3.1保留上部叶片(L3)对盐碱胁迫下燕麦的存活至关重要
表型差异:盐碱胁迫下,敏感品种DY5表型无显著变化,而耐受品种BY14地上部株高显著降低,表明抑制地上部生长可能是增强耐受性的适应性资源分配策略。
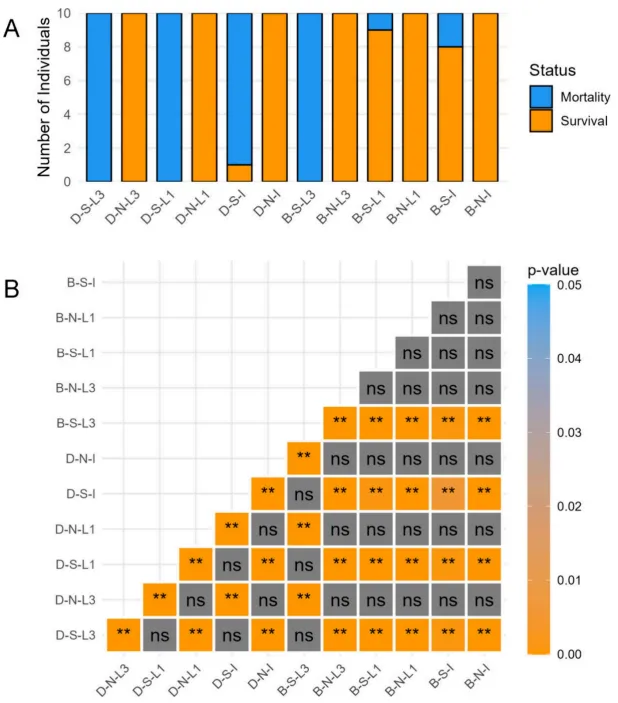
叶位效应:切除L3比切除L1对两种燕麦的生长损害更大。在胁迫下,切除L3的BY14全部死亡,而切除L1的植株存活率与完整对照无异;DY5的所有切除组及对照组最终均死亡,但切除L1的症状进展较慢。
胁迫依赖:在无胁迫条件下,切除L1或L3对植株生长均无显著不良影响,表明叶位切除效应依赖于特定胁迫环境。
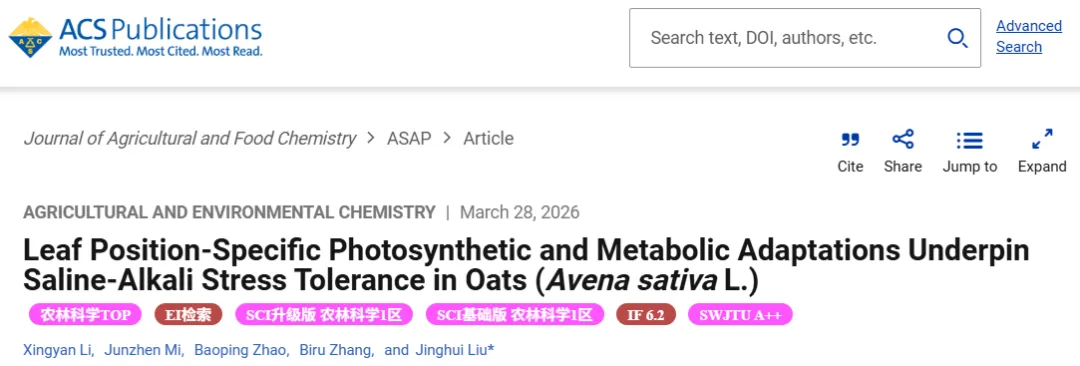
在盐碱胁迫下,耐受品种BY14通过抑制地上部生长展现出适应性资源分配策略。叶片切除实验揭示了叶位依赖的胁迫响应:保留上部叶片(L3)对植株存活起决定性作用。胁迫下切除L3导致BY14全部死亡,而切除下部叶片(L1)的植株存活率与未切除对照相当。敏感品种DY5无论是否切除叶片最终均无法存活,但保留L3能延缓衰亡。非胁迫条件下切除叶片无此负面效应,证实了上部叶片在应对盐碱胁迫中的核心地位(Fig.1)(Fig.2)。
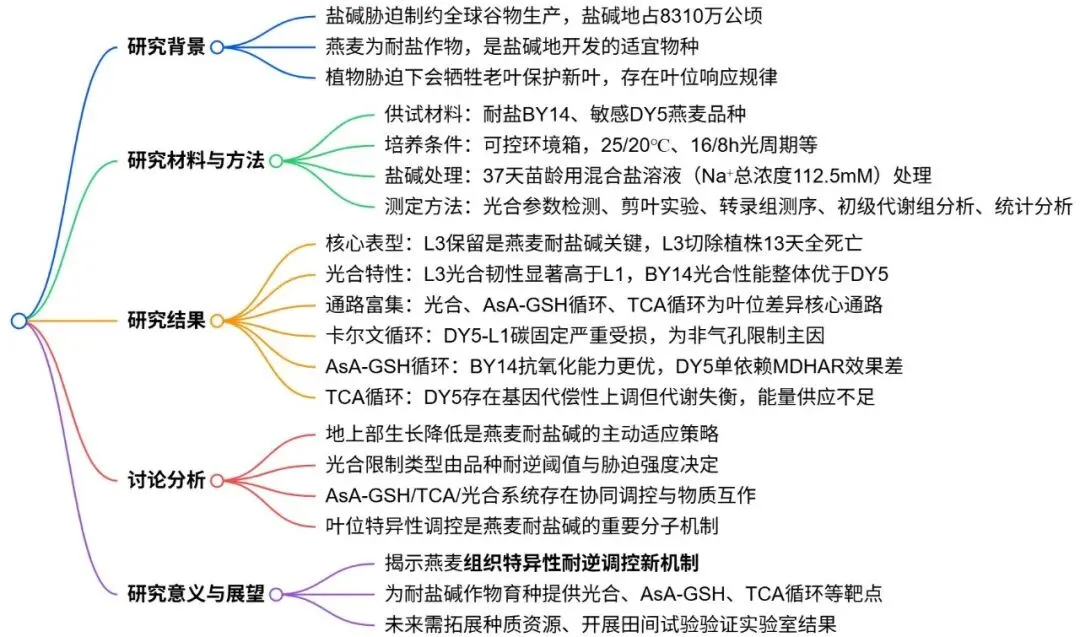
图1. 叶片切除实验的存活率统计。 盐碱敏感燕麦品种DY5(D-)和耐受品种BY14(B-)在盐碱胁迫(S)和无胁迫(N)条件下,包括切除L1和L3处理及完整(I)植株对照的叶片切除实验统计结果(A)及相应的Fisher精确检验矩阵(B)。**:p < 0.01。
