ACS Catal. 中国农业科学院-ESTEEM策略:通过酶通道进化之门革新酶催化技术
2025 年 9 月 30 日,来自中国农业科学院的Ruyue Dong等人在ACS Catalysis上发表了一篇题为ESTEEM Strategy: Revolutionizing Enzymatic Catalysis through the Evolutionary Gateway of Enzyme Tunnels的研究性论文。Abstract
阐明酶的行为和设计高效生物催化剂仍然是生物催化领域的基础性挑战。尽管酶工程技术不断进步,但仍有巨大的优化空间。在此,提出酶效率模块的进化-结构微调(ESTEEM)策略,该策略将进化原理与结构骨架分析相结合,以加速酶的设计。以具有深埋活性位点和复杂底物通道的细胞色素P450单加氧酶为模型,识别了底物通道中的关键非保守残基(I67和Q83)。这些远端位点对通道结构具有决定性影响,其突变显著提升了催化效率。最优双突变变体的总转化数提高了106倍,且稳定性未受影响,同时拓宽了底物谱。通过适应性引导分子动力学和氢氘交换质谱分析,揭示了通道介导的调控机制:变体与野生型之间的活性差异由骨架的刚性-柔性平衡决定,该平衡通过稳定通道几何结构来调节催化效率。实验验证和回顾性分析证实了ESTEEM策略的稳健性。本研究阐明了通道突变如何稳定底物运输通道以增强催化作用,为针对需要深埋活性位点的底物设计具有特定活性的酶提供了理论框架——推动生物催化和合成生物学领域的应用发展。
Introduction
酶类以其无与伦比的催化效率和选择性,推动了生物过程、工业制造及合成生物学领域的变革性发展。然而,天然酶在工业应用中常因活性和稳定性的固有局限而表现不足。近期深度神经网络技术的突破性进展,推动了蛋白质设计的革新,加速了该领域的发展。值得注意的是,AlphaFold和RosettaFold已经彻底改变了深度学习辅助的蛋白质结构预测。尽管取得了这些进展,酶的理性工程改造仍面临挑战。虽然实验室进化有效,但该方法资源密集,且大多数突变会损害功能(30-40%)或属于自然变异(60-70%)。这源于复杂的序列-结构-功能关系,常导致进化轨迹偏离功能优化方向。优化这一探索过程需要缩小序列空间以筛选有益残基取代。
