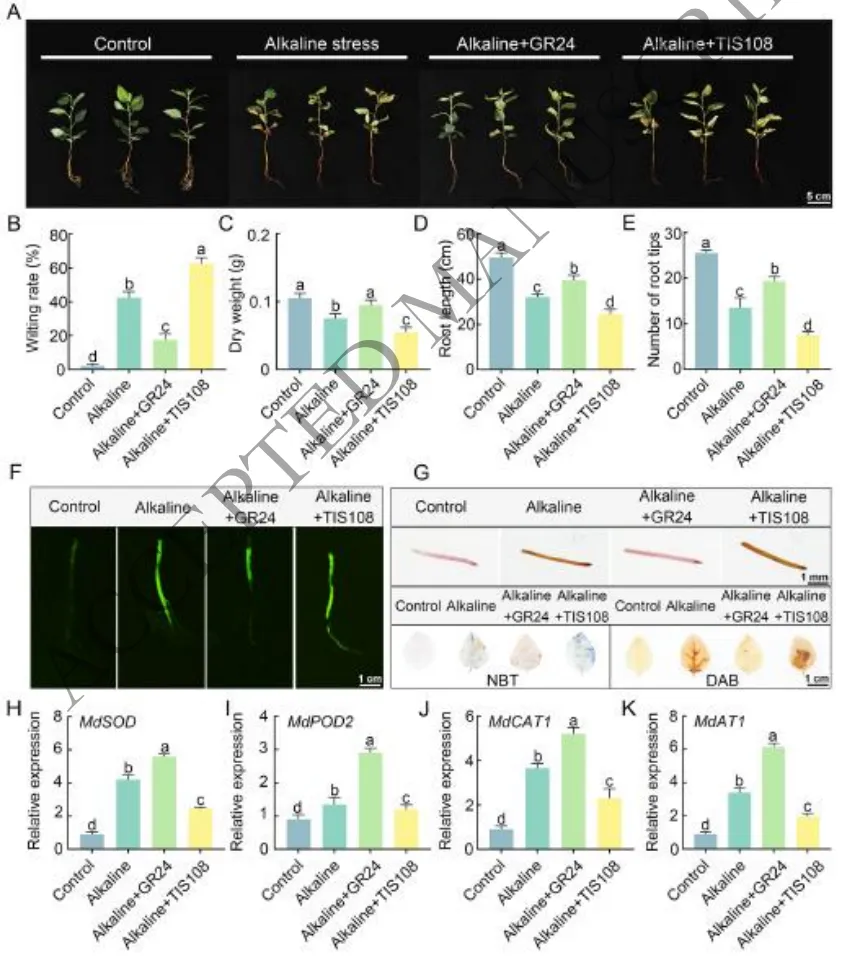
表型改善:外源施用SL类似物GR245DS显著缓解了盐碱胁迫对平邑甜茶幼苗造成的生长抑制、叶片萎蔫和干重下降,而SL合成抑制剂TIS108则加剧了这些损害。
根系保护:盐碱胁迫严重抑制根系伸长和发育,GR245DS处理有效减轻了这种抑制作用,维持了较长的总根长和较多的根尖数,而TIS108处理则进一步恶化了根系生长。
缓解氧化:盐碱胁迫导致根系和叶片中活性氧(ROS)大量积累及膜脂过氧化。GR245DS处理显著降低了H₂O₂和O₂·⁻水平及MDA含量,维持了根际活力。
激活抗氧:GR245DS处理显著提高了SOD、POD和CAT等关键抗氧化酶的表达和活性,并上调了耐碱基因MdAT1的表达,从而通过增强内源抗氧化系统来清除ROS。
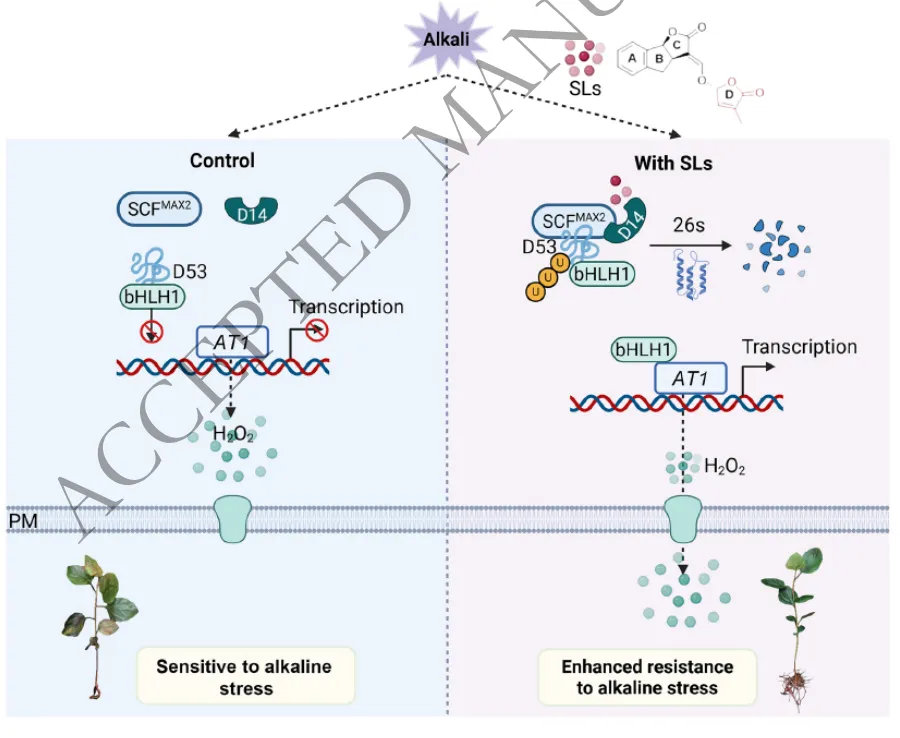
研究表明,外源施用独脚金内酯类似物GR245DS能够有效缓解盐碱胁迫对苹果幼苗造成的生长抑制和根系损伤。在生理层面,GR245DS通过显著提升SOD、POD和CAT等关键抗氧化酶的活性,上调耐碱基因MdAT1的表达,有效清除了体内过量积累的活性氧(H₂O₂和O₂·⁻),降低了膜脂过氧化程度,从而维持了细胞的氧化还原稳态和根际活力。相反,SL合成抑制剂TIS108则加剧了盐碱胁迫引起的氧化损伤和生长抑制。这些结果证实了SL信号通过协同激活内源抗氧化系统介导植物对盐碱胁迫的适应(Fig.1)。
