3.1铝毒诱导白三叶中活性氧的积累
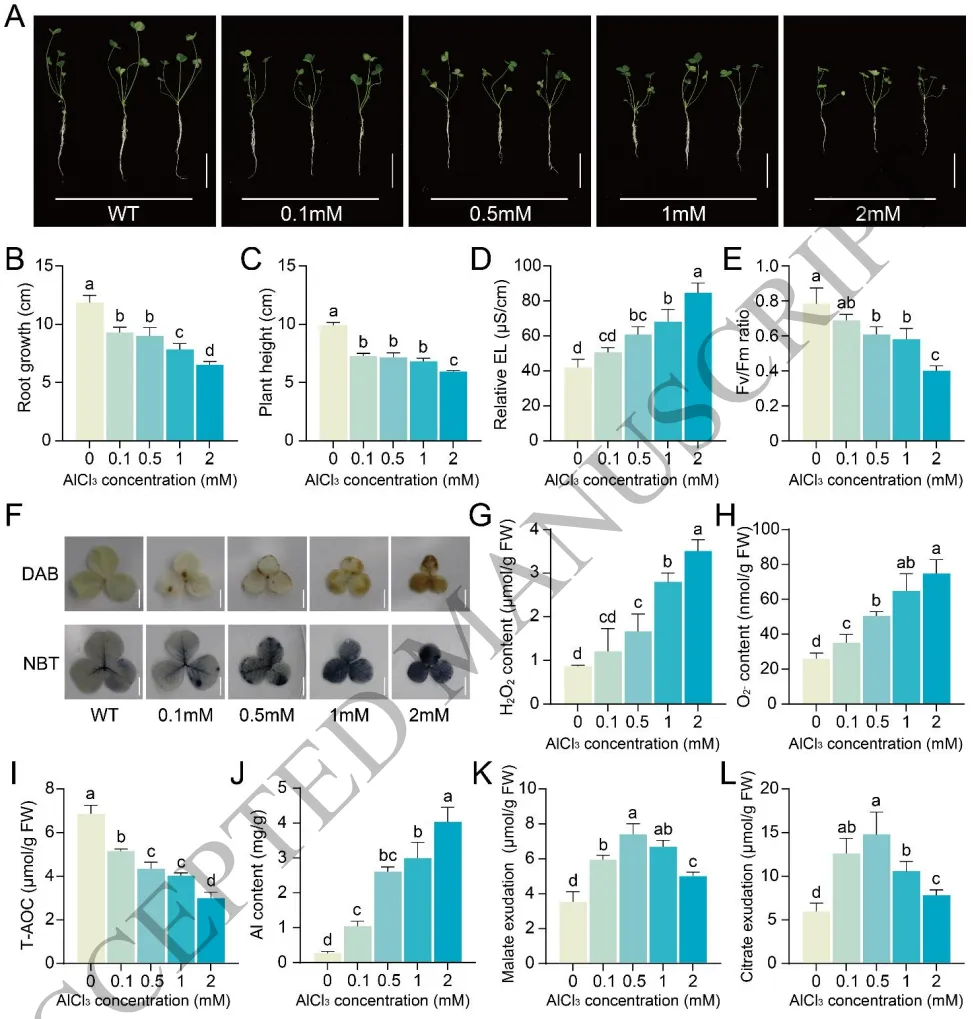
生长抑制:铝胁迫显著抑制白三叶幼苗的根长、株高及生物量积累,且呈剂量依赖性。
生理损伤:铝毒导致电解质渗漏增加和光合效率下降,并引发根和叶中活性氧及丙二醛的大量积累,降低总抗氧化能力。
有机酸分泌:低浓度铝促进苹果酸和柠檬酸分泌以螯合铝离子,但在高浓度下该解毒途径发生崩溃。
评估白三叶对铝(Al)毒害的响应发现,铝胁迫对幼苗生长产生显著的剂量依赖性抑制作用。随着铝浓度的增加,植物光合功能严重受损,细胞膜通透性显著增加,并在根和叶组织中诱发强烈的活性氧(H₂O₂和O₂⁻)积累及氧化损伤。尽管植物在低浓度铝胁迫下能够通过增加根系苹果酸和柠檬酸的分泌来螯合毒性Al³⁺,但高浓度铝会导致该解毒机制全面崩溃,从而加剧铝在根系的富集与毒害。(Fig.1)
图1. 铝毒诱导白三叶中活性氧的积累。 铝处理下白三叶幼苗的表型及根长、生物量等生长指标变化;电解质渗漏和PSII最大光化学效率的测定;叶片和根系中活性氧(H₂O₂和O₂⁻)及丙二醛的积累与组织化学染色分析;总抗氧化能力的变化;根系铝含量及苏木精染色结果;不同浓度铝处理下根系分泌的苹果酸和柠檬酸含量测定。
3.2 白三叶响应铝毒的转录组分析
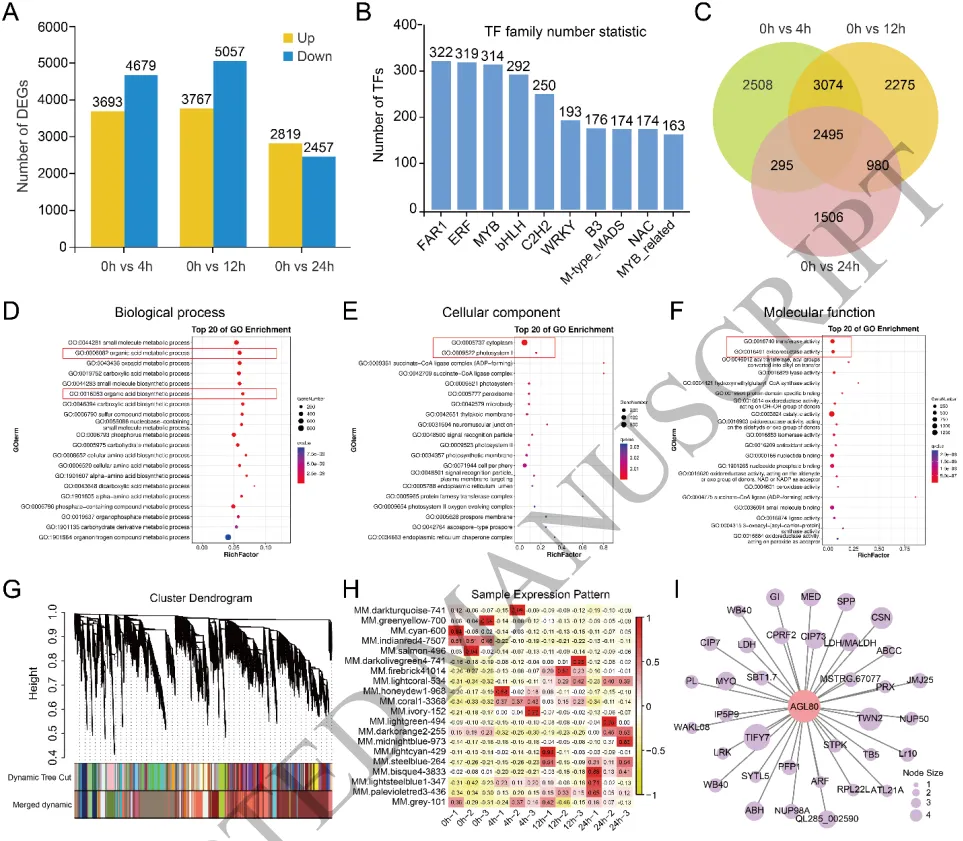
