羽衣甘蓝(Brassica oleracea var. acephala)是十字花科芸薹属二年生观叶草本花卉,叶形多样,耐寒性强,是适宜秋冬冷凉季节园林绿地应用的优良观赏植物。经过长期的自然变异与人工驯化选择,羽衣甘蓝形成了裂叶、圆叶、皱叶、羽叶等多种叶片形态,使其成为研究植物叶片发育的理想材料。近年来,对于羽衣甘蓝叶形的研究主要集中在裂叶性状遗传规律及其候选基因定位方面,但是裂叶形成的分子机制尚不明晰。深入研究羽衣甘蓝叶缘发育的调控机制对定向培育优良叶形的羽衣甘蓝新品种具有重要的理论指导意义。近日,The Plant Journal在线发表了沈阳农业大学祝朋芳教授团队题为“BoSKU5-BoARF8 complex modulates leaf margin development via the BoUIF1-BoCUC2cascade in kale”的研究论文,该研究通过N-糖基化蛋白质组学分析鉴定出一个含铜氧还蛋白域的蛋白BoSKU5,并证实了N-糖基化修饰对于BoSKU5的核定位及其维持叶缘全缘的功能至关重要,揭示了BoSKU5/BoARF8-BoUIF1-BoCUC2模块在羽衣甘蓝叶缘发育中的新功能。1. BoSKU5在BoALG10S0835-3过表达株系中特异性发生N-糖基化修饰
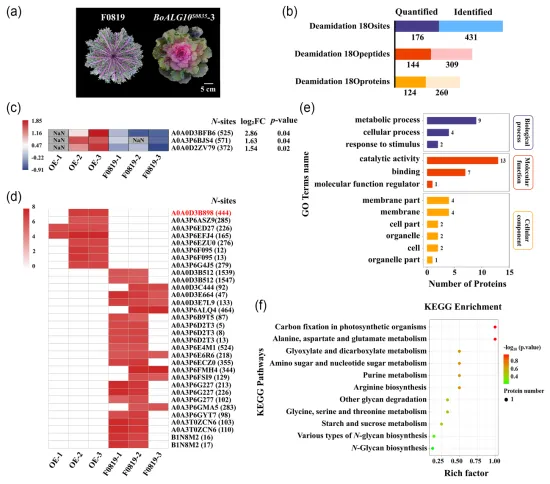
N-糖基化是植物中常见的蛋白质翻译后修饰,其中脂质连接寡糖(lipid linked oligosaccharide,LLO)在内质网中逐步装配形成完整的N-糖链前体。在LLO生物合成的最后阶段,α-1,2葡萄糖基转移酶(alpha-1,2 glucosyltransferase,ALG10)负责向寡糖链末端添加最后一个葡萄糖分子,ALG10功能缺失会破坏N-糖基化途径的正常进行并导致拟南芥叶片发育异常。在先前的工作中,研究人员已经证实BoALG10是控制羽衣甘蓝全缘叶性状的关键基因。在此次研究中,对BoALG10S0835-3过表达株系和裂叶野生型‘F0819’进行了N-糖基化蛋白质组学分析,发现BoSKU5蛋白在BoALG10S0835-3过表达株系中特异性发生了N-糖基化。此外,BoSKU5只有一个N-糖基化位点Asn-444在BoALG10S0835-3和‘F0819’之间表现出定量差异,表明该位点可能对BoSKU5的N-糖基化修饰具有重要影响。图1 BoALG10S0835-3过表达株系与野生型‘F0819’的N-糖蛋白质组学分析