图1. 外源GR245DS和TIS108对盐碱胁迫下平邑甜茶幼苗生长和生理的影响。(A) 对照、盐碱胁迫以及GR245DS或TIS108处理下平邑甜茶幼苗的生长表型;(B) 萎蔫率;(C) 干生物量;(D) 总根长;(E) 根尖数。(F) 根系中ROS荧光信号;(G) 根际活力的TTC染色;叶片ROS检测的NBT和DAB染色;(H-K) 处理下抗氧化酶基因的相对表达水平:(H) MdSOD;(I) MdPOD2;(J) MdCAT1;(K) MdAT1。数据代表三个生物学重复的平均值 ± SD。不同小写字母表示根据Fisher的LSD检验具有显著性差异(P < 0.05)。比例尺:5 cm (A),1 cm (F),1 mm (G)。
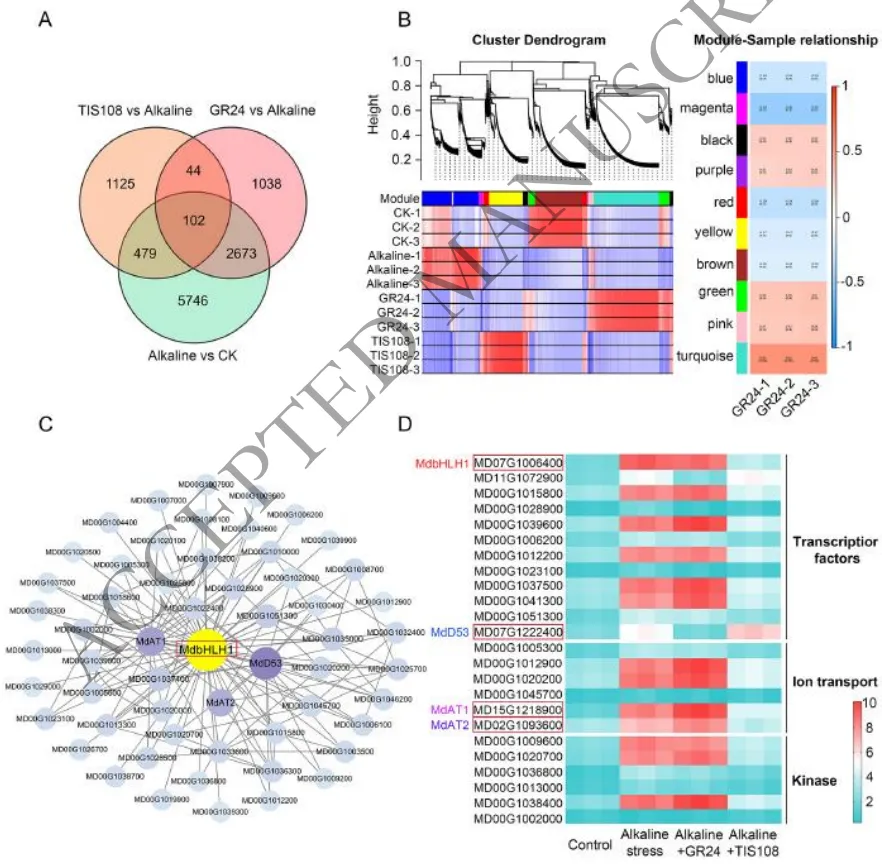
3.2 盐碱胁迫下SL调控核心基因网络的转录组分析
基因筛选:通过对不同处理下的幼苗进行RNA-seq分析,利用韦恩图筛选出102个共同响应盐碱胁迫、GR245DS和TIS108处理的核心差异表达基因(DEGs)。
功能富集:GO和KEGG富集分析表明,这些DEGs主要富集于细胞和代谢过程、膜组分、结合与转运活性,以及植物-病原体互作和植物激素信号传导等通路。
核心鉴定:结合加权基因共表达网络分析(WGCNA),在响应盐碱和GR245DS的模块中鉴定出关键转录因子MdbHLH1,其表达受盐碱胁迫和GR245DS双重强烈诱导,并经RT-qPCR验证为核心响应基因。