图2. 耐盐碱和敏感燕麦品种在无胁迫和盐碱胁迫条件下切除叶片的生理后果。 子图比较处理如下:(A)DY5在无胁迫(D-N_d0)与胁迫(D-S_d0)切除前。(B)切除后2天的DY5切除L3(D-S-L3_d2)与DY5切除L1(D-S-L1_d2)。(C)BY14在无胁迫(B–N_d0)与胁迫(B–S_d0)切除前。(D)切除后2天的BY14切除L3(B–S-L3_d2)与BY14切除L1(B–S-L1_d2)。(E,F)切除后13天DY5在无胁迫(E)和胁迫(F)下的存活情况,比较完整对照(I)、切除L3(L3)和切除L1(L1)。(G,H)切除后13天BY14在无胁迫(G)和胁迫(H)下的存活情况,比较完整对照(I)、切除L3(L3)和切除L1(L1)。代码说明:“D-/B-” = DY5/BY14;“S/N” = 胁迫/无胁迫;“L3/L1/I” = L3/L1/完整对照;“d0/d2/d13” = 切除前/后天数。白色比例尺代表15 cm高度。
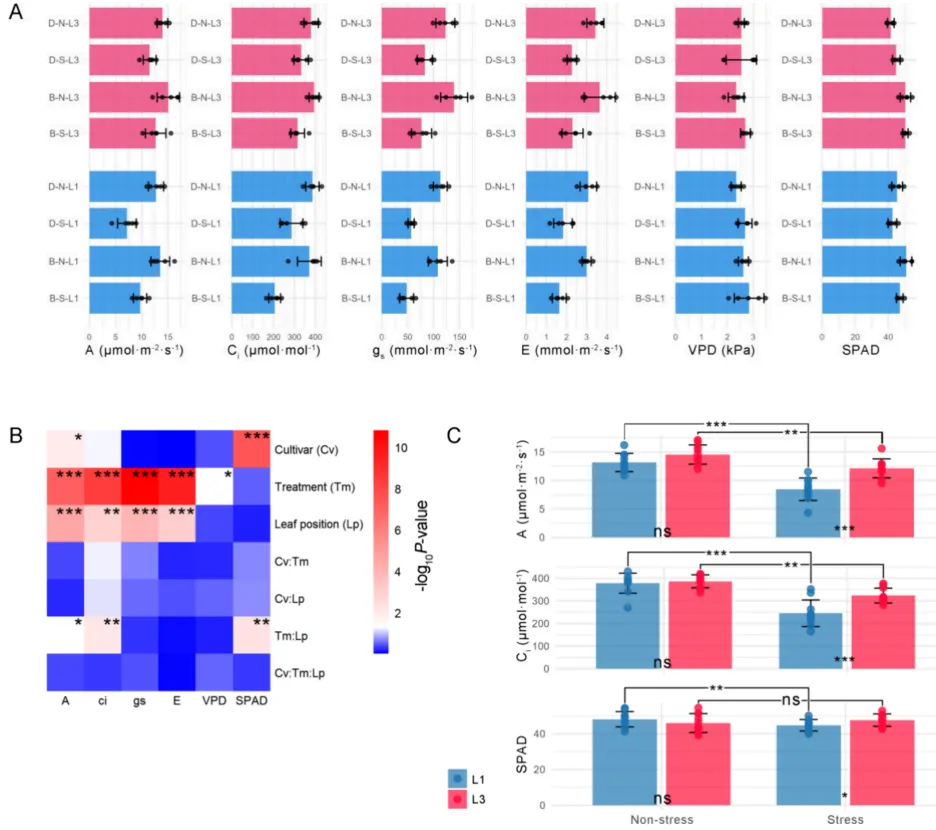
3.2. 对盐碱胁迫的光合响应差异:燕麦的品种和叶位特异性效应
同化速率:胁迫显著降低了L1和L3的同化速率,但L1的降幅更严重。DY5-L1的同化速率损失远高于其他组。胁迫下L3的同化速率显著高于L1,且耐受品种BY14的整体光合能力强于敏感品种DY5。
气孔导度:胁迫和叶位独立影响气孔导度(gs)和蒸腾速率(E)。胁迫显著降低了gs和E,且在所有处理中,L3的gs和E均显著高于L1。