差异表达:转录组测序鉴定出大量随时间变化的差异表达基因,主要富集于FAR1、ERF和MYB转录因子家族。
功能富集:差异基因在有机酸代谢与生物合成、氧化还原酶活性等途径中表现出显著富集。
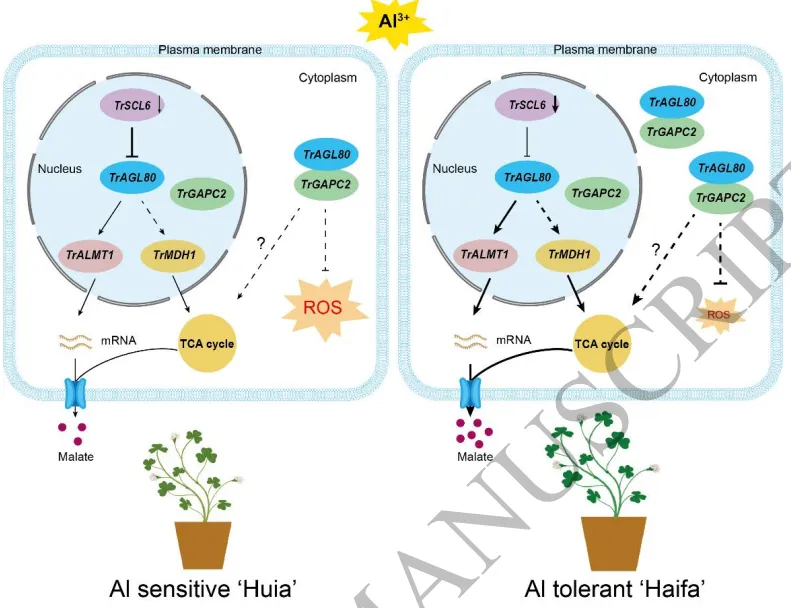
共表达网络:WGCNA分析识别出与铝胁迫高度相关的共表达模块,并鉴定出核心转录因子TrAGL80及TCA循环关键基因。
为深入解析白三叶响应铝毒的分子机制,对铝胁迫下的根系进行了时间序列转录组测序,鉴定出大量差异表达基因。功能富集分析表明,这些基因主要参与有机酸代谢、生物合成以及氧化还原酶活性调节,提示有机酸合成与活性氧调控在抗铝毒中发挥关键作用。进一步的加权基因共表达网络分析(WGCNA)识别出一个与铝胁迫持续时间高度相关的基因模块,并成功鉴定出该模块的核心转录因子TrAGL80及其共表达的三羧酸循环关键基因(TrLDH和TrMALDH),暗示TrAGL80可能通过调控TCA循环相关基因的表达来增强植物耐铝性。(Fig.2)
图2. 白三叶响应铝毒的转录组分析。 铝处理不同时间点根系差异表达基因的数量统计与转录因子家族分类;差异表达基因的Venn图及GO和KEGG功能富集分析;加权基因共表达网络分析(WGCNA)识别出的模块与铝胁迫时间的关联;MM.bisque4模块的核心基因TrAGL80及其共表达网络。
3.3 白三叶中TrAGL80的鉴定与表征
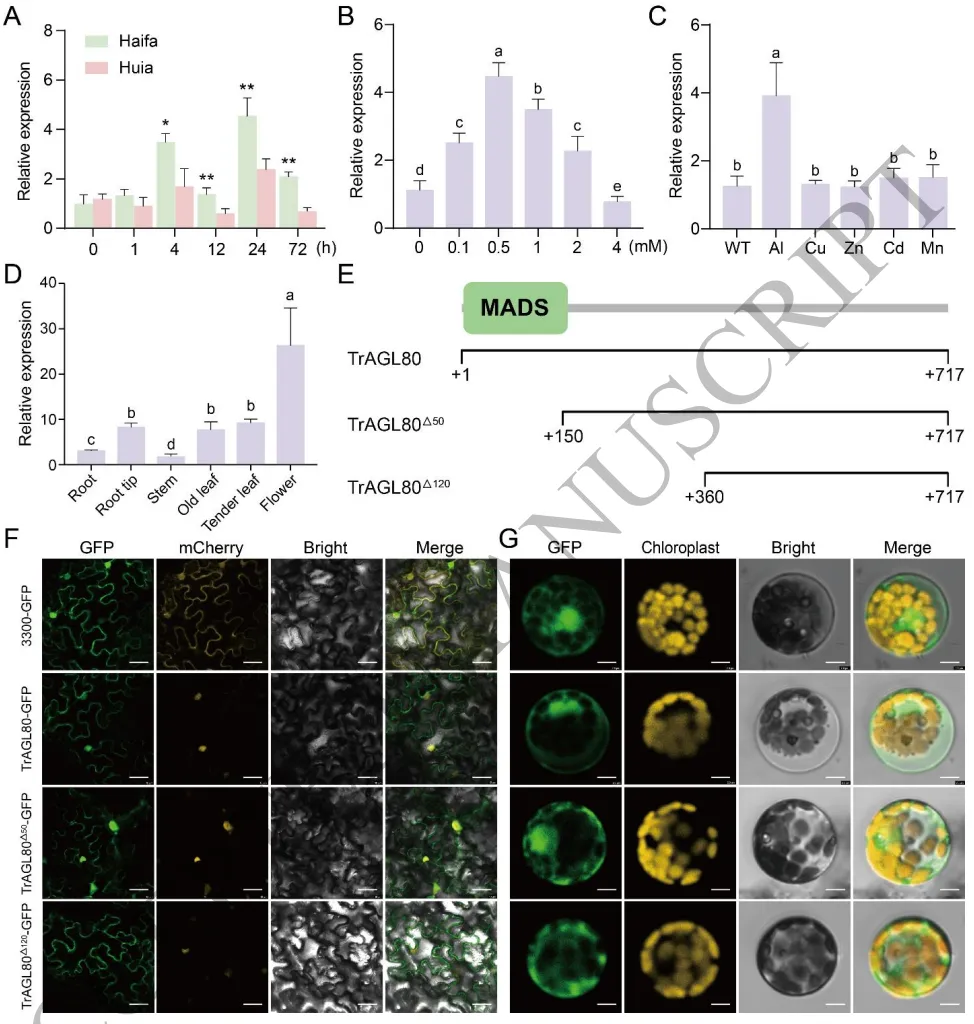