为解析SL信号在盐碱胁迫下的调控网络,研究对不同处理的苹果幼苗进行了转录组测序分析。通过差异表达基因筛选与共表达网络分析(WGCNA),鉴定出102个核心响应基因,这些基因主要参与代谢过程、膜转运及激素信号传导等通路。进一步整合分析发现,转录因子MdbHLH1是一个关键的枢纽基因,其表达不仅受到盐碱胁迫的强烈诱导,还能被GR245DS显著上调,而TIS108则抑制其表达。这表明MdbHLH1是SL介导苹果盐碱胁迫响应网络中的核心调控因子(Fig.2)。
图2. 盐碱胁迫和GR245DS处理6小时后平邑甜茶植株的转录组分析。(A) 韦恩图显示盐碱胁迫和盐碱胁迫+GR245DS处理下共响应基因的分布;(B) 基于所有样本拓扑重叠的共表达聚类树状图,显示八个基因模块与不同处理之间的关联。模块-性状相关值范围从-1到1,括号内为P值;(C) 棕色模块中共表达基因的Cytoscape网络互作图;(D) 盐碱响应基因、转录因子和植物激素信号通路中差异表达基因(DEGs)的热图。
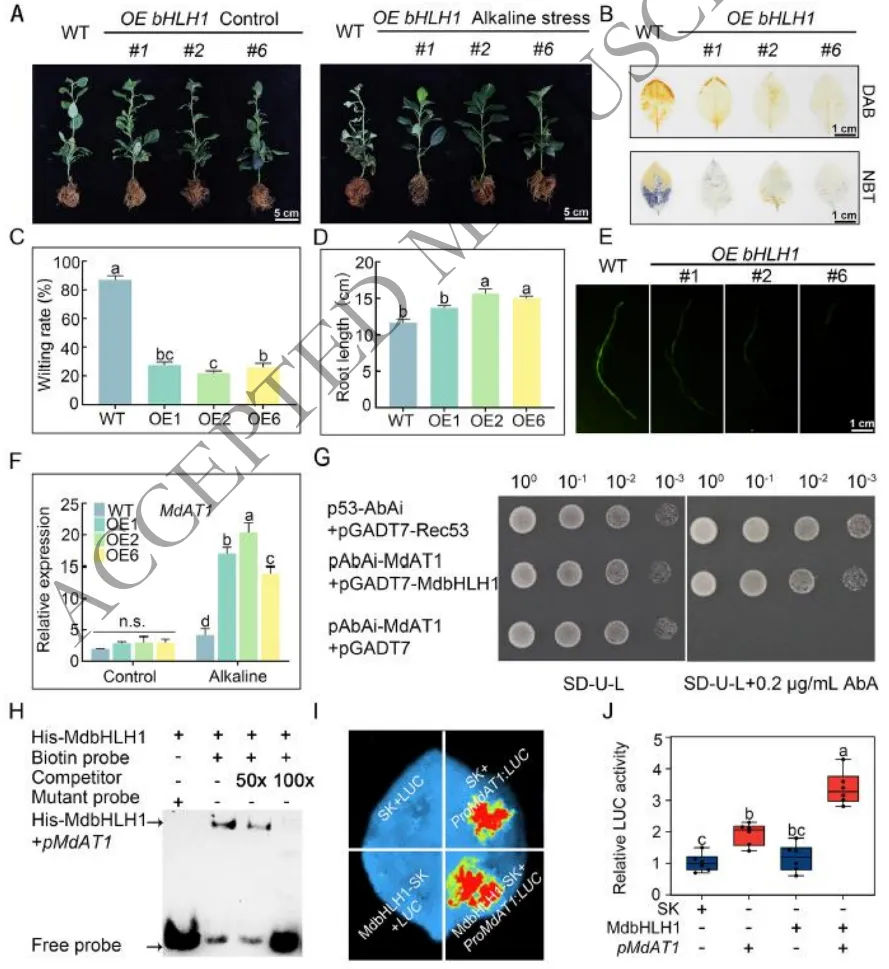
3.3 MdbHLH1通过直接激活MdAT1的表达增强苹果耐盐碱性
表型验证:过表达MdbHLH1的苹果株系在盐碱胁迫下表现出显著改善的生长状态和较低的萎蔫率,而RNAi干扰株系则表现出更严重的受害症状及根系生长抑制。
生理机制:组织化学染色和定量分析显示,过表达MdbHLH1显著减少了叶片和根系中H₂O₂和O₂·⁻的积累及细胞死亡,而RNAi株系中ROS水平大幅升高,证实其能有效缓解氧化损伤。
