2.1 试验材料制备与基因鉴定分析
为探究高三叶中目标基因的表达特征与功能,研究首先对TaUPL基因家族进行了系统的鉴定与生物信息学分析。在基因表达水平的测定上,提取了植物组织的总RNA,并针对目标基因设计特异性引物,执行了实时荧光定量PCR(qRT-PCR)检测。在遗传转化材料的准备方面,从试验田采集返青的根茎芽,经温室驯化7天后用于·的转化;同时,将高三叶种子经75%乙醇消毒、无菌水冲洗及4℃春化48小时后,接种于1/2 MS培养基上促萌发,为后续构建转基因毛状根体系提供了标准化的植物材料。
2.2 遗传转化实施与DNA亲和纯化测序(DAP-Seq)
为验证目标基因在根系发育中的作用及其潜在的DNA结合活性,研究实施了毛状根遗传转化与高通量测序分析。在转化处理中,切取生根幼苗的子叶与胚根交界处,将其上部浸入含有pCAMBIA1300-TaAGL29重组质粒的K599农杆菌菌液中,随后转移至MS培养基以诱导不定根的生成。为了在全基因组水平筛选转录调控靶标,采用体外无细胞表达系统诱导合成TaUPL21重组蛋白,并提取高三叶基因组DNA构建测序文库。将重组TaUPL21蛋白与基因组DNA文库共孵育后,分离出与该蛋白特异性结合的DNA片段,最终通过高通量测序技术精准鉴定了其结合位点。
2.3 靶向互作验证与数据统计分析
为进一步证实TaUPL21蛋白与TaAGL29启动子之间的直接结合及转录激活关系,研究设计并执行了酵母单杂交(Y1H)与双荧光素酶(Dual-Luc)报告基因试验,并辅以严格的统计学检验。Y1H试验中,将分别搭载TaUPL21和TaAGL29启动子的pGADT7与pHIS2重组载体转入Y187酵母细胞,并在SD/-His/-Leu及SD/-His/-Leu/-Trp缺陷型培养基上进行阳性克隆筛选。在双荧光素酶试验中,将相关重组载体转入GV3101农杆菌并注射至五叶期烟草叶片,48小时后利用活体成像仪观察荧光信号,随后裂解叶片组织并分别测定萤火虫荧光素酶(LUC)与海肾荧光素酶(REN)的活性比值。