2.N-糖基化影响BoSKU5蛋白的定位
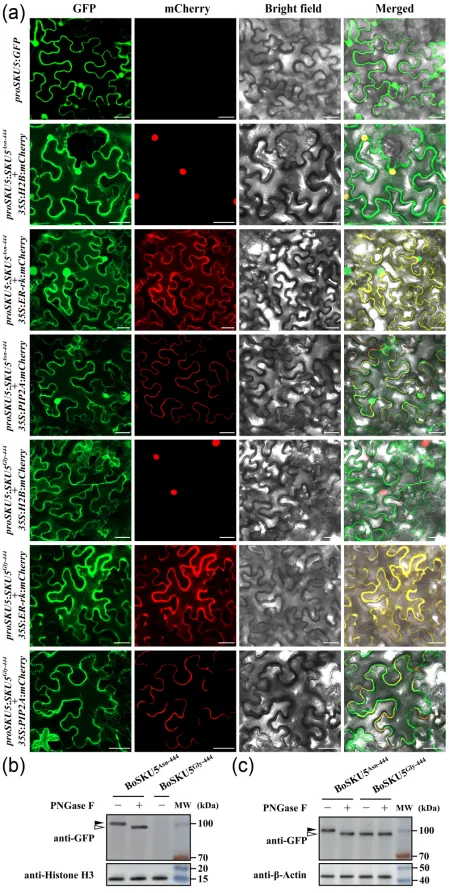
为确定BoSKU5的亚细胞定位,生成了由其自身启动子驱动的BoSKU5Asn-444-GFP构建体并在烟草叶片中瞬时表达,结果表明BoSKU5Asn-444-GFP主要定位于细胞核和内质网。当用甘氨酸(Gly)替代Asn-444位点后,BoSKU5Gly-444-GFP主要定位于内质网,而没有分布在细胞核中。免疫印迹分析进一步证实了Asn-444是BoSKU5蛋白一个重要的N-糖基化位点。图2 BoSKU5Asn-444和BoSKU5Gly-444的亚细胞定位及免疫印迹分析
3. BoSKU5维持羽衣甘蓝叶缘的全缘形态
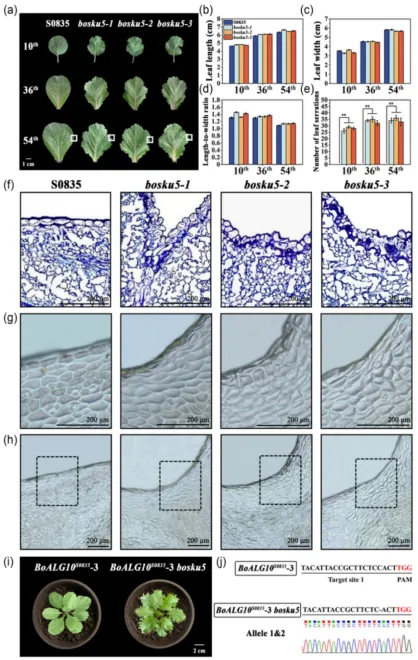
在羽衣甘蓝全缘叶自交系‘S0835’背景下,敲除BoSKU5基因显著增加了叶缘锯齿的数量。石蜡切片和指甲油印迹分析均显示bosku5突变体的叶缘处有许多形状不规则的表皮细胞紧密堆叠在一起。在BoALG10S0835-3过表达植株中敲除BoSKU5基因能够产生更深的锯齿,表明BoALG10调节叶缘发育的功能可能部分依赖于BoSKU5。此外,在bosku5突变体中重新引入BoSKU5Asn-444可以恢复突变体的全缘叶表型,而回补BoSKU5Gly-444则无法恢复突变体的叶片表型。图3 BoSKU5促进羽衣甘蓝全缘叶的形成
4. BoSKU5突变影响了生长素信号通路和细胞壁修饰相关基因的表达
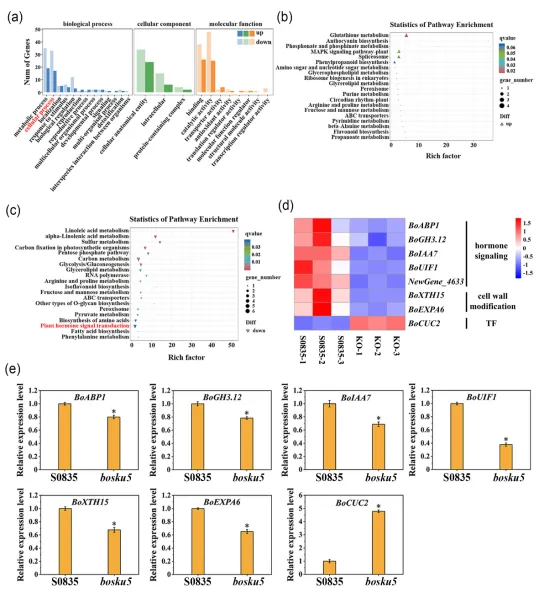