胞间浓度:胁迫导致L1的胞间CO₂浓度(Ci)降幅远大于L3。DY5-L1的Ci降幅小于BY14-L1,结合其更显著的同化速率下降和较小的gs降幅,表明DY5-L1光合效率的降低更多由非气孔因素引起。
叶绿素值:品种是影响相对叶绿素含量(SPAD值)的主导因素。胁迫显著降低了L1的SPAD值,但对L3无显著影响,导致胁迫下L1和L3之间出现显著差异。
盐碱胁迫诱导了显著的差异性光合响应,受处理、叶位和品种的共同影响。胁迫大幅降低了光合参数,但上部叶片(L3)表现出比下部叶片(L1)更强的光合韧性,维持了更高的同化速率、气孔导度和相对叶绿素含量。耐受品种BY14展现出优于敏感品种DY5的整体光合性能。特别是DY5的L1叶片在胁迫下同化速率急剧下降,且主要受非气孔因素限制,突显了叶位和品种特异性的光合适应机制(Fig.3)。
图3. 光合气体交换参数和SPAD值统计。 盐碱耐受(BY14,包括子组B–N-L3、B–N-L1、B–S-L3、B–S-L1)和敏感(DY5,包括子组D-N-L3、D-N-L1、D-S-L3、D-S-L1)燕麦品种之间光合气体交换参数的比较,包括同化速率(A)、胞间CO₂浓度(Ci)、气孔导度(gs)、蒸腾速率(E)和蒸汽压亏缺(VPD),以及SPAD值(相对叶绿素含量)。实验条件包括正常(N-)和盐碱胁迫(S-)处理,区分上部(L3)和下部(L1)叶片;面板显示:(A)BY14和DY5的L3(红色柱)和L1(蓝色柱)中各参数的统计;(B)三因素ANOVA结果;(C)对净同化速率(A)、Ci和SPAD值中具有显著“处理 × 叶位”交互作用的指标进行简单效应分析,随后进行Tukey事后检验。、和分别表示p < 0.05、p < 0.01和p < 0.001。