所有测定均包含三个独立的生物学重复,数据采用SPSS软件进行方差分析(ANOVA),并通过Duncan多重比较法检验各处理间的差异,统计学显著性水平设定为p < 0.05。
3.1 高三叶UPL基因家族的鉴定与理化性质
全基因鉴定:从高三叶转录组数据中成功鉴定出32个TaUPL基因。
理化性质:编码蛋白的氨基酸长度与分子量差异较大,多数等电点偏酸性,呈亲水性且多具核定位信号。
分子对接:预测结合能结果显示,TaUPL21蛋白表现出最强的结合亲和力。
基于高三叶转录组数据,共鉴定出32个TaUPL基因(TaUPL1至TaUPL32)。理化性质分析表明,这些基因编码的蛋白在氨基酸长度和分子量上存在较大差异,且绝大多数(94%)的理论等电点低于7,提示其主要在弱酸性微环境中发挥功能。所有TaUPL蛋白的亲水性总平均值均为负值,证实了其亲水性特征,且超过七成的成员具有假定的核定位信号。此外,分子对接结果显示,TaUPL家族的预测结合能呈现不同程度的亲和力,其中TaUPL21表现出最强的预测结合能力。
3.2 系统发育与组织表达水平分析
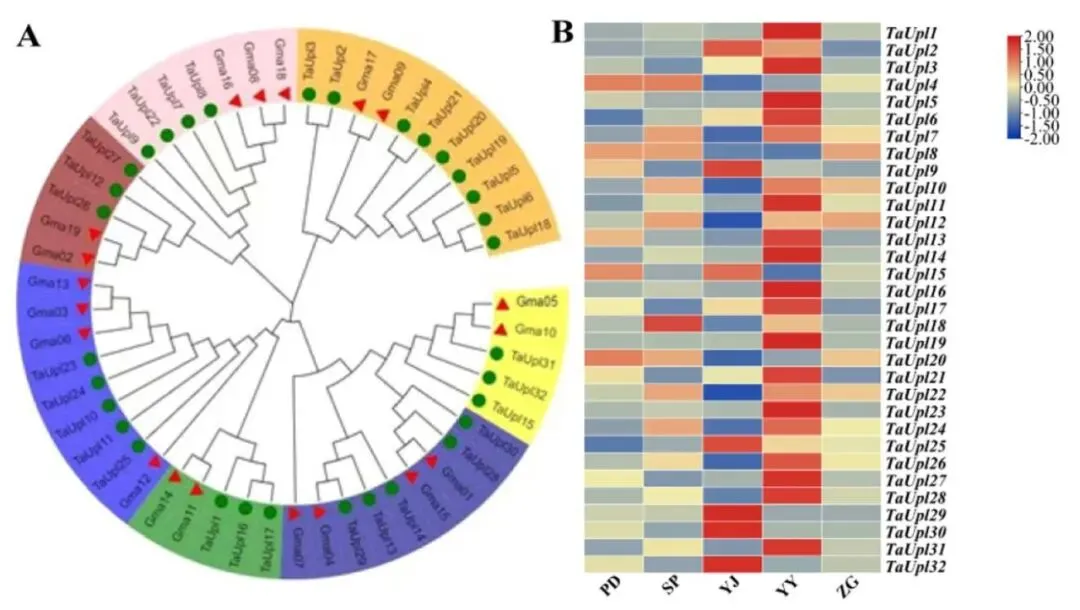
利用大豆UPL蛋白序列构建的最大似然系统发育树将UPL基因划分为七个不同的进化分支(Class I–VII)。这种跨物种的分布模式表明TaUPL基因在植物进化过程中保持了高度的功能保守性。进一步结合已发表的转录组数据对不同根部组织的表达谱进行分析,发现32个TaUPL基因中有25个在根茎芽中的表达量显著高于其他四种根部组织(Fig.1)。
图1.大豆与高三叶TaUPL基因家族的最大似然(ML)系统发育树及组织表达谱 (A) 使用全长序列构建的系统发育树(Bootstrap值为1000)。红色三角形和绿色圆形分别代表大豆和高三叶。不同分支代表不同的UPL亚家族。(B) TaUPL家族在高三叶不同根部组织的表达水平。PD:主根膨大部,SP:水平根,YJ:根茎芽尖;YY:根茎芽;ZG:主根。
