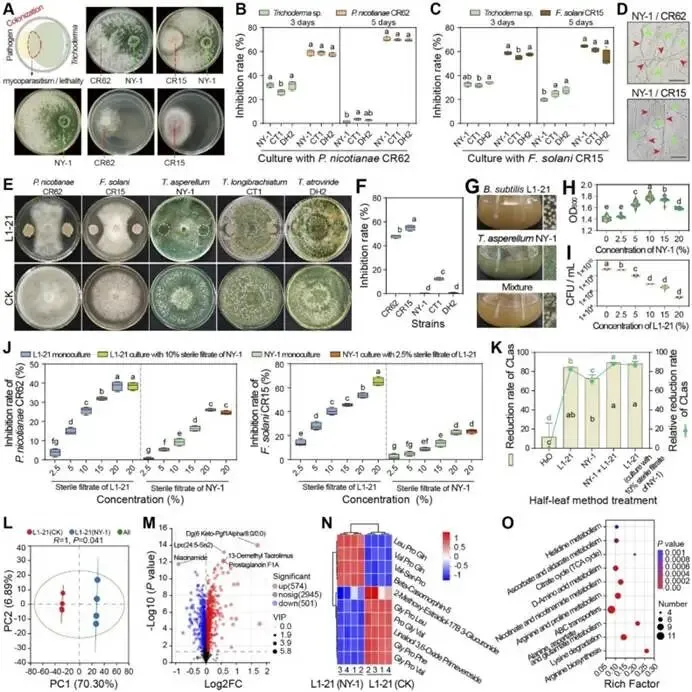
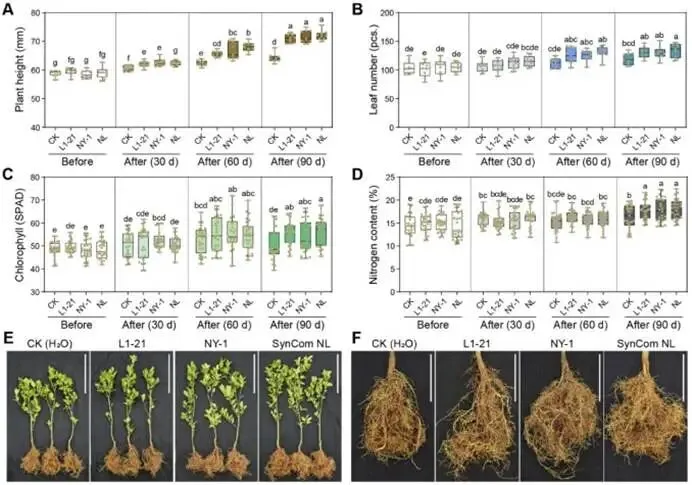
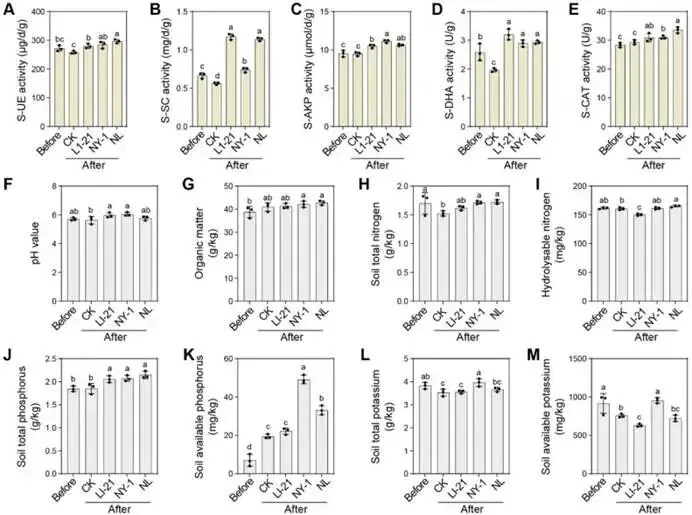
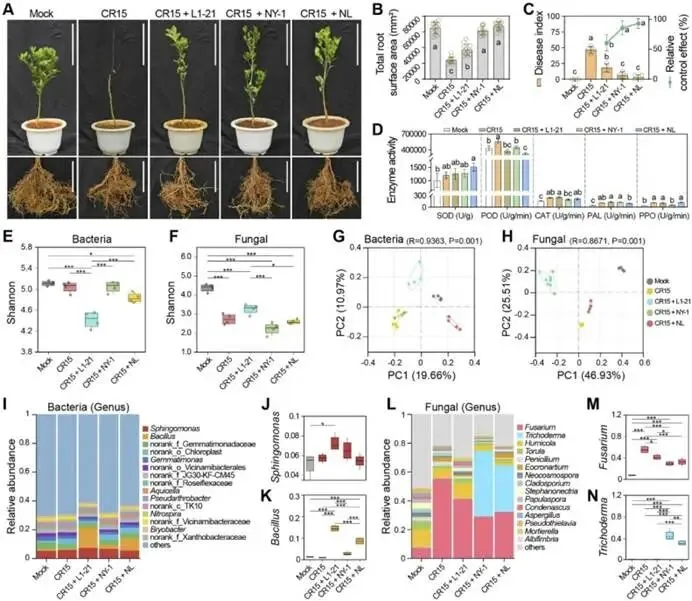
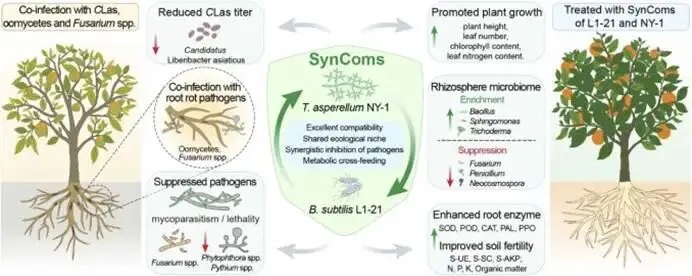
由柑橘黄龙病菌(CLas)引起的柑橘黄龙病(HLB)与根腐病的协同发生严重威胁全球柑橘产业。本研究对中国7个省份的9个主要柑橘产区开展田间调查,筛选出高致病性病原菌烟草疫霉 CR62(Phytophthora nicotianae)和茄病镰刀菌 CR15(Fusarium solani )作为靶标,用于筛选具有寄生活性的木霉菌株。通过与防治黄龙病的枯草芽孢杆菌 L1-21(Bacillus subtilis)的代谢互作兼容性分析,成功构建跨界合成菌群(SynCom)。盆栽实验评估了促生与根腐病防控效果,高通量测序(16S rRNA/ITS)解析了根际微生物组的调控机制。结果表明:黄龙病与根腐病在田间广泛共存,其中卵菌纲(疫霉菌 Phytophthora spp. 和腐霉菌 Pythium spp.)占根腐病病原体的65.22%。烟草疫霉与茄病镰刀菌致病力最强,但根腐病病情指数与CLas菌量无显著相关性。定向筛选显示,棘孢木霉 NY-1(Trichoderma asperellum)对根腐病病原菌抑制率超过60%,且与枯草芽孢杆菌L1-21高度兼容。此外,NY-1的无细胞滤液通过调控氨基酸代谢、能量代谢及ABC转运蛋白通路,显著重塑L1-21的整体代谢谱,全面改变其细胞代谢活性。关键的是,SynCom NL处理90天后显著促进植株生长,并提高根际脲酶和蔗糖酶活性。该群落富集了有益微生物(芽孢杆菌属、鞘氨醇单胞菌属、木霉属、腐质霉属),并抑制病原菌(镰刀菌属、青霉属、新赤壳属),同时激活根系防御酶(超氧化物歧化酶、过氧化氢酶、苯丙氨酸解氨酶、多酚氧化酶),增强根系保护能力,实现92.86 ± 8.25%的根腐病防控效果。本研究针对黄龙病-根腐病复合病害开发的木霉-芽孢杆菌跨界合成菌群,为生物综合防控与柑橘可持续生产提供了技术基础。篇名: Cross-kingdom synthetic microbial consortium controls citrus Huanglongbing and root rot by modulating the rhizosphere microbiome and plant defense期刊: Microbiological ResearchDOI: 10.1016/j.micres.2026.128528本研究在中国主要柑橘产区(云南、四川、广西、广东、江西、湖南、福建)开展田间调查与样本采集。采用分层随机抽样选取具有典型黄龙病症状的果园,利用五点取样法收集根系与叶片样本,并依据0-5级标准评估根部腐烂严重度及计算病情指数。通过TaqMan qPCR检测样品中柑橘黄龙病菌(CLas)的携带情况。