基因鉴定:全基因组鉴定出151个TrMADS基因,系统发育分析显示TrAGL80属于I型Mγ进化分支。
表达模式:TrAGL80在铝胁迫早期被显著且特异性地诱导表达,并在花组织中高表达。
亚细胞定位:TrAGL80同时定位于细胞核与细胞质,其C端结构域是决定细胞质定位的关键。
结合转录组数据与生物信息学分析,在白三叶全基因组中鉴定出151个MADS-box家族成员,其中TrAGL80被归类为进化上保守的I型Mγ分支成员。表达谱定量分析显示,TrAGL80是一个重要的铝胁迫响应基因,在受铝毒刺激后迅速且显著地上调表达。亚细胞定位实验证实,TrAGL80不仅定位于细胞核,还存在于细胞质中,且其C端序列是引导其实现细胞质定位的关键功能结构域。(Fig.3)
图3. 白三叶中TrAGL80的鉴定与表征。 耐铝和敏感品种在铝胁迫下TrAGL80的表达模式差异;不同浓度铝及不同金属处理下TrAGL80的特异性诱导表达;TrAGL80的组织特异性表达分析;TrAGL80及其截短突变体在本氏烟草叶片和白三叶原生质体中的亚细胞定位。
3.4 TrAGL80通过调节TrALMT1增加苹果酸分泌从而增强白三叶的铝耐受性
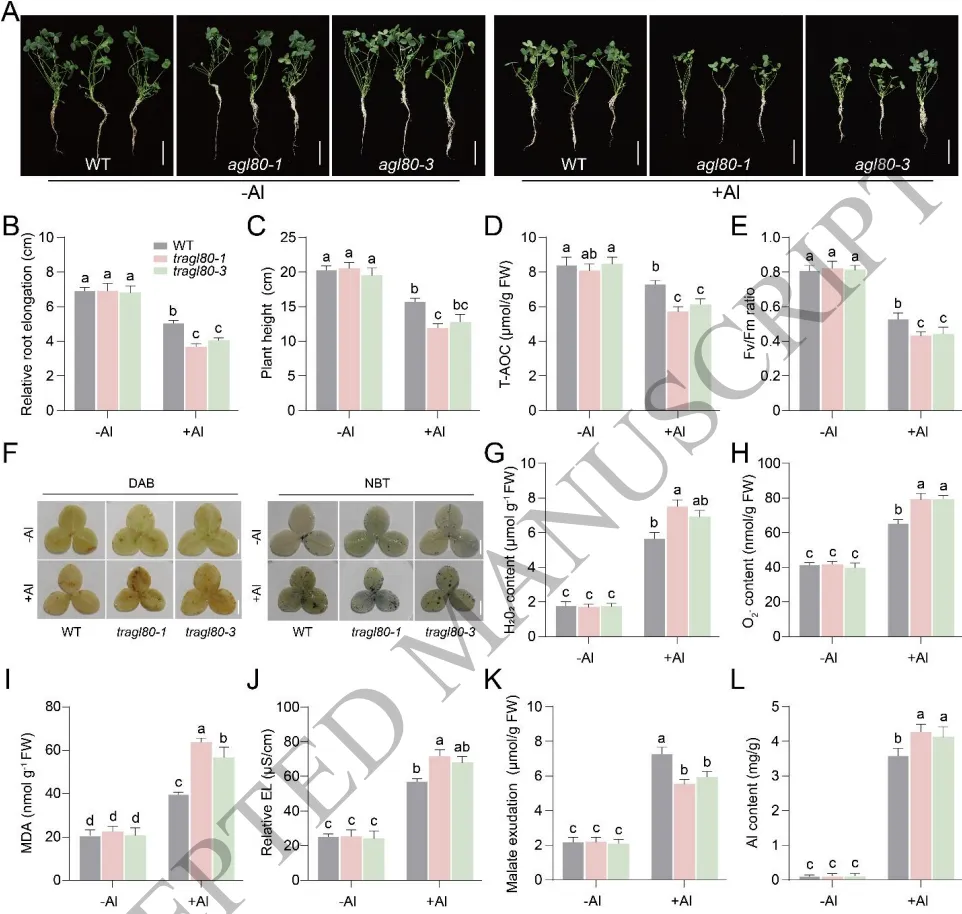
正向调控:TrAGL80干扰或沉默株系在铝胁迫下表现出更严重的生长抑制和强烈的氧化损伤。