靶标激活:表达分析、酵母单杂交(Y1H)、凝胶阻滞试验(EMSA)及双荧光素酶报告试验共同证实,MdbHLH1能够直接特异性结合并转录激活下游关键耐碱基因MdAT1的启动子。
为明确MdbHLH1在耐盐碱中的功能,研究构建了其过表达和RNAi干扰苹果株系。表型和生理分析表明,过表达MdbHLH1能够显著降低植株在胁迫下的萎蔫率,大幅减少体内活性氧(H₂O₂和O₂·⁻)的积累并减轻细胞死亡;而敲降该基因则加剧了氧化损伤和生长抑制。分子机制探究进一步揭示,MdbHLH1能够直接结合下游耐碱关键基因MdAT1的启动子区域并激活其转录。这表明MdbHLH1通过直接调控MdAT1的表达来增强ROS清除能力,从而赋予苹果更强的盐碱耐受性(Fig.3)。
图3. MdbHLH1过表达增强耐盐碱性的表型和分子机制。(A) 正常和胁迫条件下WT和OE株系的表型;(B) 叶片的台盼蓝(细胞死亡)和DAB(H₂O₂积累)染色;(C) 盐碱胁迫下的萎蔫率;(D) 根长;(E) 根系中ROS荧光强度;(F) qRT-PCR检测MdAT1的相对表达;(G) Y1H试验显示MdbHLH1结合MdAT1启动子;(H) EMSA证实MdbHLH1特异性结合MdAT1启动子探针;(I-J) 双荧光素酶试验证明MdbHLH1激活MdAT1启动子。数据代表三个生物学重复的平均值 ± SD。不同小写字母表示根据Fisher的LSD检验具有显著性差异(P < 0.05)。比例尺:5 cm (A),1 cm (C/D)。