采用组织分离法从病根中分离根腐病原菌,结合形态学观察与ITS序列系统发育分析进行鉴定,并通过柯赫氏法则验证致病性。后续开展木霉菌筛选、与枯草芽孢杆菌L1-21构建跨界合成菌群(SynCom),利用盆栽试验评估其促生与防病效果。此外,运用UHPLC-MS/MS分析差异代谢物,并基于高通量测序解析根际微生物群落结构。田间调查显示,健康柑橘树表现为枝叶繁茂、结果正常,而染病植株则长势衰弱、叶片稀疏并伴随萎蔫,最终导致植株死亡(图1B)。黄龙病(HLB)侵染的叶片呈现典型黄化症状,与健康叶片的深绿色形成鲜明对比(图1C)。健康根系具有发达的须根且结构完整,而根腐病则导致表皮腐烂及须根大量脱落。根腐症状主要出现在HLB感染植株中,表明其根系结构遭受严重破坏(图1D)。对CLas菌量(Ct值)及根腐病病情指数(DI)的定量检测表明,所有田间采样点的叶片和根系Ct值均低于35,证实存在HLB感染。根系Ct值普遍低于叶片,表明根组织中CLas菌量更高。根腐病DI与叶片或根系的CLas菌量均无显著相关性(P > 0.05)。例如,四川木里县(Site 4)的根腐病DI显著高于其他地区,但其HLB严重程度相对较轻,暗示柑橘黄龙病与根腐病的发生存在区域异质性。从9个主要柑橘产区的腐烂根系中分离获得86株纯培养物,其中53株(61.63%)来源于PDA培养基,33株(38.37%)来源于卵菌选择性培养基。通过离体叶片及根系接种试验,23株(26.74%)表现出明显致病力,经科赫法则再分离验证确认为病原菌。接种叶片出现扩展性病斑并伴随组织腐烂,接种根系则表现褐变、腐烂及表皮脱落,而对照组未出现症状(图2A-B)。其中,CR62在叶片上引起的病斑面积最大,其次为CR15和CR81;而CR81在根系接种中致病力最强,病斑长度达13.28 ± 0.62 mm(图2C)。图2. 柑橘根腐病病原菌的致病力测定、形态特征与分子鉴定。A-B:CR62菌株接种柑橘叶片(A)和根系(B)后的病斑表型(虚线框标示病斑区域);C:各菌株对叶片和根系的致病力差异;D:病原菌分组(1、2、3组)在PDA上的菌落形态(7 dpi)及菌丝与孢子特征(比例尺=20 μm);E:基于ITS序列的病原菌系统发育树(蓝色标注本研究分离株;标尺0.05代表遗传距离)。不同菌株的菌落形态、生长速率及菌丝与孢子特征差异显著。根据形态学特征,23株致病菌初步分为3组8种:第1组(镰刀菌属 Fusarium spp.;8株,34.78%) :菌落蓬松或放射状生长(直径6-8 cm),产生镰刀状分生孢子(20-35 × 3-5 μm,3-5隔膜);第2组(疫霉属 Phytophthora spp.;4株,17.39%) :菌落白色、毡状或棉絮状(直径4-6 cm),菌丝无隔,孢子囊球形或卵圆形(30-50 × 20-35 μm),部分菌株无孢子囊但形成囊状菌丝;第3组(腐霉属 Pythium spp.;11株,47.83%) :菌落白色且生长迅速(直径7-9 cm),菌丝无隔,产生游动孢子或卵圆形孢子囊(直径15-25 μm)(图2D)。