3.3 TaUPL蛋白结构域与基序分析
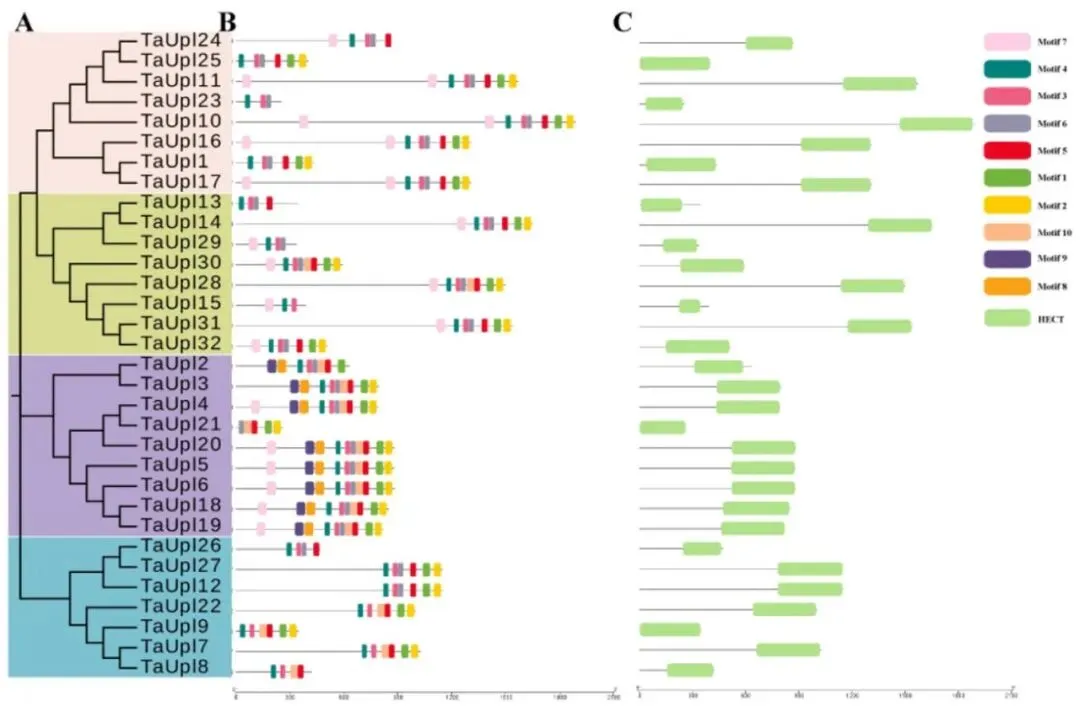
为探究功能结构域和进化关系,单独为TaUPL蛋白构建了最大似然系统发育树,将其聚类为四个不同的进化簇。保守结构域分析证实,所有32个TaUPL蛋白均包含特征性的HECT结构域,且同一进化簇内的蛋白具有相似的结构域组成。利用MEME进行的基序分析进一步识别出多个保守基序,同一系统发育簇内的成员包含完全相同的基序组成。这些分析共同证实了TaUPL基因家族在结构和进化上的高度保守性(Fig.2)。
图2. TaUPL的系统发育关系、保守基序和基因结构域 (A) 使用全长序列构建的TaUPL蛋白最大似然(ML)系统发育树(Bootstrap值为1000)。(B) TaUPL的结构域分布。(C) TaUPL蛋白中保守基序的分布。共预测了10个基序,比例尺代表100个氨基酸。
3.4 TaUPL21的生物信息学特征与无细胞表达
进化与序列:系统发育与多序列比对揭示了TaUPL21在多物种间的高度进化保守性。
结构与表达:三维结构呈现典型特征,利用无细胞系统成功表达了该亲水性蛋白。
互作网预测:预测网络包含泛素前体和蛋白酶体亚基,支持其E3泛素连接酶的经典功能。
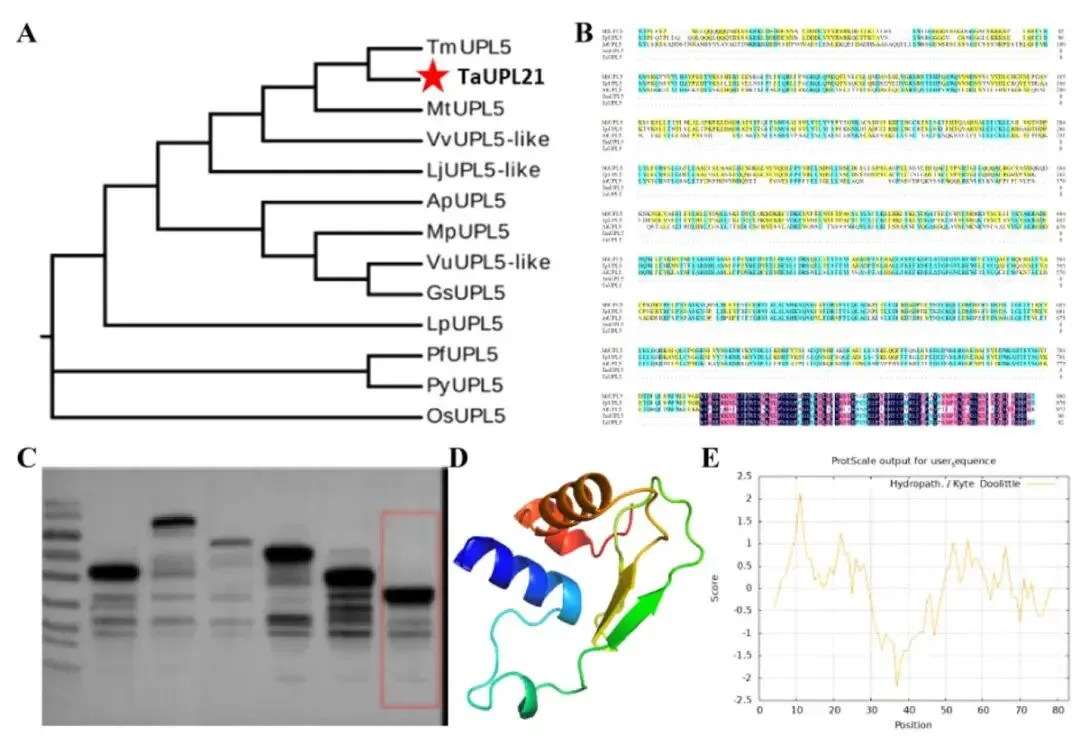
通过一系列生物信息学分析揭示了TaUPL21的进化和结构特征。系统发育分析显示TaUPL21与其在红三叶中的直系同源物紧密聚类,多序列比对进一步揭示了其在多个物种间的保守区域。负的亲水性总平均值证实了TaUPL21的亲水性质,其三维结构预测也展现出典型的结构域构型。随后,利用小麦胚芽无细胞表达系统成功合成了TaUPL21蛋白,SDS-PAGE分析在预测分子量处确认了单一目的条带。