3.4 MdAT1通过增强ROS清除赋予盐碱耐受性
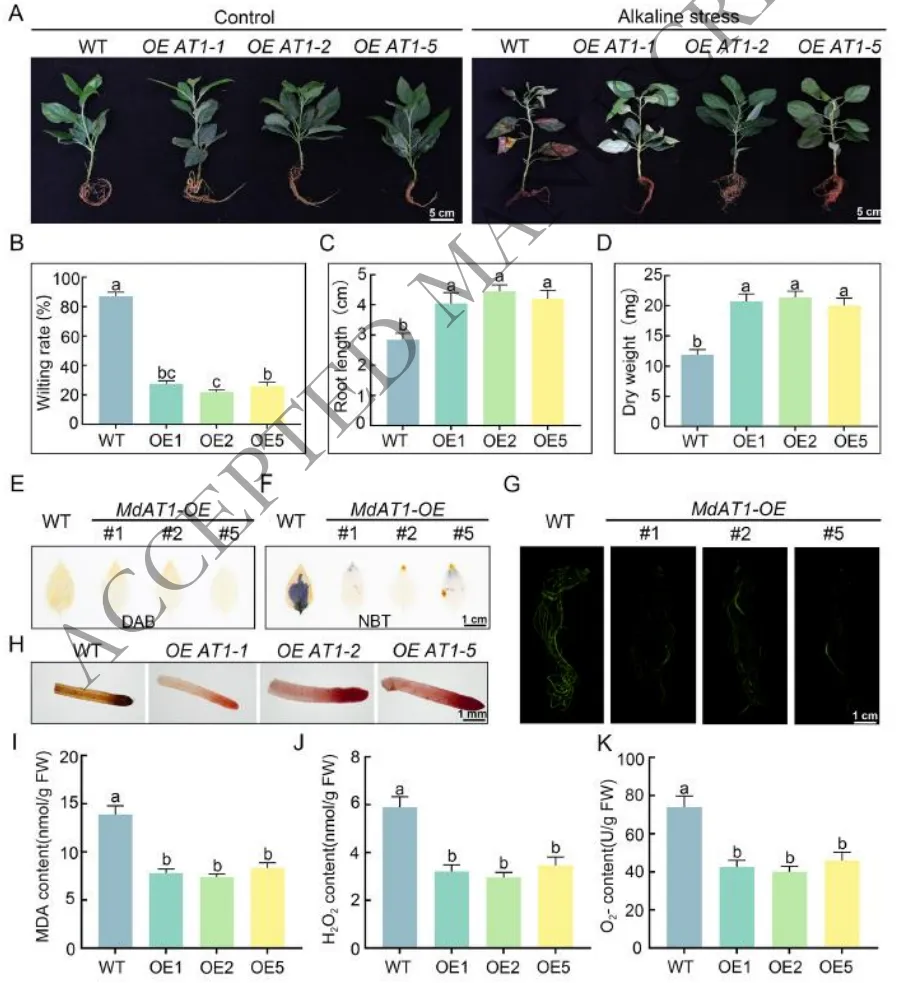
针对下游靶基因MdAT1的功能验证表明,该基因在苹果盐碱胁迫响应中发挥着关键的正向调控作用。在盐碱胁迫下,过表达MdAT1的植株展现出优异的生长状态,其萎蔫率显著降低,根系发育和生物量积累明显优于野生型。生理生化分析证实,MdAT1的过表达大幅降低了细胞内H₂O₂和O₂·⁻的含量以及膜脂过氧化水平,有效维持了根际活力。相反,MdAT1表达受抑的植株则遭受了更严重的氧化损伤。这些结果确证了MdAT1通过高效调控ROS清除机制来减轻氧化损伤,从而赋予植物盐碱耐受性(Fig.4)。
图4. MdAT1过表达增强耐碱性的表型和生理机制。(A) 对照(左)和胁迫(右)条件下WT和OE株系的表型;(B) 萎蔫率;(C) 干生物量;(D) 根长;(E) 检测H₂O₂积累的DAB染色;(F) 检测O₂·⁻积累的NBT染色;(G) 根系中ROS荧光信号;(H) 根际活力的TTC染色;(I) MDA含量的定量分析。(J) H₂O₂含量的定量分析;(K) O₂·⁻含量的定量分析。数据代表三个生物学重复的平均值 ± SD。不同小写字母表示根据Fisher的LSD检验具有显著性差异(P < 0.05)。比例尺:5 cm (A),1 cm (E/G),1 mm (H)。