促进分泌:TrAGL80表达受抑导致根系苹果酸分泌显著减少,进而引发根部铝积累大幅增加。
靶向激活:TrAGL80直接结合TrALMT1启动子并激活其表达,从而在转录水平调控苹果酸分泌。
TrAGL80在耐铝品种中的表达量显著高于敏感品种,表明其在铝耐受性中具有功能保守性。通过构建RNAi干扰和VIGS沉默株系发现,抑制TrAGL80表达会严重削弱植物清除活性氧和分泌苹果酸的能力,导致剧烈的氧化损伤和根系Al³⁺大量积累,从而加剧植物的铝敏感性。分子互作实验进一步证实,TrAGL80能够直接结合铝激活的苹果酸转运蛋白基因TrALMT1的启动子并正向激活其转录,进而通过促进苹果酸分泌来显著增强白三叶的耐铝性。(Fig.4)
图4. TrAGL80通过调节TrALMT1增加苹果酸分泌增强白三叶铝耐受性。TrAGL80-RNAi株系在铝胁迫下的表型及根长、株高统计;生理指标(T-AOC、Fv/Fm、MDA、EL)的测定;活性氧(H₂O₂和O₂⁻)积累及组织化学染色;根系铝含量和苹果酸分泌量的定量分析。
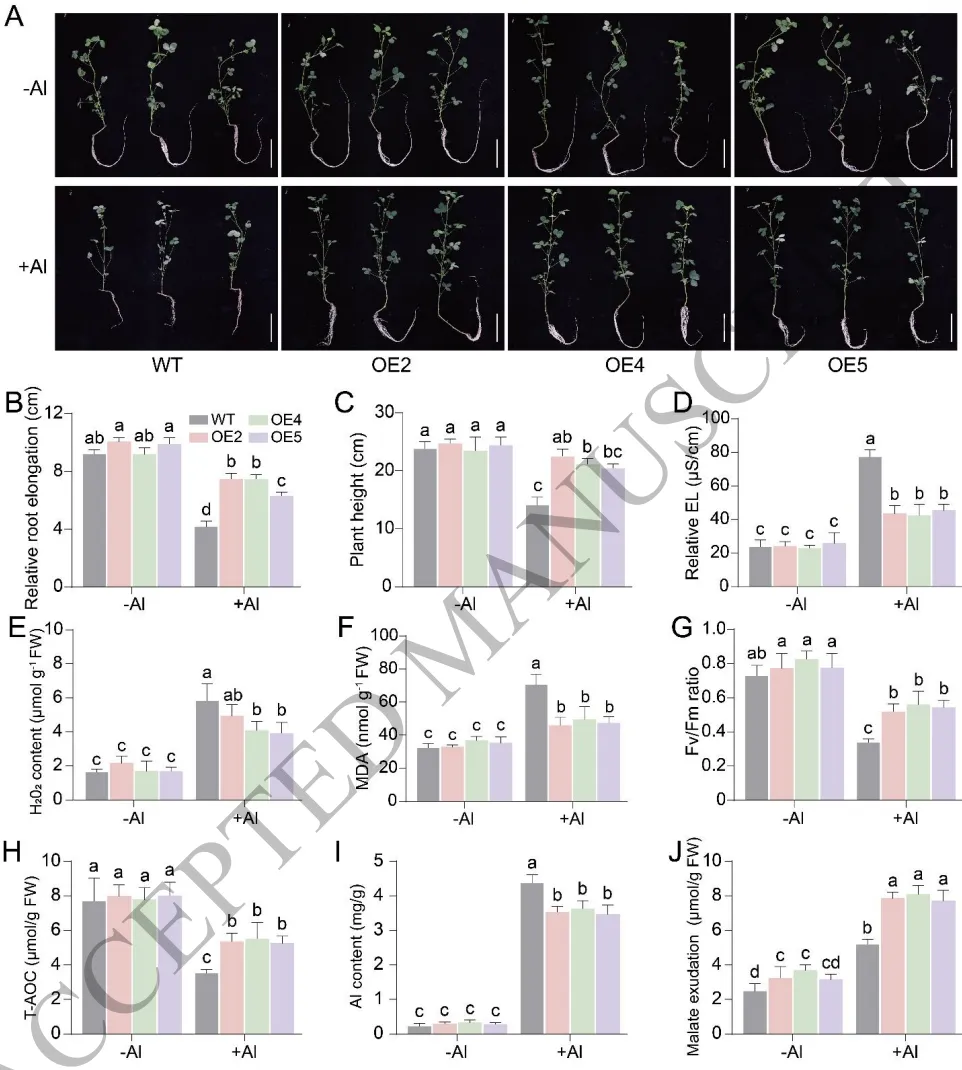
3.5 TrAGL80的异源表达提高紫花苜蓿的铝耐受性
异源表达:在紫花苜蓿中过表达TrAGL80显著改善了铝胁迫下的根系伸长和植株生长状态。
缓解损伤:过表达株系在铝处理下的活性氧积累和膜脂过氧化水平显著降低,抗氧化能力提升。