蛋白质的催化效率深深植根于跨越数十亿年的进化历史。因此,利用进化保守性是酶工程中一个至关重要但尚未充分探索的途径。酶家族中保守性低的残基往往蕴藏着未被开发的功能潜力,可用于增强活性、稳定性和底物特异性。靶向这些区域可以简化序列空间的探索,加速有益突变的发现。然而,目前仍缺乏一个将进化信息与三维结构数据相结合以指导酶优化的通用且稳健的框架。
尽管对蛋白质结构和催化功能的理解已取得显著进展,但决定酶效率的结构和分子决定因素仍未完全阐明。传统的理性设计主要关注基于结构数据和机制分析的活性位点突变、底物优化及蛋白质工程策略。然而,越来越多的证据表明,活性位点以外的区域——特别是远端调控和变构位点——是酶功能的关键调节因子。远端突变可通过影响构象动力学、平衡状态或底物通道几何结构显著改变催化活性。与此同时,进化分析(如祖先序列重建)为识别关键调控残基提供了新视角,揭示了进化过程中功能约束与序列变异之间的权衡关系。这些见解为理性选择酶的"可突变"区域奠定了基础。尽管已建立这一概念框架,但酶的复杂性阻碍了建立通用的活性增强准则。
在此,提出酶效率模块的进化-结构微调(ESTEEM)策略,这是一种探索非保守结构区域在酶调控中功能作用的创新方法(方案1)。ESTEEM整合了进化序列信息与三级结构特征,用以识别突变热点。以P450酶家族为模型验证该策略,重点关注3′-大豆苷元羟化酶——这一具有深埋活性位点和复杂底物通道的典型系统。通过综合突变分析发现,非保守残基(尤其是底物通道内的残基)比活性中心展现出更大的改造潜力。仅含两个突变的变体相较于野生型,在保持稳定性的同时显著提升了活性和偶联效率。这些结果支持通道作为进化调控热点的观点,并完善了ESTEEM的应用方向。此外,该变体的底物谱也得到了拓展。通过精细分析该变体在不同时间尺度上的结构动力学特征,阐明了增强型变体关键区域的构象变化,为酶功能优化提供了新见解。实验结果证明,ESTEEM不仅能成功预测和调控多种同源P450酶的活性,回顾性分析还表明,该策略为羧酸还原酶、酰胺酶、卤化酶及其他化工生产重要酶类的功能增强提供了宝贵的进化轨迹视角。本研究所阐明的理论通过挖掘催化过程中酶通道进化门控的潜力,加速了计算机设计酶的进化进程,并深化了我们对蛋白质科学中"序列-结构-动力学-功能"基本范式的理解。
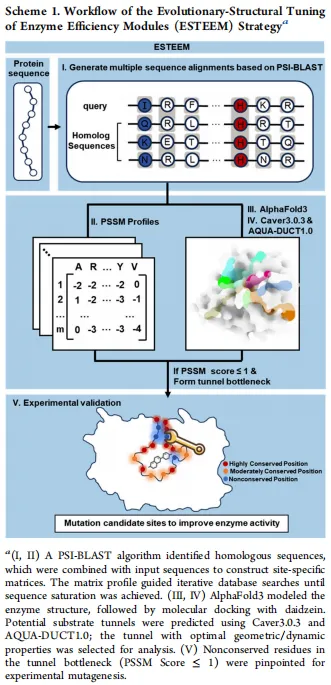