利用bosku5突变体进行转录组测序,共鉴定出222个差异表达基因(DEGs),其中包括99个上调DEGs和123个下调DEGs。GO分析表明,DEGs主要与“代谢过程”、“细胞过程”和“催化活性”相关。对于KEGG通路分析,“谷胱甘肽代谢”和“植物MAPK信号通路”包含的上调DEGs数量最多,而“植物激素信号转导”和“碳代谢”包含的下调DEGs数量最多。与野生型‘S0835’相比,生长素信号通路相关基因BoABP1,BoGH3.12,BoIAA7,BoUIF1以及细胞壁修饰相关基因BoXTH15和BoEXPA6在bosku5突变体中的表达水平显著下调。此外,一个已知的叶缘锯齿形成正调控因子BoCUC2在突变体中的表达水平显著上调。图4 bosku5突变体与野生型‘S0835’的转录组分析
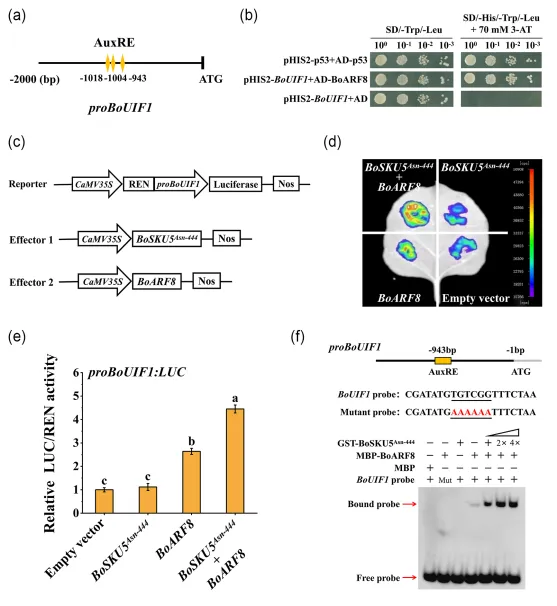
5. BoSKU5Asn-444-BoARF8蛋白复合物激活BoUIF1的转录
酵母双杂交筛库结果表明BoSKU5Asn-444在核内与生长素响应因子BoARF8形成蛋白复合物。通过CRISPR/Cas9技术生成了boarf8敲除突变体,证实了BoARF8能够促进羽衣甘蓝全缘叶的形成。双荧光素酶报告系统分析发现,BoSKU5Asn-444与BoARF8间的相互作用显著增强了BoARF8对其下游靶基因BoUIF1的转录激活能力。图5 BoSKU5Asn-444增强BoARF8对BoUIF1启动子的激活作用
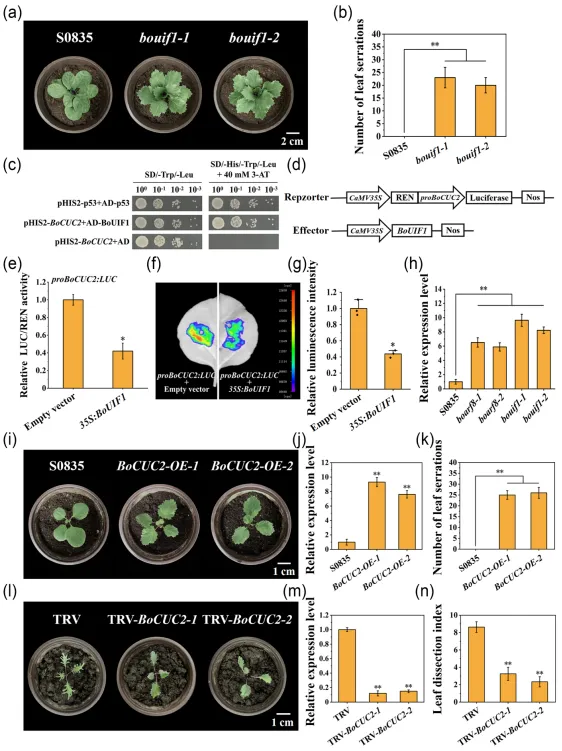
与野生型‘S0835’的全缘叶相比,bouif1突变体的叶片边缘产生了明显的锯齿。酵母单杂交与双荧光素酶试验证实BoUIF1可以结合BoCUC2基因的启动子并抑制其转录水平。RT-qPCR结果表明BoCUC2在boarf8和bouif1突变体中的表达量均显著上升。过表达BoCUC2显著增加了转基因植株叶缘锯齿的数量,而BoCUC2沉默植株则表现出叶缘锯齿深度的显著降低。
6 BoUIF1结合BoCUC2启动子并抑制其表达水平
研究意义
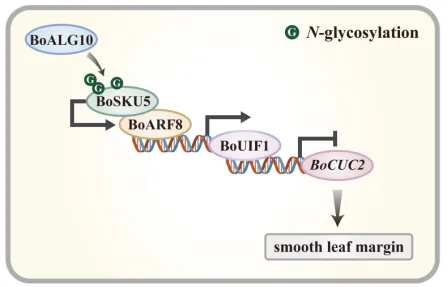
本研究揭示了N-糖基化修饰通过影响BoSKU5的核定位及其维持叶缘全缘的功能,进而参与调控羽衣甘蓝叶缘发育的作用机制,深化了N-糖基化修饰对于植物生长发育调控的认识。系统阐明了BoSKU5通过与生长素响应因子BoARF8形成复合物促进BoUIF1-BoCUC2转录级联的新途径,为羽衣甘蓝叶形分子育种提供精准靶点。在实践层面,该工作为培育具有优良叶形的羽衣甘蓝新品种提供了新策略,同时对其他园林植物的遗传改良具有重要参考价值。图7 BoSKU5调节羽衣甘蓝叶缘发育的模型图