3.5 MdD53作为SL信号的负调控因子拮抗MdbHLH1的功能
蛋白互作:通过CoIP-MS筛选及转录组数据分析锁定MdD53,随后利用酵母双杂交(Y2H)、双分子荧光互补(BiFC)和双荧光素酶等试验证实了MdD53与MdbHLH1之间存在直接的蛋白相互作用。
降解受控:免疫印迹分析表明,盐碱胁迫诱导MdD53蛋白积累,而外源SL处理促进其降解,SL合成抑制剂TIS108则阻断了这一降解过程。
负向调控:表型和生理分析显示,MdD53过表达株系在盐碱胁迫下表现出更严重的坏死和极高的ROS积累,而RNAi株系则表现出明显的抗性提升和ROS水平下降,证明其为耐盐碱的负调控因子。
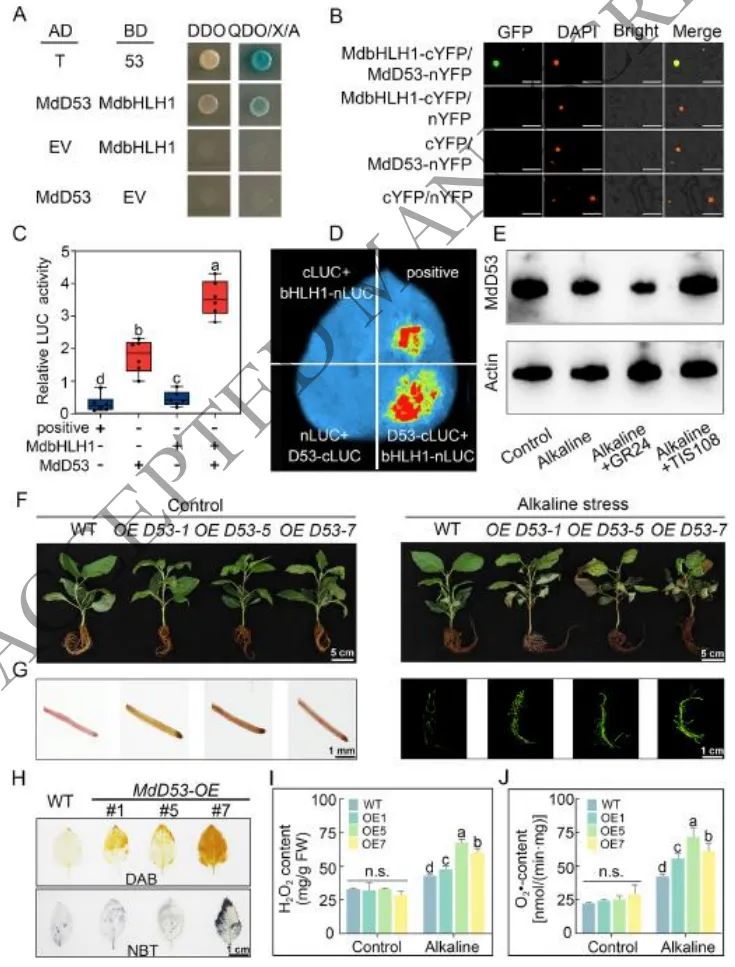
进一步的研究揭示了SL信号通路核心抑制因子MdD53在盐碱胁迫中的作用机制。多种分子互作试验(Y2H、BiFC、LCI等)一致证实,MdD53能够与转录因子MdbHLH1发生直接的物理相互作用。蛋白质水平分析发现,MdD53的丰度受SL信号的严格调控,外源SL可诱导其降解。在功能层面,过表达MdD53会导致苹果植株在盐碱胁迫下积累大量活性氧,加剧叶片坏死和生长抑制;而敲降MdD53则显著提升了植株的ROS清除能力和耐盐碱性。这表明MdD53通过与MdbHLH1互作并削弱ROS清除能力,在苹果耐盐碱胁迫中发挥负向调控作用(Fig.5)。