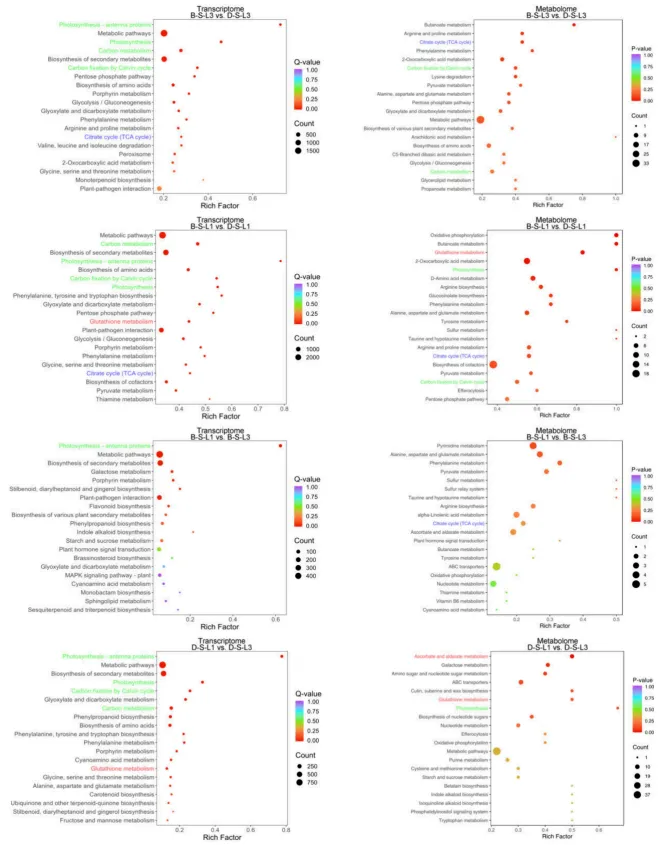
3.3. KEGG分析揭示关键代谢途径
为阐明盐碱胁迫下叶位差异性功能的机制,进行了RNA测序和初级代谢组学研究。通过对DY5和BY14不同叶位及品种间的KEGG富集分析,发现差异基因和代谢物主要集中在光合作用、抗坏血酸-谷胱甘肽(AsA-GSH)循环以及三羧酸(TCA)循环途径,揭示了这些代谢通路在调控燕麦耐盐碱性中的关键作用(Fig.4)。
图4. 盐碱胁迫下DY5和BY14的L1和L3叶片转录组和代谢组的KEGG富集分析。 盐碱敏感DY5(D-S-UL3:盐碱胁迫,L3;D-S-L1:盐碱胁迫,L1)和耐受BY14(B–S-L3:盐碱胁迫,L3;B–S_L1:盐碱胁迫,L1)燕麦品种转录组和代谢组的KEGG富集分析。显著富集的途径用颜色表示:绿色代表光系统相关途径,红色代表抗坏血酸-谷胱甘肽(AsA-GSH)相关途径,蓝色代表TCA循环途径。Rich Factor表示某途径中差异代谢物与该途径注释的代谢物总数的比值。p值对应超几何检验结果。
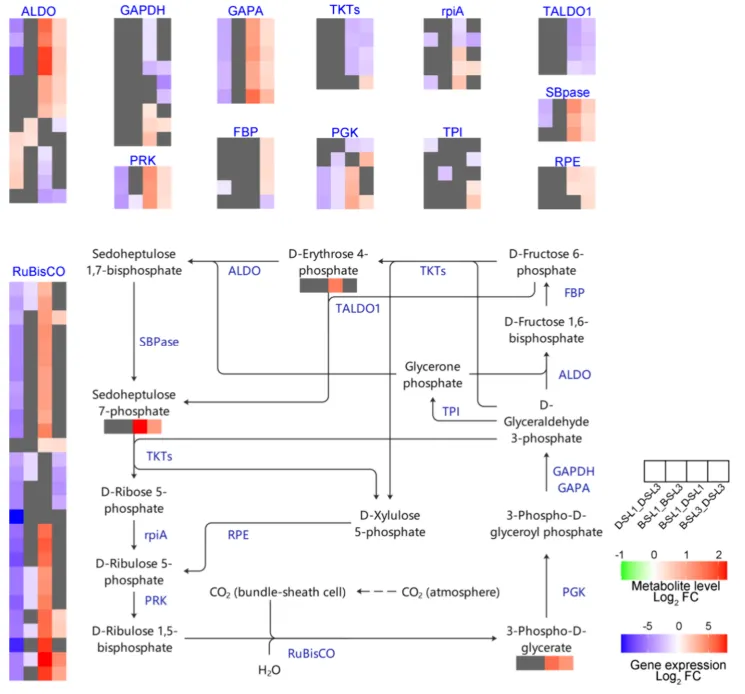
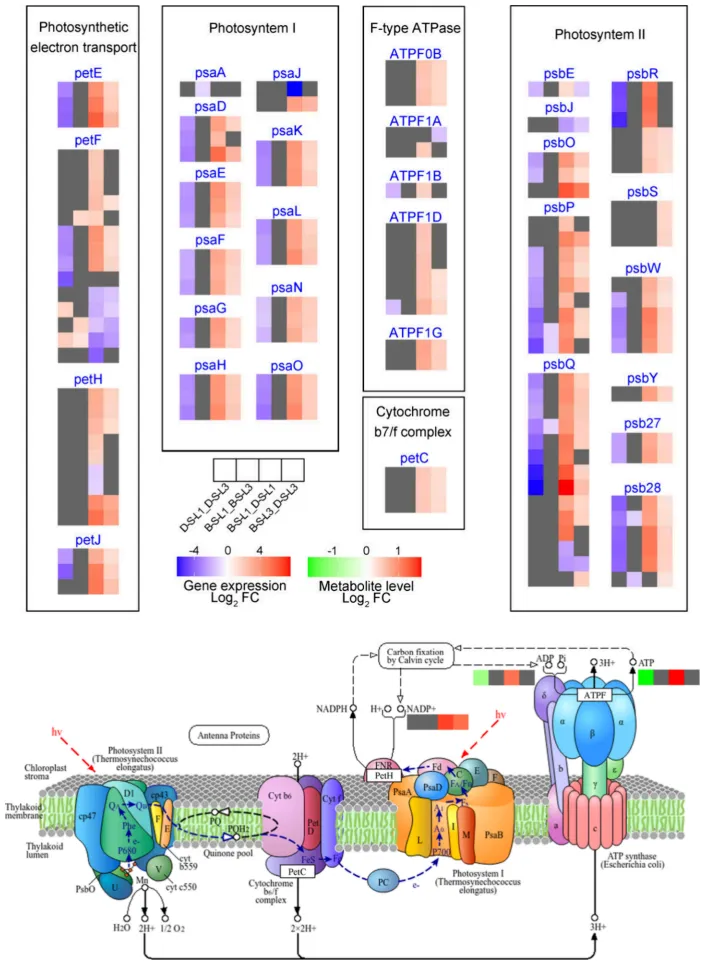
3.4. 盐碱胁迫下燕麦卡尔文循环和光合电子传递的差异揭示了光合损伤的潜在机制
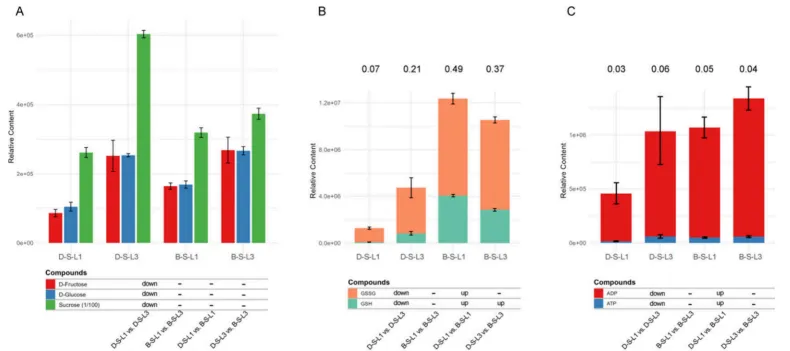
碳固定受损:胁迫下,DY5-L1中关键的RuBisCO家族基因表达量低于DY5-L3和BY14-L1。代谢组学证实,BY14中RuBisCO催化产物3-磷酸-D-甘油酸水平显著高于DY5。DY5-L1的碳固定严重受损,导致葡萄糖、果糖和蔗糖含量显著低于DY5-L3。