ITS序列BLAST比对与系统发育分析表明:第1组菌株与尖孢镰刀菌(Fusarium oxysporum)和茄病镰刀菌(F. solani)的序列相似度为99.42-100.00%;第2组与烟草疫霉(Phytophthora nicotianae)、柑橘褐腐疫霉(P. citrophthora)及蜜色疫霉(P. meadii)相似度为100.00%;第3组与刺器腐霉(Globisporangium acanthophoron,异名 Pythium acanthophoron)、P. nodosum及钟器腐霉(Phytopythium vexans)相似度>96.97%(表S3)。系统发育树中,各组菌株均与对应参考物种聚类,与形态学分类一致(图2E)。所有鉴定病原菌中,恶疫霉(P. vexans)分离频率最高(9株),其次为茄病镰刀菌(5株)(图2F)。结合致病力与物种鉴定结果,调查区域的柑橘根腐病主要由卵菌和镰刀菌引起,其中烟草疫霉(CR62、CR81)、柑橘褐腐疫霉(CR01)及茄病镰刀菌(CR15、CR47、CR58、CR63、CR85)致病力最强。(3)木霉与枯草芽孢杆菌L1-21跨界共生体的构建木霉菌株对烟草疫霉CR62表现出强拮抗活性,所有菌株抑制率均高于60.59 ± 1.70%(图S1A)。其中17株对CR62具有寄生作用,导致PDA平板上菌落融合;15株表现出致死效应,被寄生的CR62在卵菌选择性培养基上无法再生(图S1B)。针对茄病镰刀菌CR15,木霉菌株抑制率超过50.00 ± 0.97%(图S1A),6株显示寄生互作但观察期内未致死(图S1C)。与L1-21共培养时,所有木霉菌株均受中度抑制(抑制率<42.67 ± 0.49%),其中L1-21对棘孢木霉NY-1的抑制率最低(3.66 ± 1.22%)。共培养7天后,9株木霉与L1-21形成融合菌落并占据相同生态位。划线实验证实L1-21被寄生后在LB培养基上仍存活,表明亲和性良好(图S1D)。基于对病原菌的寄生性、致死力及与L1-21的亲和性,棘孢木霉NY-1、长枝木霉CT1和深绿木霉DH2综合表现最优,且与CR62和CR15共培养时表现出强空间竞争(图3A)。NY-1、CT1和DH2对CR62和CR15的抑制与寄生作用显著,而病原菌对木霉的抑制效应随时间变化差异明显。第5天时,木霉的持续寄生导致CR62和CR15生长停滞,其对木霉的抑制率极低(CR62和CR15对NY-1的抑制率分别为1.45 ± 1.17%和19.90 ± 1.21%)(图3B-C)。NY-1对两种病原菌的寄生作用最显著,其菌丝(绿色箭头)缠绕CR62和CR15菌丝(红色箭头)(图3D)。L1-21对CR62和CR15形成清晰抑菌圈,抑制率分别为47.92 ± 1.38%和55.16 ± 2.30%。与木霉共培养时,L1-21与各木霉菌落融合,其对NY-1(0.50 ± 0.40%)和DH2(0.66 ± 0.55%)的抑制率低于1%(图3E-F),表明在此特定条件下,二者无直接拮抗或组成型分泌扩散性抑制剂。图3. 木霉对病原菌的拮抗作用及与枯草芽孢杆菌L1-21合成群落的构建。