1. CYP107HB.am 的挖掘、异源表达及功能鉴定细胞色素P450单加氧酶具有多样化的催化活性,包括催化大豆苷元3′位羟基化反应。为挖掘具有潜在大豆苷元羟化活性的新型P450酶,我们基于已明确底物特异性的CYP105D7序列,利用HMMER v3.3.1构建了隐马尔可夫模型(HMM)。通过该模型筛选Uniparc数据库,共鉴定出9,210条候选序列。为降低序列冗余并聚焦于最具相关性的序列,应用CD-HIT工具对鉴定序列进行聚类分析,仅保留Cluster_number ≥ 100的序列簇。通过将序列与CYP105D7晶体结构进行比对进一步优化筛选,最终选取了6条均方根偏差小于2 Å的蛋白序列进行功能鉴定(图1A,B)。序列分析表明,这6种P450均属于I类细胞色素P450单加氧酶家族,其催化活性需依赖氧化还原伴侣。
为评估其功能潜力,构建了共表达六种P450及其氧化还原伴侣(铁氧还蛋白[Fdx]和铁氧还蛋白还原酶[FdR])的重组大肠杆菌菌株,并利用这些菌株催化大豆苷元的羟基化反应。在六种P450中,源自解淀粉芽孢杆菌的P450Bam展现出最高活性,能够将大豆苷元专一性地转化为3′,4′,7-三羟基异黄酮(3′-ODI),转化率达6%,与基准酶CYP105D7相当。相比之下,其他P450的转化率均低于3%(图1C)。这些结果确立了P450Bam作为一种新型细胞色素P450单加氧酶,能够催化大豆苷元的3′-羟基化反应。为深入鉴定P450Bam,将其与其他已知P450进行了多序列比对并构建了系统发育树。引人注目的是,P450Bam与源自枯草芽孢杆菌的P450Biol(CYP107H1)氨基酸同源性最高(62%)。基于该同源性及其功能特性,将此酶命名为CYP107HB.am。
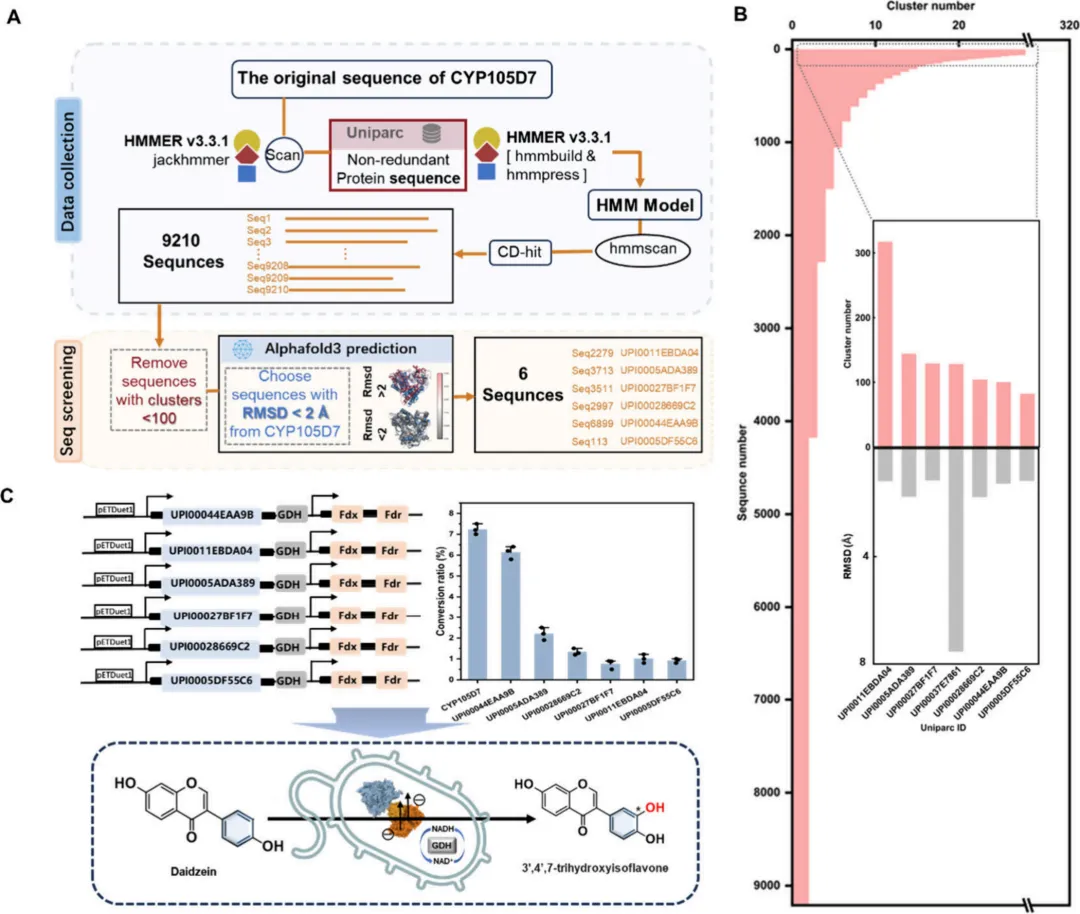
ESTEEM策略是通过一种系统性的方法开发的,该方法整合了进化见解与结构分析,为蛋白质工程提供了一个全面的框架。该方法的起点是使用位置特异性迭代基本局部比对搜索工具(PSI-BLAST)分析,识别CYP107HB.am中的进化非保守残基,随后构建CYP107HB.am的位置特异性得分矩阵(PSSM)。PSSM采用一种评分系统,其中正负整数表示特定位点的进化保守程度,得分越高代表保守性越强。具体来说,残基被分为三类:中度保守(1 < PSSM ≤ Max)、非保守(PSSM ≤ 1)和极度保守(20种氨基酸中的最大值)(图2A)。
