当细胞内部的蛋白质折叠工厂——内质网遭遇“生产故障”,未折叠蛋白大量堆积,就会引发一种被称为“内质网应激”的内部危机。长期以来,科学界认为细胞应对这种危机的“求救信号”主要由内质网自身发出,并传递至细胞核,启动相应的保护性基因表达。然而,细胞的最外层屏障——质膜,是否也参与了这场内部危机的感知与应对,一直是个未解之谜。
2026年3月17日,山东农业大学植物保护学院丁新华教授和李洋副教授团队在国际知名期刊《自然-通讯》发表了题为《Magnaporthe oryzae MoPh1 perceives ER stress and promotes adaptive responses via a plasma membrane-to-vacuole pathway》的研究论文。这项研究由尹梓屹、徐佳韵、马世杰作为共同第一作者,揭示了稻瘟病菌中一个全新的、由质膜至液泡的内质网应激响应通路。
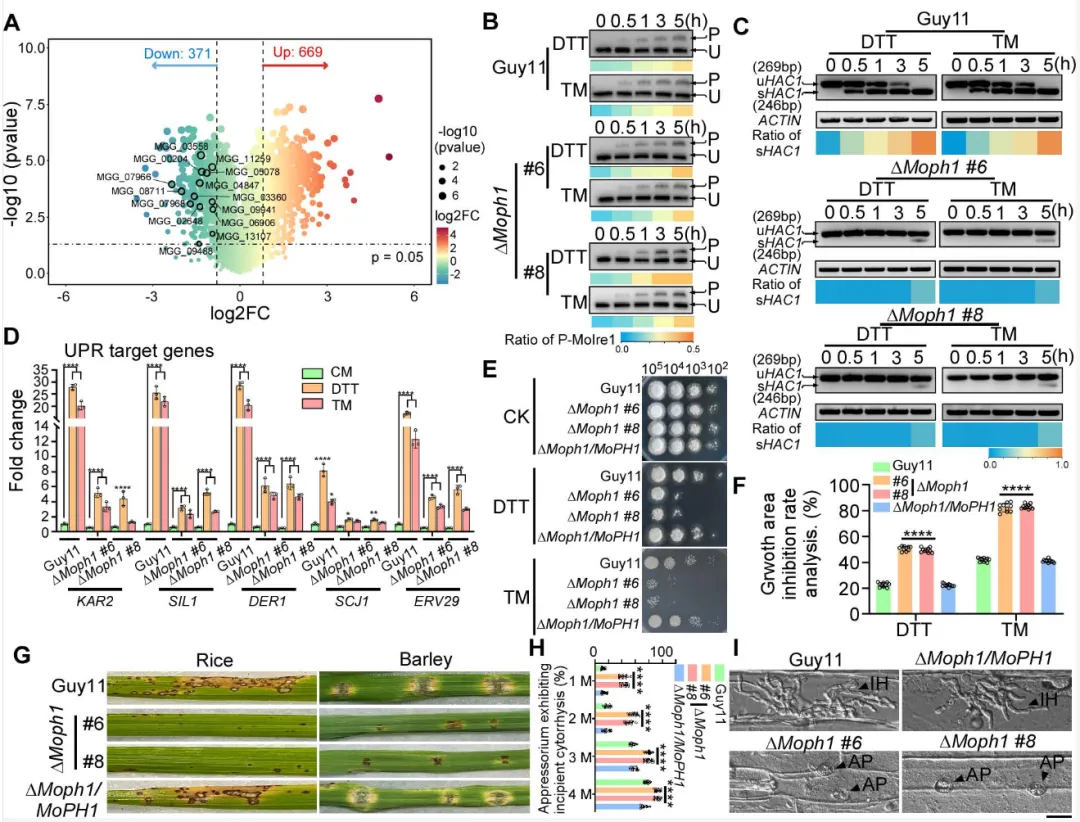
研究人员首先证实,内质网应激的精准调控对稻瘟病菌的存活和致病力至关重要。无论是使用化学药物人为加剧或抑制这种应激,都会导致真菌芽管萌发延迟、附着胞膨压下降,最终使其对水稻的致病力大打折扣,表明病菌需要维持恰到好处的应激水平才能成功侵染。
通过膜蛋白质组学分析,研究团队筛选到一个定位于质膜、含PH结构域的蛋白MoPh1。在正常条件下,MoPh1锚定在质膜上;一旦遭遇内质网应激,它便会内化进入细胞,并最终定位到自噬体和液泡。敲除MoPh1基因后,病菌对内质网应激的耐受性显著下降,致病力也随之减弱,证实了其作为内质网应激“传感器”的关键角色。
进一步研究表明,MoPh1是通过调控自噬过程来帮助细胞应对危机的。在应激状态下,MoPh1能与自噬关键蛋白MoAtg11和MoYpt7发生相互作用,促进自噬体的形成及其与液泡的融合,从而启动细胞内的“清理”程序,降解错误折叠蛋白以缓解压力。电镜观察和蛋白降解实验均证实,缺失MoPh1会导致自噬活动显著减弱。
那么,定位于细胞表面的MoPh1是如何感知来自内部内质网的“求救信号”的呢?研究发现,内质网应激会促使内质网与质膜形成更多的膜接触位点。此时,一个定位于该位点的蛋白MoTcb1能与MoPh1发生特异性结合,将应激信号传递给后者。随后,蛋白激酶MoDbf2对MoPh1进行磷酸化修饰,使其从质膜上释放,启动下游响应。
有趣的是,这条由MoPh1介导的“质膜-液泡”通路,独立于经典的、由内质网膜蛋白MoIre1主导的“内质网-细胞核”通路。但两者并非完全孤立,MoPh1能与MoIre1相互作用,促进后者形成具有高效剪切活性的凝聚体,从而增强经典通路的应激响应效率。
最后,研究还发现这种应激感知模式可能具有物种保守性。在模式植物拟南芥中,MoPh1的同源蛋白AtPH1和AtPH2同样能响应内质网应激发生从质膜到细胞质的转位,且其编码基因的缺失会降低植物对应激的耐受性。
欢迎关注本公众号,所有内容欢迎点赞👍,推荐❤️,评论,转发~
文末点击左下角阅读原文,可跳转文献原文链接~~