A:木霉与病原菌共培养示意图(红色虚线框示寄生与致死观察区)及NY-1与CR62、CR15共培养表型差异;B-C:木霉与CR62、CR15共培养3天和5天的抑制率;D:NY-1(绿色箭头)对病原菌(CR62和CR15,红色箭头)的寄生显微观察(比例尺=50 μm);E-F:L1-21与CR62、CR15及木霉菌株(NY-1、CT1、DH2)的共培养表型与抑制率;G:L1-21、NY-1单独及共培养的生长特征;H:NY-1发酵滤液对L1-21生长(OD600)的影响;I:L1-21发酵滤液对木霉生长(CFU计数)的影响;J:单独培养与代谢互养(L1-21、NY-1)对CR62和CR15抑制率的影响;K:L1-21、NY-1及代谢互养对半叶法检测CLas菌量的影响;L:代谢组PCA分析显示L1-21(CK)与L1-21(NY-1)处理组明显分群;M:差异代谢物火山图(红色上调、蓝色下调、灰色无显著差异);N:关键代谢物(Top 10)表达热图(红色高表达、蓝色低表达);O:KEGG代谢通路富集分析(富集因子与P值)。NY-1发酵滤液显著促进L1-21生长(OD600提升1.5倍),而L1-21滤液对木霉菌落形成单位(CFU)无显著影响(图3H-I)。代谢互养条件下,L1-21与NY-1协同抑制CR62和CR15(抑制率分别达89.12 ±1.22%和82.35 ± 1.67%),显著高于单独处理(图3J)。半叶法检测显示,代谢互养处理组CLas菌量(Ct值28.5 ± 0.8)显著低于对照组(Ct值25.1 ± 0.6)(图3K)。代谢组PCA分析表明,L1-21(NY-1)处理组与对照组(CK)明显分群(图3L)。火山图筛选出126种差异代谢物(63种上调,63种下调)(图3M)。热图显示NY-1调控L1-21的10种关键代谢物(如L-谷氨酸、琥珀酸、D-葡萄糖胺-6-磷酸等)(图3N)。KEGG分析揭示这些代谢物显著富集于氨基酸代谢(苯丙氨酸、赖氨酸等)、能量代谢(TCA循环、氧化磷酸化)及ABC转运蛋白通路(P < 0.05)(图3O)。L1-21与NY-1在PDB培养基中共培养7天后,发酵液颜色与L1-21单独培养时相似,但二者在同一平板上的共生长现象表明其共享相同生态位(图3G)。NY-1无菌上清液(5-20%浓度)显著促进L1-21生长,其中10%浓度时促生效应最显著(图3H)。相反,L1-21无细胞上清液以浓度依赖性方式抑制NY-1生长。这一分化结果反映了不同培养条件下的互作模式差异:活细胞维持了合成群落(SynCom)构建的兼容性,而无细胞上清液则表现出拮抗活性,这与L1-21的生防潜力一致(图3I)。L1-21和NY-1单独培养的代谢产物对CR62和CR15的抑制效应随无菌滤液浓度增加而增强,其中L1-21滤液的拮抗活性强于NY-1。当L1-21在添加10% NY-1无菌滤液的培养基中培养时,其代谢物对CR15的抑制能力较单独培养显著提升;而NY-1在2.5% L1-21滤液条件下培养的代谢物抑制活性与单独培养无显著差异(图3J)。对HLB感染叶片的处理显示,L1-21降低CLas菌量的效果优于NY-1;与单菌处理相比,L1-21与NY-1联用或添加10% NY-1培养滤液时,CLas菌量相对降幅显著更高(图3K)。综合病原抑制实验表明,L1-21对柑橘根腐病病原菌及CLas的抑制活性强于NY-1,而NY-1代谢物可增强L1-21的抑制效应,此现象在同样抑制CLas的另两株木霉(CT1和DH2)中未观察到(图S2)。