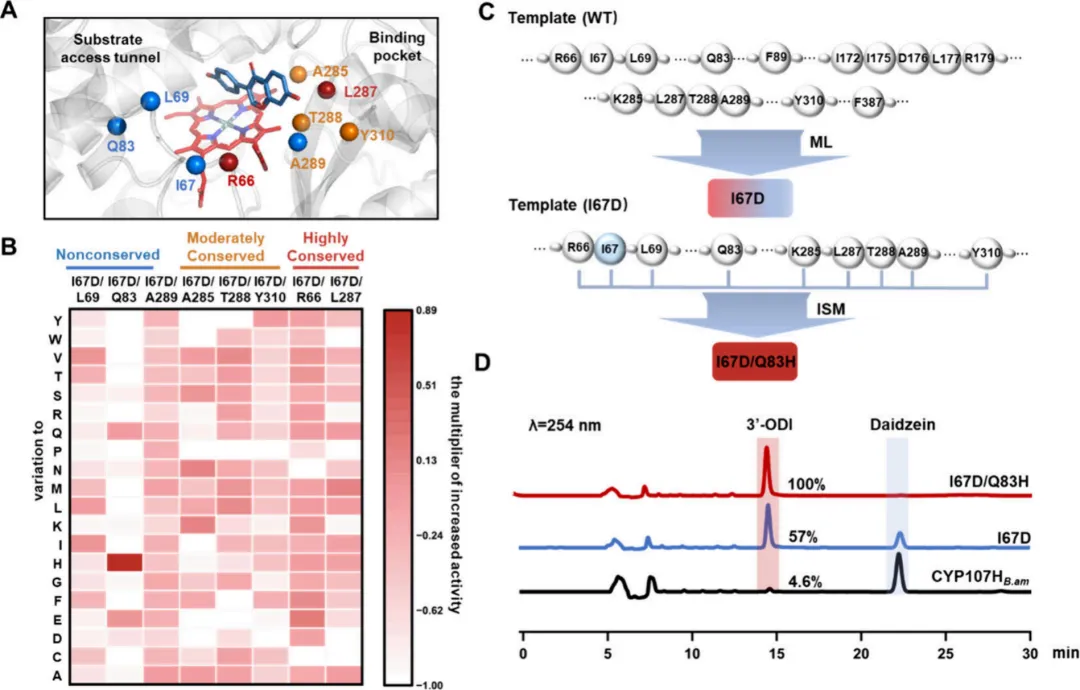
在此进化分析的基础上,采用系统性的方法来识别蛋白质结构中的全局突变热点。筛选策略涵盖两个不同的空间范围:(1)底物周围5 Å范围内的残基(第一壳层);(2)形成从第一壳层延伸至第四壳层的底物运输通道的残基(图2B)。对包含14个残基的活性中心的分析揭示了五个进化保守残基(Ile237, Ala238, His240, Glu241, 和 Thr242),它们构成了氧结合基序。考虑到修饰这些保守位点可能产生有害影响,选择了十个残基(Arg66, Ile172, Ile175, Asp176, Ala285, Leu287, Thr288, Ala289, Try310, 和 Phe387)进行后续的活性验证。
为阐明潜在的运输通道,采用CAVER 3.0.3进行通道分析。在12个已鉴定的通道中,通道1因其较高的流通率(0.737)被选为主要候选通道。为进一步证实该预测通道的生物学相关性,使用AQUA-DUCT软件进行了水分子运输分析。分子动力学模拟过程中溶剂分子的轨迹追踪显示,水分子的主要迁移路径在空间上与CAVER预测的通道1的几何结构(尤其是瓶颈区域附近)一致。这一动态验证支持所选通道作为真实功能路径的可靠性。进一步的详细结构分析聚焦于通道1内壁的残基组成,特别关注包括瓶颈和入口在内的关键区域。瓶颈区域代表通道的最窄部分,是配体运动的关键调节器,由六个关键残基(Ile67, Leu69, Gln83, Phe89, Leu177和Arg179)组成,这些残基随后被确定为工程改造的主要靶点。
对16个主要突变位点的评估揭示了不同的保守模式:四个非保守残基、五个中度保守残基和七个高度保守残基。基于进化原理,假设非保守和中度保守位点的修饰更有可能产生功能增强,而高度保守位点的改变可能会损害酶功能。为确保充分探索潜在的改进,对所有16个残基进行了完整的饱和突变,从而系统评估了每个位置上所有可能的氨基酸取代。
