3.1 基因组组装与注释
基因组组装:利用HiFi和Hi-C数据实现了染色体级别的组装,获得了极高的连续性和准确性。
基因组注释:结合从头预测与转录组证据,对蛋白编码基因、非编码RNA及重复序列进行了全面注释。
结构完整性:通过端粒和着丝粒的精确鉴定,进一步验证了染色体级别组装的完整性与准确性。
本研究利用多组学数据成功构建了高质量的染色体级别黄花苜蓿(M. falcata)二倍体参考基因组。组装结果展现出极高的连续性与准确性,绝大部分序列被精确定向至8条染色体上。通过综合策略完成了详尽的基因组注释,并成功鉴定出端粒与着丝粒结构,充分证实了该基因组组装的完整性与可靠性。
3.2 比较基因组学分析
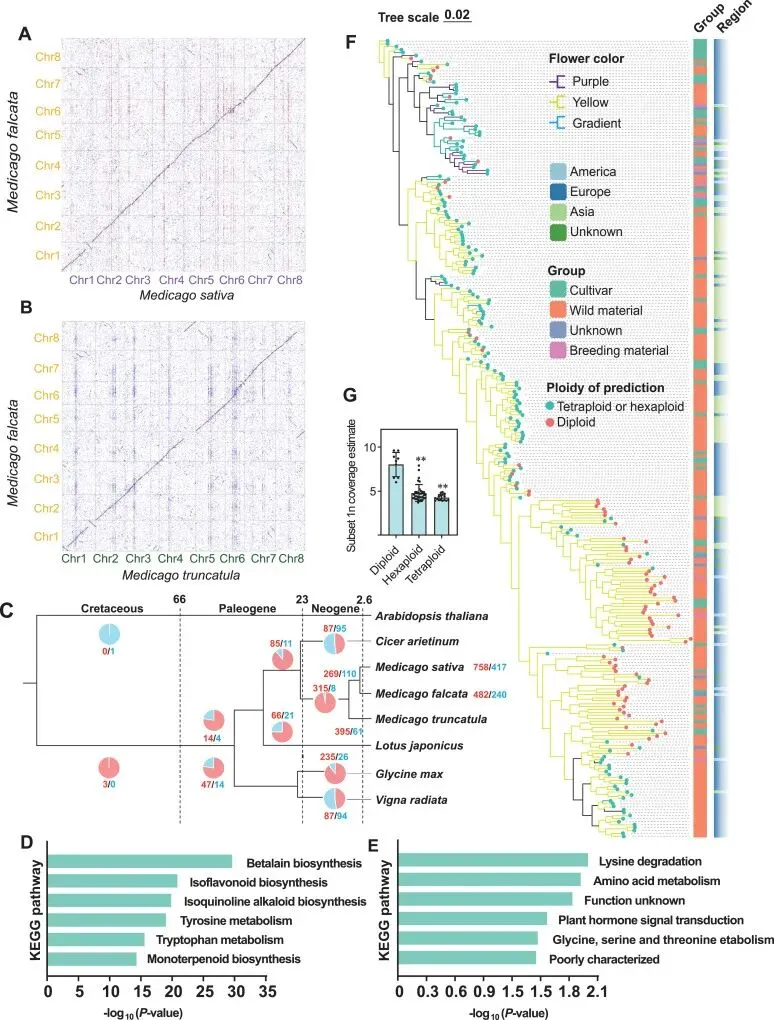
通过与紫花苜蓿(M. sativa)及蒺藜苜蓿(M. truncatula)等近缘物种的比较基因组学分析,证实了黄花苜蓿基因组在整体上保持了高度的共线性。同时,研究也揭示了若干局部的显著结构变异,如1号染色体的倒位及特定染色体末端的易位事件。这些结构层面的差异可能在染色质组织、表观遗传调控及基因表达中发挥重要作用(Fig. 1)。
3.3 系统发育与进化分析
系统发育:明确了黄花苜蓿与紫花苜蓿的最近亲缘关系及分化时间节点。
基因家族:发现抗氧化、抗逆及花色代谢相关通路的基因家族在黄花苜蓿中发生显著扩张。
正选择分析:鉴定出受正选择作用的基因,这些基因主要富集于光合作用及激素信号传导等途径。
基于保守单拷贝基因的分析明确了黄花苜蓿与紫花苜蓿的进化关系及分化节点。基因家族进化分析揭示了黄花苜蓿在次生代谢、抗氧化防御及非生物胁迫响应相关通路中的基因家族显著扩张,这为其耐寒、抗旱及特定花色等适应性表型提供了遗传基础。此外,正选择分析进一步锁定了与光合作用和激素信号传导相关的关键适应性进化基因,为抗逆苜蓿品种的培育提供了潜在靶点(Fig. 1)。
3.4 群体基因组学与倍性鉴定
群体结构:揭示了黄花苜蓿显著的地理结构分化以及与花色高度相关的遗传分化模式。
倍性鉴定:结合多种测序分析与流式细胞术,精准区分并鉴定了群体中的二倍体与多倍体种质。
区域分化:发现欧亚群体在光合作用相关途径上存在显著遗传差异,反映了对不同环境压力的适应。
通过对大规模种质资源的重测序分析,深刻揭示了黄花苜蓿群体在宏观地理尺度上的显著结构分化,并发现花色表型与遗传背景之间存在紧密联系及潜在的基因交流。结合多维度数据与流式细胞术,成功实现了对复杂群体倍性组成的精准鉴定(t检验P值分别为5.07 × 10⁻⁵和1.35 × 10⁻⁵)。进一步的区域分化研究表明,欧洲与亚洲群体在光合作用相关基因上的显著差异,深刻反映了其对各自独特气候与环境压力的适应性进化(Fig. 1)。
图1. 黄花苜蓿的基因组组装与比较基因组学分析。 A:黄花苜蓿(黄色字体)与紫花苜蓿(紫色字体)之间的点阵图共线性比较。B:黄花苜蓿与蒺藜苜蓿(绿色字体)之间的点阵图共线性比较。C:黄花苜蓿及近缘物种的时间校准系统发育树。基于二倍体黄花苜蓿基因组与四倍体紫花苜蓿单一亚基因组的比较,在每个分支上标示了基因家族的扩张(红色)和收缩(蓝色)。这些扩张与收缩为了解塑造黄花苜蓿基因组的进化过程提供了见解。D:黄花苜蓿中扩张基因家族的KEGG通路富集分析。E:黄花苜蓿与紫花苜蓿之间Ka/Ks ≥ 2的基因的KEGG通路富集分析。F:黄花苜蓿种质的系统发育树,附带推断的倍性水平、花色、栽培种分类及地理来源。G:条形图展示了通过流式细胞术确认倍性的样本在k-mer分析中得出的“Subset 1n Coverage Estimate”值。