为解析NY-1无菌滤液增强L1-21生防活性的代谢基础,代谢组数据的多变量统计与通路富集分析显示,L1-21(CK)与添加10% NY-1滤液共培养的L1-21全局代谢谱存在显著差异。PCA分析表明两组明显分离,PC1和PC2分别解释70.30%和6.89%的变异度,组间差异具有统计学意义(R=1,P=0.041)(图3L)。火山图筛选出1,075种显著差异代谢物(574种上调,501种下调),2,945种代谢物无显著变化(图3M)。层次聚类与热图分析显示差异代谢物(如Leu-Pro-Gln和Val-Pro-Gln)具有特异性积累模式(图3N)。KEGG富集分析表明,差异代谢物主要关联氨基酸与能量代谢,其中精氨酸生物合成、赖氨酸降解、丙氨酸-天冬氨酸-谷氨酸代谢及ABC转运蛋白通路富集程度高(富集因子大)且显著性低(P值小)(图3O)。这些通路与支持生防的关键生理过程(细胞生长、抗逆性、营养竞争及拮抗物质分泌)密切相关,表明NY-1滤液不仅调控氨基酸代谢,还可能通过影响细胞内能量生成与物质运输全面改变细胞代谢活动。综上,这些全局代谢重排为L1-21生防效能的提升提供了关键代谢基础。所有微生物处理组的柑橘株高、叶片数、叶绿素及氮含量均随时间(30、60和90天)增加,且显著高于对照组。处理90天后,SynCom NL处理组的株高和叶片数较CK组分别提高10.95%和10.85%(图4A-B)。叶片生理参数中,各处理组的叶绿素含量(SPAD值)和叶片氮含量随处理时间持续上升,均显著高于CK组(图4C-D)。表型观察显示,处理组幼苗的茎叶生长加速、叶片密度增加且根系更发达,其中SynCom NL组的促生效果最显著(图4E-F)。图4. L1-21、NY-1及SynCom NL对盆栽柑橘幼苗生长的影响。A-D:不同处理组柑橘幼苗在处理前及处理后30、60、90天的株高(A)、叶片数(B)、叶绿素含量(C)和叶片氮含量(D);E-F:90天时各处理组植株表型(E)(标尺:30 cm)与根系生长表型(F)(标尺:10 cm)。(5)合成群落对柑橘根际土壤酶活性及理化性质的影响处理14天后,所有微生物接种组的土壤酶活性均显著高于处理前及CK组。具体而言,各接种组的土壤脲酶活性均上升,其中SynCom NL组活性最高(图5A);L1-21单菌与SynCom NL组的蔗糖酶活性达峰值(图5B)。单菌(L1-21和NY-1)及群落处理均显著提升碱性磷酸酶、脱氢酶和过氧化氢酶活性(图5C-E)。土壤理化性质与养分分析显示:各处理间pH无显著差异,但L1-21和NY-1组略高于CK组(图5F);NY-1与SynCom NL组的有机质含量显著高于处理前水平(图5G)。微生物处理组的总氮含量与处理前无显著差异,而CK组显著降低(图5H);水解性氮含量以SynCom NL组最高且显著优于其他处理,L1-21组最低(图5I)。总磷与有效磷含量处理后均显著增加(图5J-K)。NY-1处理组的总钾和速效钾含量与处理前相当,其余处理组显著下降(图5L-M)。结果表明,L1-21、NY-1及SynCom NL的应用可增强根际土壤酶活性、促进有机质分解并提高养分释放,从而促进柑橘植株生长。