3. CYP107HB.am 突变可能性图谱的构建与迭代饱和突变利用ESTEEM策略,为CYP107HB.am内的16个候选位点构建了突变可能性图谱。共生成304个变体,并随后评估了它们对大豆苷元的羟基化活性(图2C)。初步筛选显示,所有四个非保守位点的突变均导致酶活性增强。值得注意的是,在位于酶底物通道内的残基Ile67和Gln83处观察到显著改进。在Ile67的19个变体中,大多数表现出活性增加,其中I67D显示出最显著的7.9倍增强。在Gln83处,仅鉴定出四个正向突变,其中Q83E的催化活性提高了6.7倍。此外,在五个中度保守位点中的三个位点,鉴定出转化率提高的变体(提高幅度从1.35倍到3.2倍不等)。相比之下,七个高度保守位点中仅有两个产生了活性增强的变体,尽管这些改进幅度不大,在1.22倍到1.27倍之间。
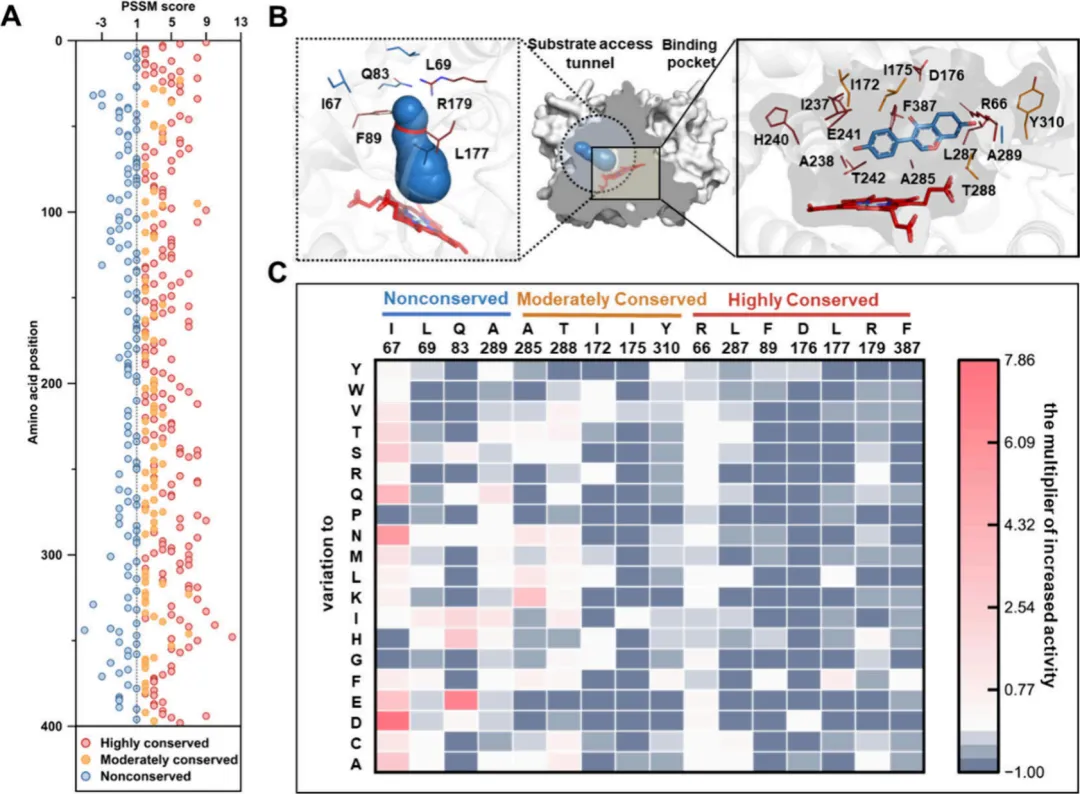
经过第一轮突变筛选,单点突变体I67D表现最为突出,在5小时内对50 μM大豆苷元的摩尔转化率达到了57%,而CYP107HB.amWT的转化率仅为4.6%。为进一步优化其活性,选择I67D作为模板进行第二轮迭代饱和突变。为减少上位效应,突变聚焦于另外八个正向突变位点(图3A,C)。该轮突变共生成152个双突变体(图3B),其中I67D/Q83H表现出显著的性能,其活性较野生型对照提高了21倍。具体来说,I67D/Q83H在5小时内实现了对50 μM大豆苷元的完全转化(100%)(图3D)。
这些发现表明,针对酶通道内进化非保守区域的改造工程,在调节催化活性方面比修饰活性中心更为有效。值得注意的是,与活性增强相关的四个非保守区域中有三个位于酶的通道内(图3A)。为验证这一观察结果,对通道和活性中心区域的突变热点进行了进化保守性与活性之间的相关性分析。皮尔逊相关性分析显示,通道瓶颈残基的PSSM得分与活性之间存在显著的负相关(皮尔逊相关系数 r = -0.90),表明在这些位置,酶活性随着进化保守性的降低而增加。相比之下,活性中心残基未观察到这种相关性(皮尔逊相关系数 r = -0.24)。这些结果强调了靶向进化非保守通道残基以利用进化原理增强酶催化活性的潜力。
4. CYP107HB.amWT 及其变体(I67D, I67D/Q83H)的性能表征为阐明 CYP107HB.amWT 及其变体之间催化效率的差异,对 CYP107HB.amWT、I67D 和 I67D/Q83H 催化大豆苷元羟基化的能力进行了全面的动力学分析。这些变体经过两轮严格筛选,被认为是最有前景的候选者。动力学分析表明,I67D 和 I67D/Q83H 均表现出显著的催化性能,其 kcat/Km 值分别为 2,191/M/min 和 7,923/M/min。