图5. MdD53与MdbHLH1互作调控苹果耐盐碱性的分子机制。(A) Y2H验证MdD53与MdbHLH1之间的互作;(B) BiFC在烟草表皮细胞中证实MdD53-MdbHLH1互作;(C) 双荧光素酶报告系统定量测量互作强度;(D) 使用荧光素酶互补成像可视化蛋白互作;(E) 盐碱胁迫和SL处理下MdD53蛋白积累的蛋白质印迹分析;(F) 正常和胁迫条件下WT和OE株系的表型;(G) 根系中ROS荧光信号检测;(H) 叶片ROS检测的DAB和NBT染色;(I) H₂O₂含量的定量分析;(J) O₂·⁻含量的定量分析。数据代表三个生物学重复的平均值 ± SD。不同小写字母表示根据Fisher的LSD检验具有显著性差异(P < 0.05)。比例尺:5 cm (F),1 cm (H),1 mm (G)。
3.6 MdD53-MdbHLH1-MdAT1模块协同调控氧化胁迫响应
转录抑制:EMSA和双荧光素酶报告试验表明,MdD53的加入显著削弱了MdbHLH1对MdAT1启动子的结合能力及其转录激活作用,证实MdD53抑制了该调控途径。
遗传互补:瞬时转化试验显示,MdAT1过表达能部分逆转由MdD53过表达引起的生长抑制,证明MdbHLH1-MdAT1通路能够拮抗MdD53介导的负向效应。
生理协同:氧化应激指标与基因表达高度一致,MdD53过表达或MdbHLH1敲降导致ROS激增及MdAT1表达下降,而共表达MdAT1有效降低了由MdD53引起的ROS积累。
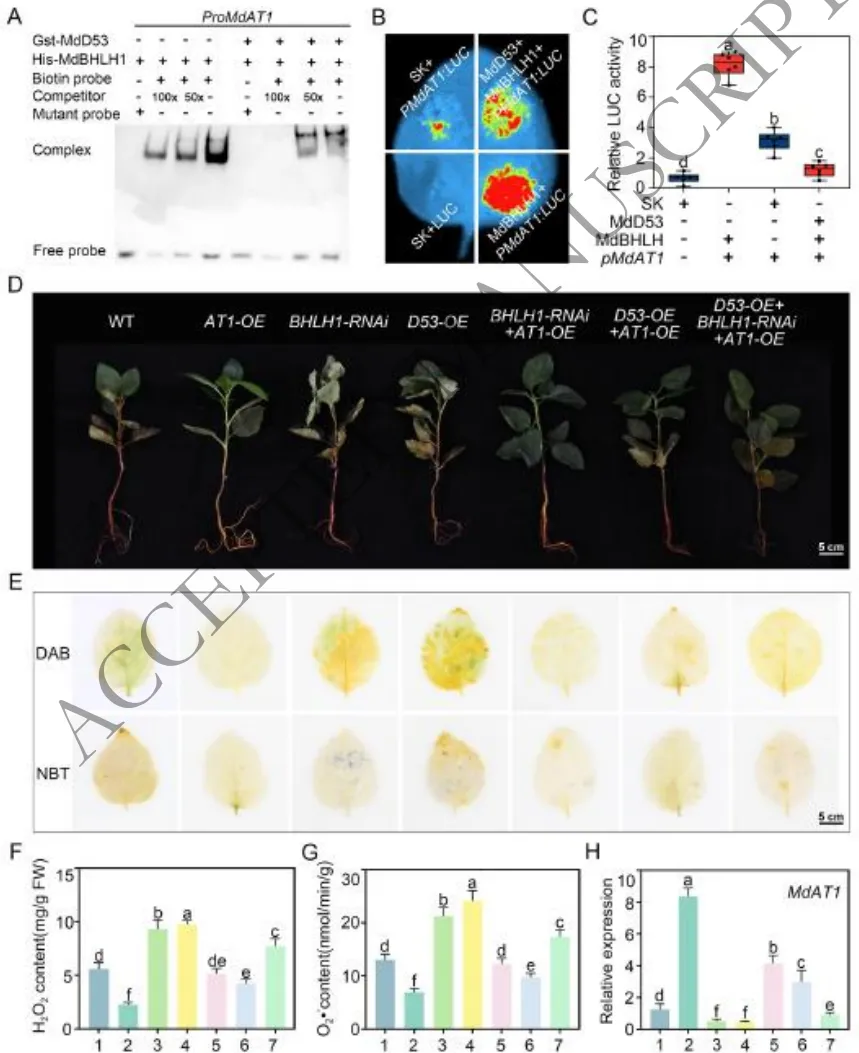