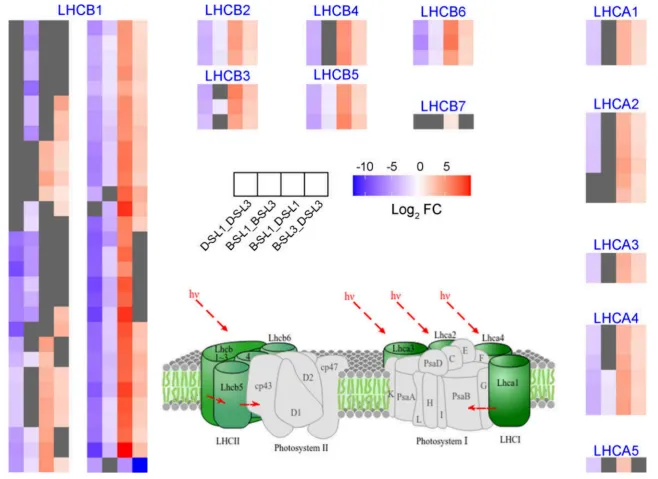
光合调控:光系统I、II及电子传递相关基因在同叶位比较中,BY14的表达水平显著高于DY5,且L1的种间差异大于L3。在品种内,L1的相关基因表达均低于L3,且这种叶位抑制在DY5中更为剧烈。
能量代谢:转录水平的失调导致DY5-L1的能量代谢和电子传递效率下降。代谢组学显示DY5-L1的ATP、ADP和NADP⁺含量显著低于DY5-L3和BY14-L1,证实了其系统性的能量代谢损伤。
转录组与代谢组联合分析表明,卡尔文循环碳固定途径的基因表达和代谢物积累在品种和叶位间存在显著差异。DY5-L1的RuBisCO相关基因表达和中间代谢物含量大幅降低,证实其光合效率下降主要源于非气孔因素导致的碳固定受损。此外,光系统和电子传递途径的基因在DY5-L1中遭遇了更严重的多层次转录抑制,导致ATP和NADP⁺等能量物质匮乏。相比之下,耐受品种BY14维持了更活跃的碳固定和光合功能,并最小化了叶位依赖性的衰退,揭示了DY5下部叶片光合崩溃的系统性机制(Fig.5)(Fig.6)(Fig.7)。
图5. 盐碱胁迫下BY14和DY5燕麦品种L1和L3叶片中卡尔文-本森循环的转录组和代谢组动态。 D-S-L1:DY5-L1;D-S-L3:DY5-L3;B–S-L1:BY14-L1;B–S-L3:BY14-L3。基因符号缩写见表S2。使用DESeq2鉴定差异表达基因(DEGs),阈值为|log₂ FC| ≥ 1且FDR < 0.05。通过OPLS-DA模型确定显著差异代谢物,阈值为变量投影重要性(VIP)> 1且倍数变化≥2或≤0.5。途径模式对应KEGG图00710。