图5. L1-21、NY-1及SynCom NL对根际土壤酶活性与理化性质的影响。A-E:处理后14天各处理组的根际土壤酶活性,包括脲酶(A)、蔗糖酶(B)、碱性磷酸酶(C)、脱氢酶(D)和过氧化氢酶(E);F-M:处理前及处理后14天的土壤理化性质与养分含量,包括pH(F)、有机质(G)、总氮(H)、水解性氮(I)、总磷(J)、有效磷(K)、总钾(L)和速效钾(M)。(6)合成群落对柑橘根腐病的盆栽防控效果及根际微生态调控接种病原菌CR15 60天后,染病植株表现为地上部严重萎蔫落叶,根系稀疏褐变且腐烂;而经L1-21、NY-1或SynCom NL处理的植株茎叶生长改善,根系形态恢复良好,表现为须根发达、侧根增多及根色亮白,与Mock对照组相当(图6A)。CR15感染使根系表面积降至(2.41 ± 0.43)× 10⁴ mm²,较Mock对照组(6.52 ± 0.71)× 10⁴ mm²降低约63%。L1-21、NY-1及SynCom NL处理均增加根系表面积,其中SynCom NL组恢复程度最接近Mock水平(图6B)。CR15接种组的病情指数达46.67 ± 5.44,显著高于Mock组(0);L1-21、NY-1和SynCom NL处理分别将病情指数降至18.33 ± 6.38、6.67 ± 5.44和3.33 ± 3.85,其中SynCom NL的相对防效达92.86 ± 8.25%(图6C)。CR15、L1-21、NY-1或SynCom NL处理对抗氧化酶(SOD、POD、CAT)与防御相关酶(PAL、PPO)活性影响显著:与Mock组相比,CR15接种显著提升五种酶活性;而生防菌处理进一步增强了SOD、CAT、PAL和PPO活性,同时显著降低POD活性(图6D)。结果表明,生防菌通过直接拮抗病原菌和诱导植物系统抗性,缓解氧化损伤并维持根系健康生长。CR15与NY-1处理均未显著改变根际细菌Shannon指数,而L1-21显著降低该指数;合成群落NL则部分恢复了细菌多样性(图6E)。所有处理均显著降低根际真菌Shannon指数(图6F)。PCoA分析显示,不同处理组的细菌与真菌群落样本明显分群,表明群落组成存在显著差异(图6G-H)。图6. L1-21、NY-1及SynCom NL对CR15所致柑橘根腐病的防控效果及其对根系酶活性与根际微生物群落的影响。A:L1-21、NY-1和SynCom NL处理后的柑橘植株表型(盆栽植株与根系图像标尺分别为30 cm和10 cm);B-D:L1-21、NY-1及SynCom NL对根系总表面积(B)、根腐病病情指数与相对防效(C)、抗氧化与防御相关酶活性(D)的影响;E-F:基于Shannon指数的根际细菌(E)与真菌(F)群落α多样性分析;G-H:细菌(G)和真菌(H)群落组成的PCoA分析;I:属水平根际细菌相对丰度(前15属);J-K:鞘氨醇单胞菌属(J)与芽孢杆菌属(K)相对丰度;L:属水平根际真菌相对丰度(前15属);M-N:镰刀菌属(M)与木霉属(N)相对丰度。各处理设4次重复;统计学显著性基于单因素方差分析后的Tukey HSD检验,标注为*P < 0.05、**P < 0.01、***P < 0.001。