为系统阐明该调控模块的协同作用机制,研究从分子互作、基因互补和生理响应三个维度进行了深入探讨。体外和体内分子试验证实,MdD53通过与MdbHLH1结合,直接阻碍了MdbHLH1对下游靶基因MdAT1启动子的识别与转录激活。遗传互补与生理分析表明,过表达MdD53或敲降MdbHLH1均会抑制MdAT1的表达,导致植株体内H₂O₂和O₂·⁻大量积累并引发严重的生长抑制;而同时过表达MdAT1能够有效逆转MdD53过表达所造成的ROS激增和表型受害。这些结果完整勾勒出一条清晰的调控链条:MdD53通过抑制MdbHLH1的转录活性来下调MdAT1的表达,最终削弱了植物的ROS代谢与盐碱耐受能力(Fig.6)。
图6. MdD53-MdbHLH1-MdAT1调控模块在苹果耐碱性中的分子机制。(A) MdD53对MdbHLH1结合MdAT1启动子影响的EMSA分析;(B-C) 双荧光素酶报告试验检测MdD53对MdbHLH1介导的MdAT1激活的抑制作用;(D) 盐碱胁迫下通过瞬时过表达的不同遗传材料的表型比较;(E) 用于ROS检测的DAB和NBT染色;(F) H₂O₂含量的定量分析;(G) O₂·⁻含量的定量分析;(H) MdAT1基因的表达水平分析。数据代表三个生物学重复的平均值 ± SD。不同小写字母表示根据Fisher的LSD检验具有显著性差异(P < 0.05)。比例尺:5cm (D/E)。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