图6. 盐碱胁迫下BY14和DY5燕麦品种L1和L3中光合作用的转录组和代谢组动态。 D-S-L1:DY5-L1;D-S-L3:DY5-L3;B–S-L1:BY14-L1;B–S-L3:BY14-L3。基因符号缩写见表S2。使用DESeq2鉴定DEGs,阈值为|log₂ FC| ≥ 1且FDR < 0.05。通过OPLS-DA模型确定显著差异代谢物,阈值为VIP > 1且倍数变化≥2或≤0.5。途径模式对应KEGG图00195。
图7. 盐碱胁迫下BY14和DY5燕麦品种L1和L3中捕光叶绿素蛋白复合体的转录组和代谢组动态。 D-S-L1:DY5-L1;D-S-L3:DY5-L3;B–S-L1:BY14-L1;B–S-L3:BY14-L3。基因符号缩写见表S2。使用DESeq2鉴定DEGs,阈值为|log₂ FC| ≥ 1且FDR < 0.05。途径模式对应KEGG图00196。
3.5. 盐碱胁迫下BY14与DY5燕麦差异性AsA-GSH循环调控及优越的抗氧化能力
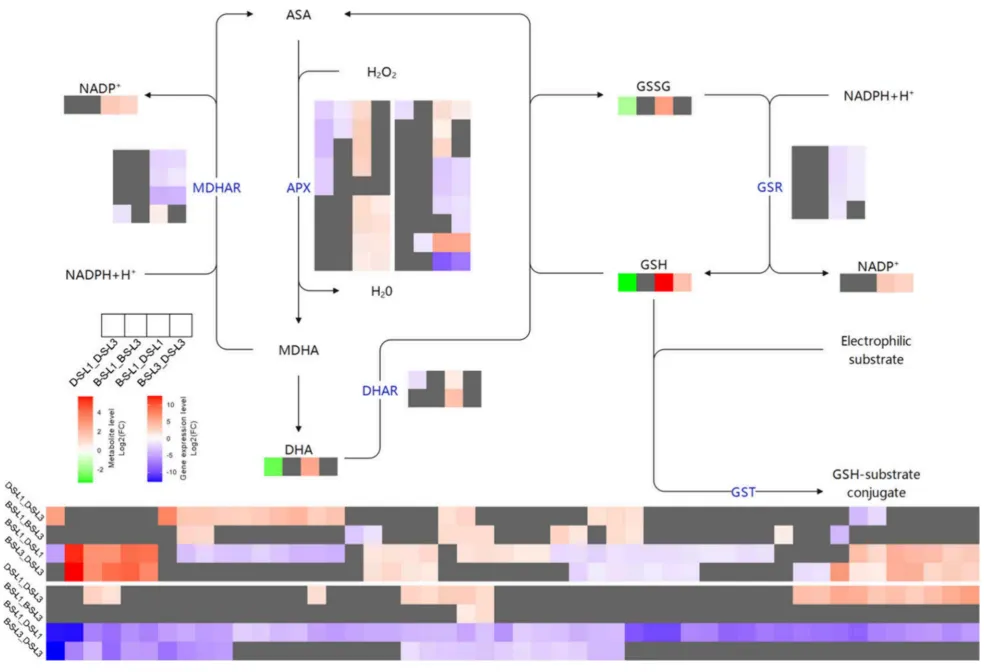
基因表达:与BY14相比,DY5的L1和L3中谷胱甘肽S-转移酶(GST)和谷胱甘肽还原酶(GSR)基因显著上调,表现出代偿性反应。BY14则维持了更高的抗坏血酸过氧化物酶(APX)表达水平。
代谢差异:尽管DY5相关基因上调,其还原型(GSH)和氧化型(GSSG)谷胱甘肽含量却显著低于BY14。BY14保持了显著更高的脱氢抗坏血酸(DHA)含量,表明其抗氧化机制更依赖于自发歧化和GSH依赖性还原途径。