增强解毒:转基因株系根尖的苹果酸分泌量显著增加,有效减少了毒性铝离子在根部的积累。
为验证TrAGL80的抗铝功能,将其在紫花苜蓿中进行异源过表达。在铝胁迫下,过表达TrAGL80的转基因紫花苜蓿幼苗表现出更优的根系发育和植株生长表型。生理生化分析表明,转基因植株体内的活性氧积累和氧化损伤显著降低,总抗氧化能力明显提升。同时,异源表达TrAGL80显著促进了根尖苹果酸的分泌,从而大幅减少了根系中的铝积累。这表明TrAGL80通过维持氧化还原稳态和增加有机酸分泌,有效提升了紫花苜蓿的铝耐受性。(Fig.5)
图5. TrAGL80的异源表达提高紫花苜蓿的铝耐受性。 过表达TrAGL80转基因紫花苜蓿株系在铝胁迫下的表型、根长和株高;电解质渗漏、H₂O₂、MDA含量及Fv/Fm、T-AOC的测定;根尖铝含量及苹果酸分泌量的定量分析。
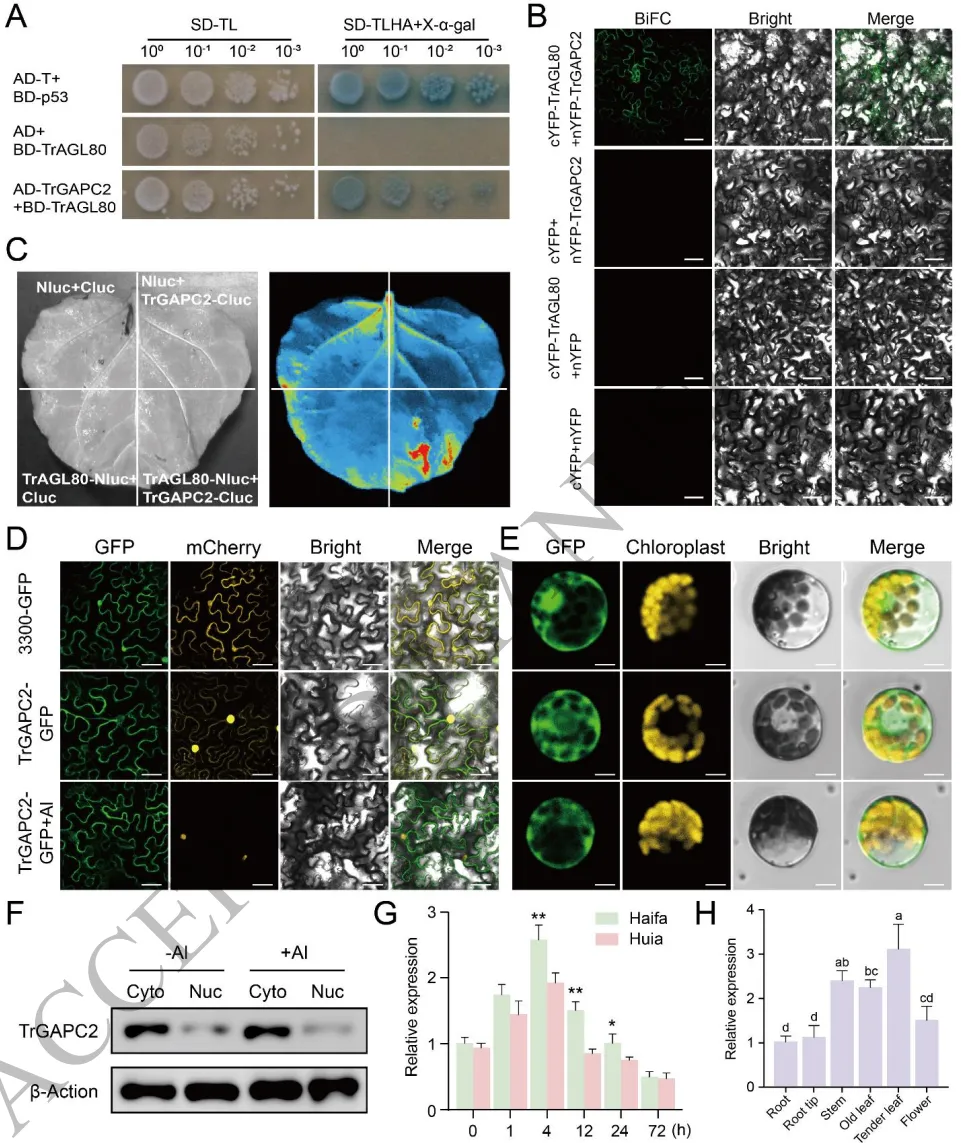
3.6 TrAGL80与TrGAPC2发生物理相互作用
蛋白互作:Y2H、BiFC和LCA实验一致证实TrAGL80与细胞质酶TrGAPC2存在直接物理互作。
表达特征:TrGAPC2在耐铝品种中表达量更高,受铝胁迫诱导,且主要在幼叶组织中高表达。
亚细胞定位:TrGAPC2定位于细胞核与细胞质中,且铝胁迫并未诱导其发生向细胞核的易位。