沈阳农业大学祝朋芳教授团队长期从事羽衣甘蓝重要性状功能基因的研究,在羽衣甘蓝叶形性状方面,证实了BoALG10基因在维持叶缘全缘形态中的重要功能(Feng et al., 2020, Theoretical and Applied Genetics;Feng et al., 2022, Horticulture Research)。此次研究进一步鉴定出位于BoALG10下游参与叶缘发育的功能蛋白BoSKU5,构建了BoSKU5/BoARF8-BoUIF1-BoCUC2三层调控模块。在羽衣甘蓝其他重要经济性状方面,团队挖掘到了控制粉色叶性状的功能基因BoDFR1(Feng et al., 2021;Zhang et al., 2022, Theoretical and Applied Genetics),并解析了响应强光和低温转色的分子机制(Liu et al., 2024, The Plant Journal;Liu et al., 2024, Plant Physiology and Biochemistry);揭示了羽衣甘蓝蜡质合成功能基因BoORP3a的作用机理(Zhang et al., 2022, Horticulture Research);明确了羽衣甘蓝株高相关基因BoFHY1的生物学功能(Huo et al., 2025, Plant Physiology and Biochemistry)。目前,团队自主选育的羽衣甘蓝F1杂交新品种已推广应用。论文第一作者为沈阳农业大学林学院在读博士生张雨婷,通讯作者为沈阳农业大学林学院祝朋芳教授。青年教师冯馨以及团队研究生刘洋、金航标、李亚殊、方蕴蒙也参与了此项工作。该研究得到了国家自然科学基金(32171850和32471945)的资助。
论文链接:
https://onlinelibrary.wiley.com/doi/10.1111/tpj.70584

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