利用STRING数据库预测的潜在互作蛋白网络中包含了泛素前体和蛋白酶体亚基,这与其作为HECT型E3泛素连接酶的分类相一致。结合保守HECT结构域的存在,这些计算机模拟分析表明TaUPL21除了具有新发现的DNA结合功能外,很可能保留了经典的泛素连接酶活性(Fig.3)。
图3. TaUPL21的系统发育、序列比对及结构分析 (A) 使用全长序列构建的TaUPL21最大似然(ML)系统发育树(Bootstrap值为400)。Ap:相思子;Gs:野大豆;Lj:百脉根;Lp:宿根亚麻;Mp:刺毛黧豆;Mt:截形苜蓿;Os:水稻;Pf:紫苏;Py:染井吉野樱;Tm:红三叶;Vu:赤豆;Vv:长柔毛野豌豆。(B) TaUPL21的氨基酸多序列比对。(C) 蛋白表达结果。(D) TaUPL21蛋白的三维结构。(E) 疏水性分析。
3.5 TaUPL21促进高三叶根系生长
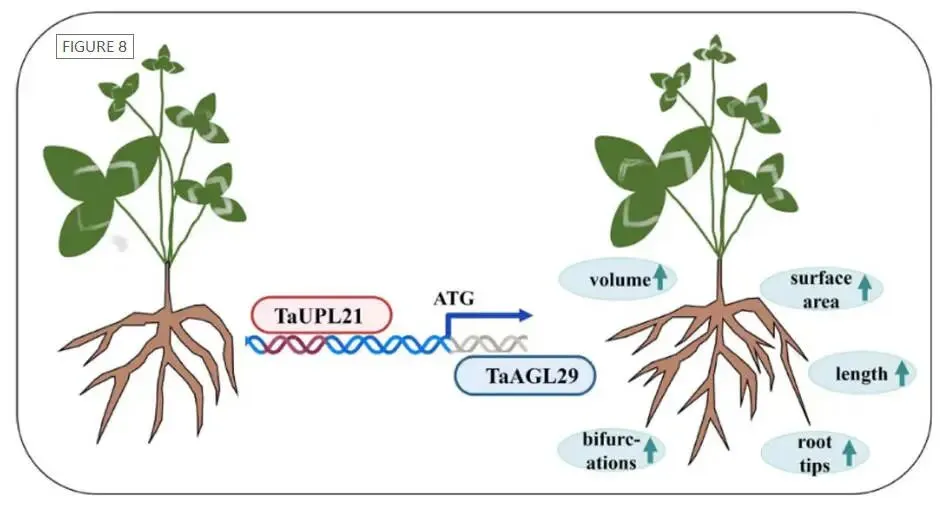
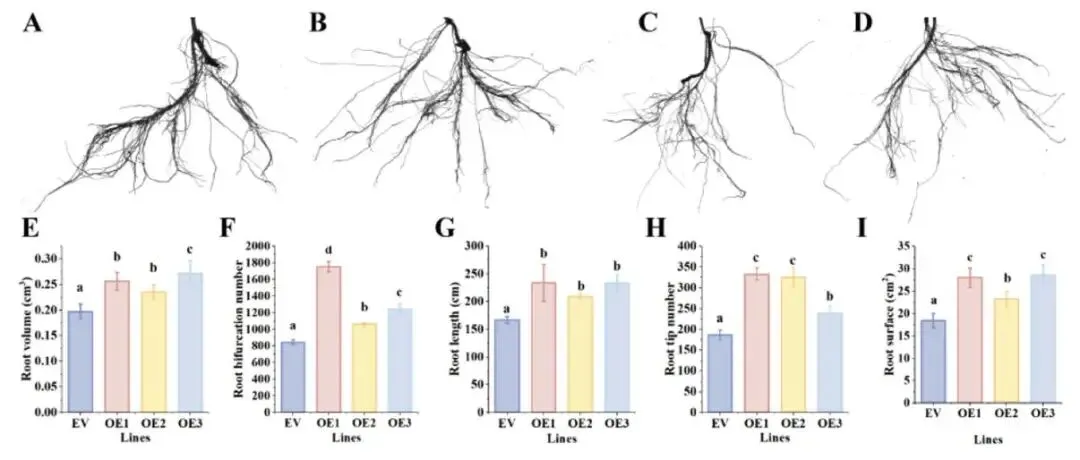
为深入探究TaUPL21的功能,通过农杆菌介导的遗传转化生成了过表达TaUPL21的毛状根。定量RT-PCR分析证实,转基因毛状根中TaUPL21的表达量显著高于对照组。利用根系扫描仪对过表达TaUPL21的毛状根进行表型分析,结果显示,与对照组相比,过表达株系在根长、根体积、根尖数、分叉数以及根表面积方面均表现出显著增加(p < 0.05)。其中,OE1株系的表型最为明显,各项指标分别达到对照组的1.4倍、1.3倍、1.7倍、2.0倍和1.5倍。这些结果充分表明TaUPL21正向调控高三叶的根系生长与发育(Fig.4)。