为了解析TrAGL80发挥功能的分子网络,通过酵母双杂交文库筛选鉴定出其互作蛋白TrGAPC2(一种细胞质3-磷酸甘油醛脱氢酶)。酵母双杂交、双分子荧光互补(BiFC)和荧光素酶互补(LCA)实验共同证实了TrAGL80与TrGAPC2在细胞质中发生直接的物理相互作用。表达谱分析显示,TrGAPC2在耐铝品种中的表达水平显著高于敏感品种。尽管前期研究表明GAPC在某些胁迫下会发生核易位,但亚细胞定位和免疫印迹分析表明,铝胁迫并未诱导TrGAPC2在白三叶中向细胞核转移。(Fig.6)
图6. TrAGL80与TrGAPC2发生物理相互作用。 酵母双杂交(Y2H)验证TrAGL80与TrGAPC2的互作;双分子荧光互补(BiFC)和荧光素酶互补(LCA)实验在植物细胞中证实互作;TrGAPC2在原生质体和烟草叶片中的亚细胞定位及铝处理下的免疫印迹分析;TrGAPC2在不同品种和组织中的表达模式。
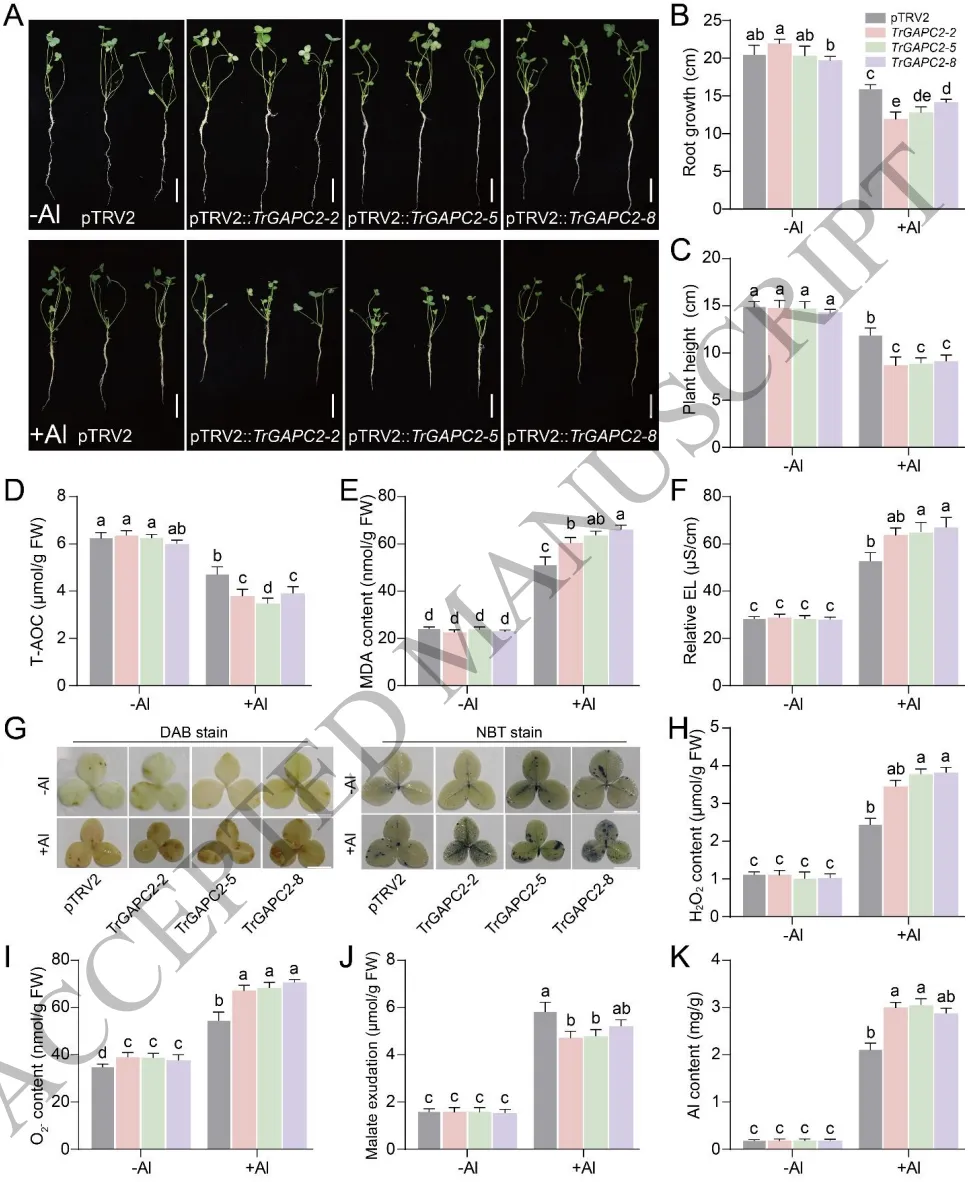
3.7 沉默TrGAPC2通过增加活性氧积累和减少苹果酸分泌提高铝敏感性
加剧毒害:沉默TrGAPC2导致白三叶在铝胁迫下生长严重受抑,活性氧大量积累且氧化损伤加剧。
抑制分泌:TrGAPC2表达沉默显著降低了根尖的苹果酸分泌,导致根组织中铝富集量急剧增加。
代谢偶联:TrGAPC2沉默下调了苹果酸合成关键基因TrMDH1的表达,导致细胞质苹果酸含量降低。