在AsA-GSH循环代谢中,敏感品种DY5虽然表现出GST和GSR基因的代偿性显著上调,但其GSH和GSSG含量却大幅低于耐受品种BY14。BY14在两个叶位均维持了更高的DHA含量和APX基因表达,且较少依赖MDHAR基因,表明其可能更倾向于利用单脱氢抗坏血酸的自发歧化和依赖GSH的还原途径。这些结果证实,尽管DY5启动了转录层面的代偿机制,BY14在整体上仍具备更卓越的抗氧化能力以应对盐碱胁迫(Fig.8)。
图8. 盐碱胁迫下BY14和DY5燕麦品种L1和L3中抗坏血酸-谷胱甘肽(AsA-GSH)循环的转录组和代谢组动态。 D-S-L1:DY5-L1;D-S-L3:DY5-L3;B–S-L1:BY14-L1;B–S-L3:BY14-L3。基因符号缩写见表S2。使用DESeq2鉴定DEGs,阈值为|log₂ FC| ≥ 1且FDR < 0.05。通过OPLS-DA模型确定显著差异代谢物,阈值为VIP > 1且倍数变化≥2或≤0.5。途径模式对应KEGG图00053和00480。
3.6. 盐碱胁迫下DY5与BY14燕麦TCA循环基因的代偿性上调与代谢物失衡
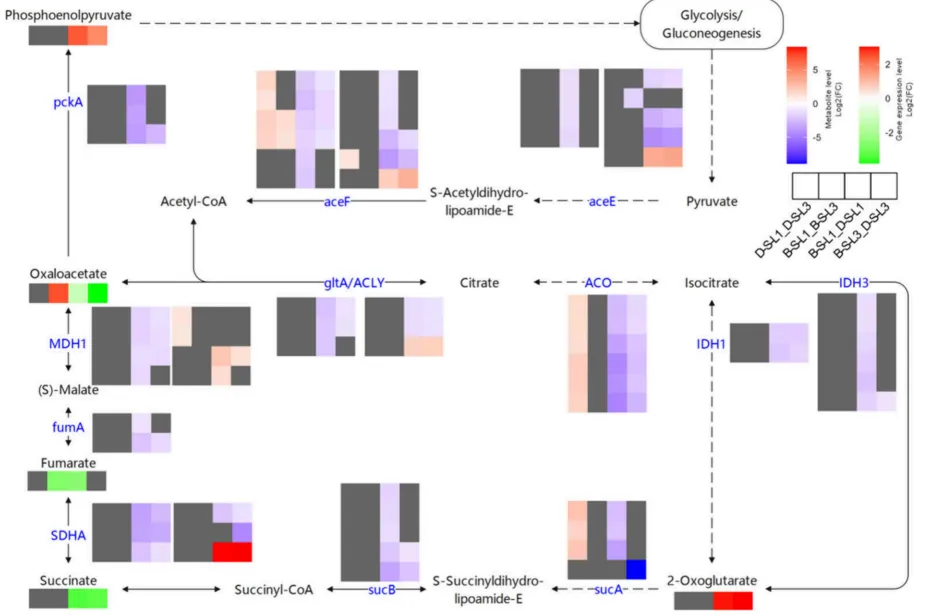
基因代偿:与BY14相比,DY5的TCA循环基因家族呈现整体代偿性上调,且L1的差异幅度显著大于L3。DY5-L3中多个关键酶基因表达一致高于L1。
代谢失衡:DY5中琥珀酸和草酰乙酸水平显著升高,而α-酮戊二酸和磷酸烯醇式丙酮酸(PEP)浓度急剧下降。DY5-L1的延胡索酸含量大幅低于DY5-L3和BY14-L1。