3.5 研究结论
本研究成功构建了黄花苜蓿染色体级别的高质量二倍体基因组组装。不仅揭示了黄花苜蓿、紫花苜蓿与蒺藜苜蓿之间的结构与功能差异,还鉴定了抗逆相关基因家族的扩张及正选择信号。此外,通过对来自不同地理区域的271份种质资源进行重测序,阐明了其遗传聚类模式、倍性组成以及花色与遗传背景之间的复杂关系。总而言之,该研究为推进黄花苜蓿的进化与遗传学研究提供了宝贵的基因组资源和理论框架,同时也为苜蓿属植物的抗逆育种、品种改良及草原生态修复奠定了坚实基础。
填补二倍体参考基因组空白,实现高质量染色体级组装。长期以来,黄花苜蓿(M. falcata)的基因组研究受限于其复杂的倍性变异,此前仅有连续性较低的四倍体基因组(Contig N50仅为5.67 Mb)。本研究首次采用具有罕见直立生长习性和耐盐碱特性的二倍体种质(PI631549),综合利用PacBio HiFi、Hi-C、Illumina和转录组等多组学数据,成功组装了迄今为止最高质量的染色体级别二倍体参考基因组(Contig N50大幅提升至47.39 Mb,99.99%的序列被精确定向,并成功鉴定出端粒和着丝粒),为苜蓿属植物的遗传学研究提供了结构简化的、高质量的底层数据支撑。
揭示黄花苜蓿独特抗逆性与表型特征的进化遗传基础。研究首次从全基因组层面解析了黄花苜蓿区别于紫花苜蓿(M. sativa)的独特表型(如黄花、极强的耐寒和抗旱能力)的遗传基础。创新性地发现其在甜菜红素(betalain)、异黄酮(isoflavonoid)和异喹啉生物碱(isoquinoline alkaloid)等次生代谢途径中的基因家族发生显著扩张。这些扩张不仅解释了其花色决定的机制,更揭示了其通过增强渗透调节和抗氧化活性来应对非生物胁迫的适应性进化策略。
精准鉴定近缘物种间的大规模复杂结构变异(SVs)。通过与紫花苜蓿和蒺藜苜蓿(M. truncatula)进行高精度的比较基因组学和长程序列比对分析,本研究不仅证实了物种间广泛的共线性,还精准定位了特定的局部结构变异。例如,首次明确揭示了黄花苜蓿在1号染色体远端和近端的特异性倒位,以及在4号和8号染色体末端的明显易位事件,为理解染色质组织、表观遗传调控和基因表达的结构性驱动因素提供了新见解。
创新多维度的倍性鉴定策略,全面解析复杂群体遗传结构。面对黄花苜蓿种质资源中复杂的倍性变异(二倍体、四倍体、六倍体混杂)且部分样本测序深度有限的挑战,本研究创新性地提出了一种整合策略:将基于测序数据的k-mer分析(Jellyfish)、SNP组成分析(Smudgeplot)与传统的流式细胞术(以“Subset 1n Coverage Estimate”为核心指标)深度结合。这一多维度交叉验证的方法成功且精准地鉴定了271份重测序种质的倍性组成,为多倍体植物群体的倍性鉴定提供了方法学参考。
阐明地理环境驱动的区域适应性进化机制。研究不仅在宏观地理尺度上揭示了欧洲、亚洲和美洲群体间显著的系统发育分离,还深入挖掘了环境压力驱动的区域分化机制。创新性地发现欧洲群体(适应寒冷、短生长季)与亚洲群体(适应干旱)在光合作用相关途径的基因上存在显著的等位基因频率差异(包含457个非同义突变)。这一发现深刻揭示了不同生态位环境压力如何塑造黄花苜蓿光合作用效率的适应性分化。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