通过病毒诱导的基因沉默(VIGS)技术验证了TrGAPC2在铝耐受中的核心功能。在铝胁迫下,沉默TrGAPC2的植株表现出极度敏感的表型,根系生长严重受阻,体内活性氧(H₂O₂和O₂⁻)大量积累,抗氧化防御系统受损。此外,TrGAPC2的缺失显著抑制了根尖苹果酸的分泌,导致根组织中铝的大量富集。进一步分析表明,沉默TrGAPC2会显著下调苹果酸脱氢酶基因TrMDH1的表达,从而减少细胞质苹果酸的合成。结合酶活性测定结果,提示TrAGL80可能通过增强TrGAPC2酶活性来促进内源苹果酸的积累,进而提升根系的铝耐受性。(Fig.7)
图7. 沉默TrGAPC2通过增加活性氧积累和减少苹果酸分泌提高铝敏感性。TrGAPC2沉默株系在正常和铝胁迫条件下的表型及生长指标;生理指标(T-AOC、EL、MDA)及活性氧(H₂O₂和O₂⁻)积累的测定与组织化学染色;根尖苹果酸分泌量、铝含量及苏木精染色分析。
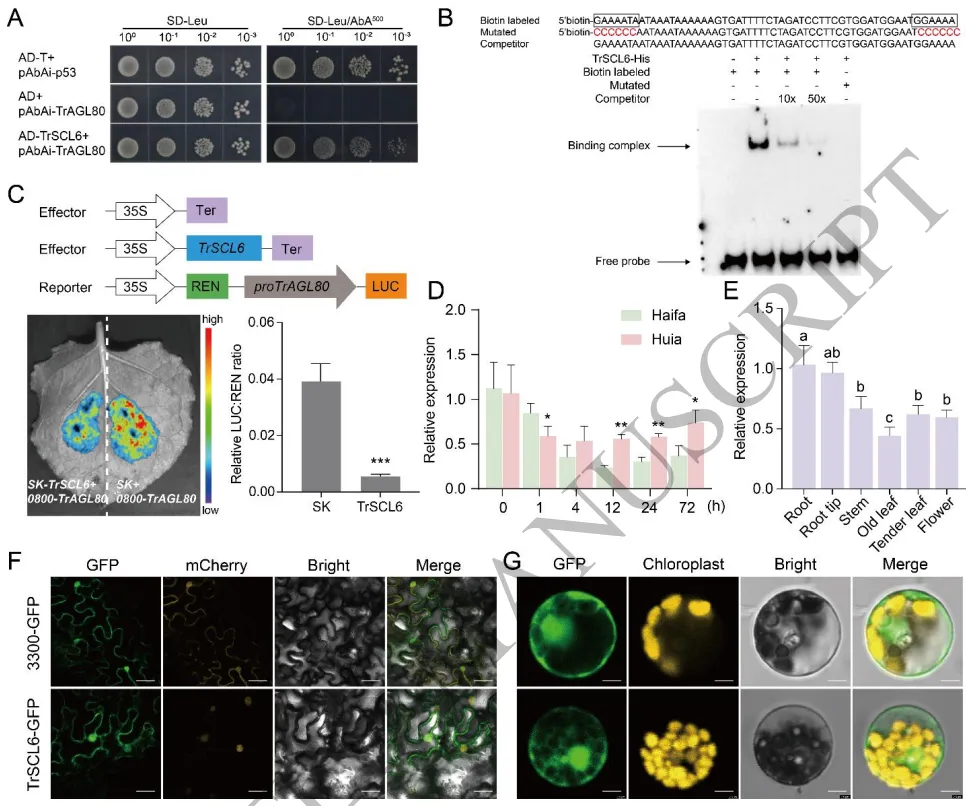
3.8 TrAGL80受GRAS家族成员TrSCL6的负向调控
上游调控:酵母单杂交和EMSA证实GRAS家族转录因子TrSCL6能够结合TrAGL80的启动子区域。
转录抑制:TrSCL6特异性结合GT-box基序,并在转录水平对TrAGL80的表达发挥强烈的抑制作用。
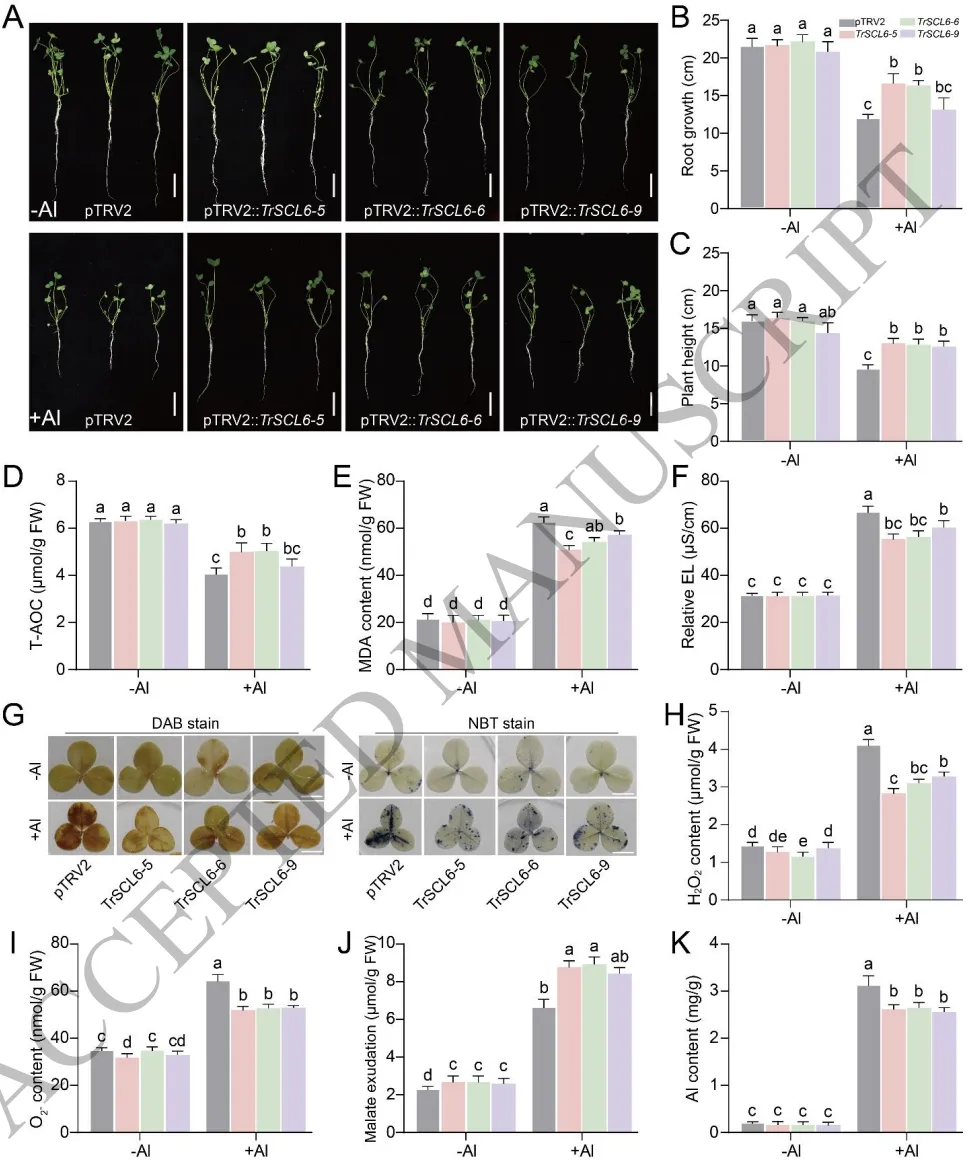
负向调节:沉默TrSCL6可显著减轻铝诱导的氧化损伤,增加苹果酸分泌,从而提高植物铝耐受性。