图4. 过表达TaUPL21促进高三叶毛状根发育 (A–D) 高三叶毛状根中过表达TaUPL21的表型。A:空载体(EV);B:OE-1;C:OE-2;D:OE-3。(E) 过表达TaUPL21的根体积。(F) 过表达TaUPL21的根分叉数。(G) 过表达TaUPL21的根长。(H) 过表达TaUPL21的根尖数。(I) 过表达TaUPL21的根表面积。不同小写字母表示同一株系在不同处理时间下的显著差异(p < 0.05)。
3.6 DAP-Seq揭示TaUPL21意想不到的DNA结合活性
DNA结合:DAP-seq分析鉴定出数百个高置信度结合峰,且多数位于启动子区域。
基序鉴定:识别出特异性结合基序,并明确了其在转录起始和终止位点附近的分布模式。
通路富集:靶基因显著富集于次级代谢、基因组稳定性、表观遗传调控及发育等过程。
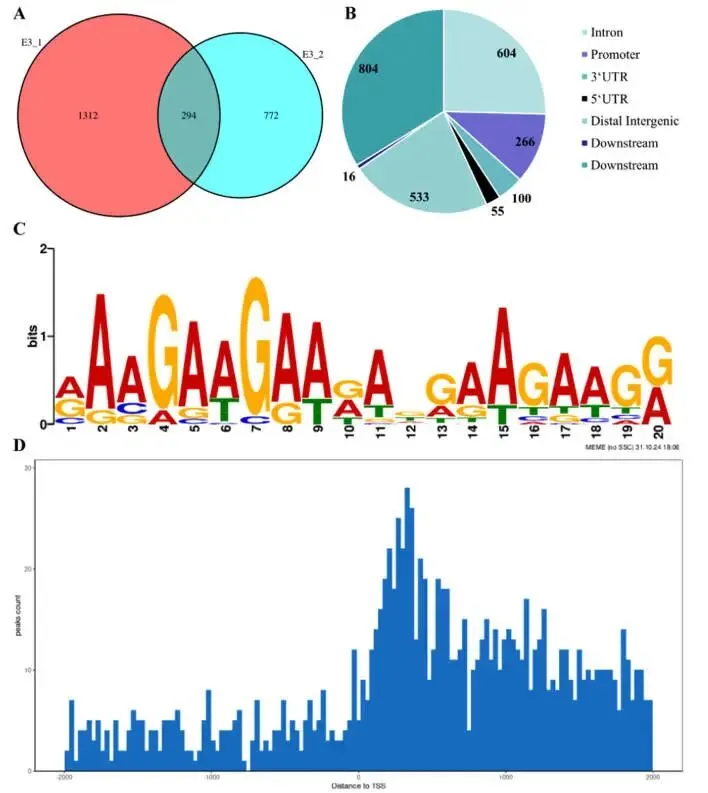
除了作为E3泛素连接酶的经典作用外,DAP-seq分析揭示了TaUPL21具有内在的DNA结合活性。对两个生物学重复的分析分别鉴定出1607和1066个结合峰,其中294个高置信度峰在两个重复中共有。值得注意的是,这些共有峰中有266个(11.2%)位于启动子区域,支持其在转录调控中的作用。从头基序分析鉴定出TaUPL21特异性的DNA结合基序。在转录起始位点(TSS)上游2 kb区域内,结合信号呈现均匀分布模式;而在转录终止位点(TTS)下游区域,大部分结合峰显著富集于近端下游1 kb区域内。KEGG和GO富集分析显示,靶基因在甜菜红素生物合成、萜类骨架生物合成、DNA复制、错配修复等通路,以及双链甲基化DNA结合、三肽转运和花发育正向调控等生物学过程中显著富集。这些结果表明TaUPL21蛋白参与了次级代谢、基因组稳定性、表观遗传调控、应激反应和发育等多种生物学过程(Fig.5)。