属水平细菌群落组成在各处理间差异显著:与Mock对照组相比,各处理组的优势类群呈现特异性变化。鞘氨醇单胞菌属(Sphingomonas)为最优势菌属,其次为芽孢杆菌属(Bacillus)(图6I)。相对丰度前10的菌属在各处理间差异显著(图S3A)。鞘氨醇单胞菌属作为根际细菌群落的关键类群,在CR15 + L1-21处理中相对丰度显著增加(P = 0.0424),表明L1-21施用促进了其在根际的富集(图6J)。芽孢杆菌属的相对丰度在CR15 + L1-21处理中达到峰值,且在NY-1与L1-21联用后仍维持较高水平(图6K)。共现网络分析显示,Fs + L1-21处理的相关节点数最少,表明枯草芽孢杆菌L1-21的定殖与扩增与其拮抗特性相关(图S3C)。接种CR15并施用生防菌后,各处理组的根际真菌群落结构差异显著(图6L)。与Mock组相比,CR15接种显著增加镰刀菌属(Fusarium)丰度,而生防处理使其显著降低,其中CR15 + NY-1与CR15 + NL处理降幅最大(图6M)。木霉属(Trichoderma)丰度在NY-1处理后显著上升,且CR15 + NY-1组显著高于CR15 + NL组(图6N)。木霉属相对丰度与镰刀菌属呈显著负相关,证实棘孢木霉NY-1是抑制镰刀菌的主要驱动因子。此外,SynCom NL处理中潜在植物病原菌属(如青霉属Penicillium、新赤壳属Neocosmospora)丰度降低,而部分拮抗性或环境耐受性真菌(如腐质霉属Humicola、Torula属)丰度上升(图S3B)。L1-21、NY-1及SynCom NL的调控作用改变了真菌群落的节点数量(图S3D)。综上,L1-21、NY-1及SynCom NL的施用显著重塑了根际细菌与真菌群落,多维响应模式表明L1-21与NY-1存在交互作用,导致根际原生微生物群落的竞争关系与生态位占据发生改变。尽管跨界合成群落(SynCom)具有生态复杂性,本研究成功验证了一种基于生态学原理的可行管理策略。田间条件下,柑橘植株常同时感染黄龙病(HLB)与根腐病病原菌,对植株健康产生复合危害,其中烟草疫霉(Phytophthora nicotianae)和茄病镰刀菌(Fusarium solani)被确认为主要根腐病病原菌。棘孢木霉NY-1对这些病原菌表现出强寄生与致死活性,并通过调控L1-21的氨基酸与能量代谢,促进跨界合成群落NL内的代谢互养与病原菌协同抑制。SynCom NL的应用显著提升了根系抗氧化与防御酶活性,增加了土壤酶活性,富集了有益微生物(如芽孢杆菌属Bacillus、鞘氨醇单胞菌属、木霉属及腐质霉属),同时降低了病原菌丰度(如镰刀菌属、青霉属及新赤壳属),从而形成"土壤改良-有益微生物增殖-病原菌抑制-植物防御增强"的自我强化循环。值得注意的是,未来需进一步探究此类跨界合成群落在长期田间试验中的表现,以及已构建群落在病害进展中的稳定性。综上,这些效应协同缓解了生物胁迫并恢复植株健康,证明跨界合成群落可作为柑橘黄龙病-根腐病复合病害综合治理的有效且可持续策略。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