为探寻TrAGL80的上游调控网络,通过酵母单杂交筛选鉴定出GRAS家族转录因子TrSCL6。凝胶阻滞试验(EMSA)和双荧光素酶报告实验证实,TrSCL6能够直接特异性地结合到TrAGL80启动子区域的GT-box基序上,并对其转录表达发挥强烈的抑制作用。在铝胁迫初期,TrSCL6的表达受到显著抑制,从而解除了对TrAGL80的转录遏制。功能验证表明,沉默TrSCL6能够显著改善植物在铝胁迫下的生长状态,降低活性氧积累和细胞膜损伤,同时大幅增加根尖的苹果酸分泌并减少铝富集。这表明TrSCL6作为负调控因子,通过直接靶向抑制TrAGL80的表达来精细调节白三叶的铝耐受性。(Fig.8) (Fig.9) (Fig.10)
图8. TrAGL80受GRAS家族成员TrSCL6的负向调控。 酵母单杂交(Y1H)和凝胶阻滞试验(EMSA)验证TrSCL6与TrAGL80启动子的结合;双荧光素酶(dual-LUC)报告实验检测转录抑制活性;TrSCL6在铝胁迫下的表达模式及组织特异性表达;TrSCL6在原生质体和烟草叶片中的亚细胞定位。
图9. 沉默TrSCL6增强白三叶的铝耐受性。TrSCL6沉默株系在铝胁迫下的表型及生长指标;生理指标(T-AOC、H₂O₂、O₂⁻、MDA、EL)的定量测定与组织化学染色;根尖苹果酸分泌量及铝含量的分析。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