图5. TaUPL21 DNA结合活性的DAP-seq分析及功能验证 (A) TaUPL21 DAP-seq实验两次技术重复间Peaks的一致性分析。(B) TaUPL21结合Peaks在功能区域(如启动子、外显子和内含子)的基因组注释统计。(C) 从TaUPL21识别区间富集到的保守基序。(D) 所有TaUPL21靶基因结合位点中心到转录起始位点(TSS)的距离。
3.7 TaUPL21通过结合TaAGL29启动子发挥功能
靶标筛选:选取MADS-box转录因子TaAGL29作为候选直接靶标进行深入验证。
互作预测:分子对接预测显示TaUPL21关键残基与TaAGL29启动子形成特异性氢键。
转录激活:酵母单杂交和双荧光素酶实验证实TaUPL21直接结合并激活该启动子。
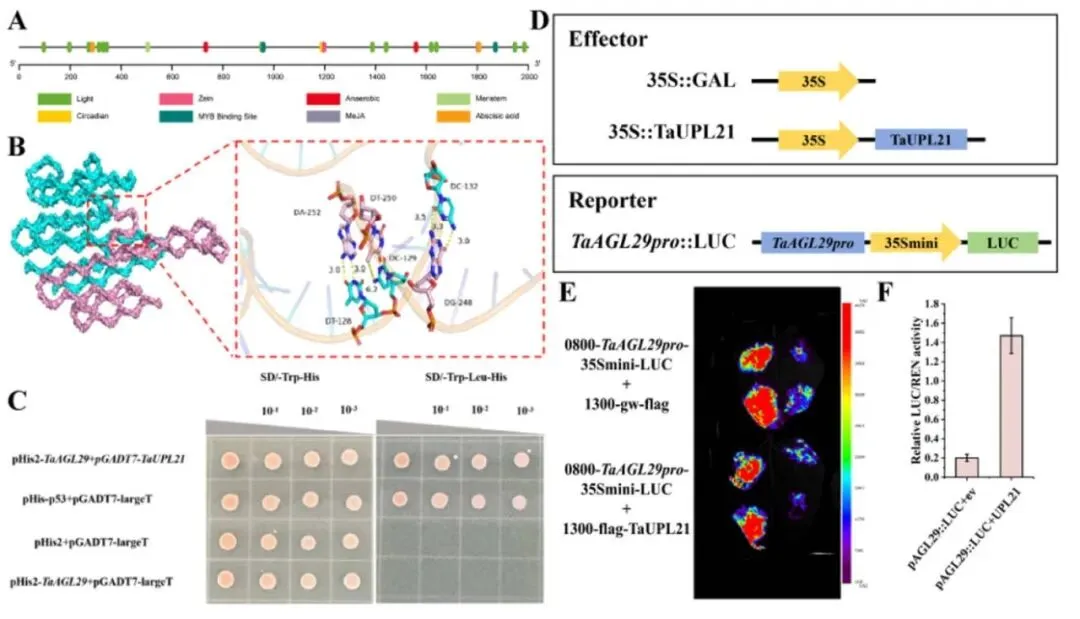
在DAP-seq鉴定的靶标中,MADS-box转录因子TaAGL29被选为候选直接靶标进行验证。系统发育分析确认了其与红三叶等物种中直系同源物的密切关系,且其启动子序列包含多种与光响应、分生组织表达及激素响应相关的顺式作用元件。分子对接预测了TaUPL21与TaAGL29启动子之间的特异性相互作用,TaUPL21中的关键氨基酸残基能够与启动子DNA上的核苷酸碱基形成氢键。在毛状根中过表达TaUPL21显著提高了TaAGL29的相对表达水平(p < 0.05)。随后,酵母单杂交(Y1H)实验证实了TaUPL21与TaAGL29启动子的直接结合。在本氏烟草叶片中进行的双荧光素酶报告基因实验进一步表明,共表达TaAGL29启动子-LUC报告基因与35S::TaUPL21效应物可显著提高LUC活性。这些结果确凿地证明了TaUPL21作为TaAGL29启动子的转录激活因子发挥作用(Fig.6)。
图6. TaUPL21直接结合并下调TaAGL29 (A) TaAGL29启动子序列及重要顺式作用元件的分布。(B) TaUPL21与TaAGL29启动子的结合位点。字母代表氨基酸,数字表示具体氨基酸位置。关键残基以杆状显示。黄色虚线代表蛋白结合能力,越短表示结合能力越强。