能量亏缺:代谢组学揭示DY5-L1的ATP/ADP比值及相对含量大幅降低,表明能量代谢严重受损。琥珀酸积累和PEP减少暗示相关酶活性受损及糖酵解途径受抑。
TCA循环为生命活动提供核心能量与中间代谢物。在盐碱胁迫下,DY5展现出TCA循环基因的全面代偿性上调,但伴随着严重的代谢流失衡,表现为琥珀酸和草酰乙酸的异常积累以及α-酮戊二酸和PEP的急剧消耗。代谢数据进一步证实DY5-L1面临严重的能量代谢障碍,其ATP/ADP比值大幅下降。这些发现表明,尽管DY5在转录层面进行了代偿性努力,但氧化损伤、代谢流紊乱和能量供应不足共同削弱了其耐盐碱能力,凸显了TCA循环稳态在植物抗逆中的核心作用(Fig.9)(Fig.10)。
图9. 盐碱胁迫下BY14和DY5燕麦品种L1和L3中TCA循环的转录组和代谢组动态。 D-S-L1:DY5-L1;D-S-L3:DY5-L3;B–S-L1:BY14-L1;B–S-L3:BY14-L3。基因符号缩写见表S2。使用DESeq2鉴定DEGs,阈值为|log₂ FC| ≥ 1且FDR < 0.05。通过OPLS-DA模型确定显著差异代谢物,阈值为VIP > 1且倍数变化≥2或≤0.5。途径模式对应KEGG图00020。虚线表示复杂的多步反应。
图10. 代谢组学鉴定的关键代谢物相对含量。 (A)果糖、葡萄糖和蔗糖;(B)GSH和GSSG;(C)ADP和ATP。柱状图上方的字母分别表示GSH/GSSG比值(B)和ATP/ADP比值(C)。柱状图下方出现的显著性标记,指定为“up(上调)”、“down(下调)”或“-(不显著)”,是基于变量投影重要性(VIP)得分大于1结合倍数变化≥2或≤0.5得出的。