(C) 酵母单杂交实验显示TaUPL21结合TaAGL29启动子。菌落体积被稀释10、100和1000倍。SD/-Trp-His为缺Trp和His的SD培养基;SD/-Trp-His-Leu为缺Trp、His和Leu的SD培养基。(D) 瞬时表达实验中使用的效应物和报告物构建体示意图。由CaMV 35S启动子驱动的TaUPL21作为效应物。将TaAGL29启动子中的TaUPL21结合位点连接到报告载体中。(E) 瞬时表达实验中TaUPL21对TaAGL29启动子的转录调控。(F) 转化杨树原生质体12小时后测定的LUC与REN相对荧光素酶活性。
3.8 TaAGL29正向调控根系生长
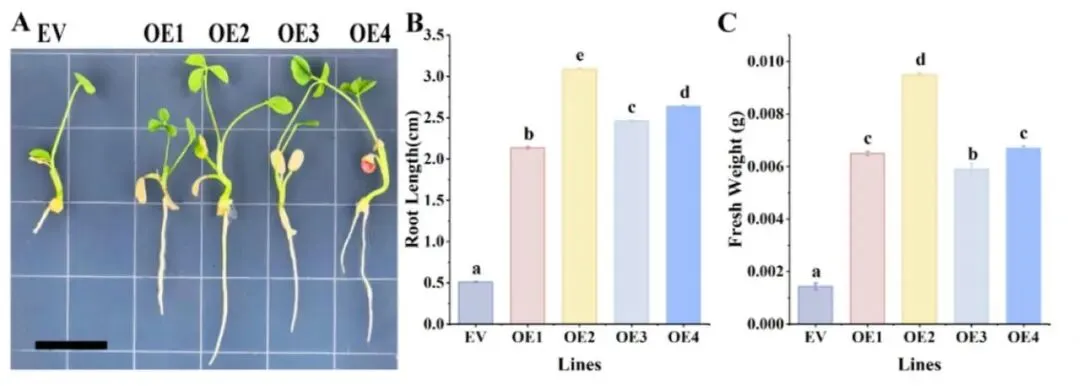
为了更好地理解TaAGL29在高三叶中的功能,通过毛状根遗传转化系统获得了过表达TaAGL29的毛状根。RT-qPCR结果显示,毛状根中TaAGL29的表达水平显著高于空载体(EV)对照(p < 0.05)。在过表达TaAGL29的毛状根中,观察到了与过表达TaUPL21相似的表型结果。转化14天后,过表达TaAGL29的毛状根长度显著增加(p < 0.05),最长可达3.10厘米。此外,过表达TaAGL29的毛状根鲜重也显著高于对照组(p < 0.05)。同时意外发现部分过表达株系表现出更强的生根能力,提前长出多条根系。这些结果充分表明TaAGL29正向调控高三叶的根系发育(Fig.7)。
图7. 过表达TaAGL29促进高三叶毛状根发育 (A) 高三叶毛状根中过表达TaAGL29的表型。在培养皿中观察TaAGL29的过表达情况,比例尺代表每个小格的长度为1.4 cm。(B) 过表达TaAGL29的根长。(C) 过表达TaAGL29的根鲜重。使用直尺进行测量。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