与 CYP107HB.amWT 的 kcat/Km 值(97/M/min)相比,这些值分别代表了显著的 23 倍和 82 倍增强。I67D 催化效率的提高主要归因于 kcat 的大幅增加以及 Km 的降低。相比之下,I67D/Q83H 的显著性能则是由 kcat 的急剧增加驱动的,其 kcat 从 CYP107HB.amWT 的 0.03 min⁻¹ 上升至 5.63 min⁻¹。
总转化数进一步突显了变体增强的催化效率。具体而言,I67D 和 I67D/Q83H 的 TTN 分别为 4.93 和 15.99,与 CYP107HB.amWT 的 TTN(0.15)相比,分别增加了 32 倍和 106 倍。这强调了变体整体催化能力的显著提升。此外,观察到催化效率与偶联效率之间存在强正相关性。与 CYP107HB.amWT 相比,I67D 和 I67D/Q83H 的偶联效率从 0.6% 分别提高到 22.5% 和 69.85%,对应 36.5 倍和 115 倍的增强。为了排除蛋白表达水平对变体活性的影响,进行了 SDS-PAGE 分析,确认观察到的改进是蛋白质变体本身的内在特性。
为评估这些变体的实际应用可行性,使用 CYP107HB.amWT 及其变体在高底物浓度下进行了大豆苷元羟基化反应。CYP107HB.amWT 的转化效率随着底物浓度的增加而逐渐下降,对 1 mM 底物的转化率仅为 1.4%(3′-ODI 产量:3.78 mg/L)。相比之下,这些变体在相同条件下表现出显著的高转化率。具体来说,I67D 在 7 小时内转化了 69% 的底物(3′-ODI 产量:187 mg/L),而 I67D/Q83H 实现了 93% 的转化率(3′-ODI 产量:253 mg/L)。这是迄今为止报道的由 P450s 催化生产 3′-ODI 的最高转化水平。
还通过测定变体和 CYP107HB.amWT 的熔解温度评估了其热稳定性。结果显示,I67D(52.4 ± 0.10 °C)和 I67D/Q83H(53.5 ± 0.18 °C)的 Tm 值分别比 CYP107HB.amWT(50.8 ± 0.30 °C)高出 1.6 °C 和 2.7 °C。这些发现表明,这两种变体不仅表现出显著增强的活性,而且有效地平衡了活性与稳定性之间的关系。
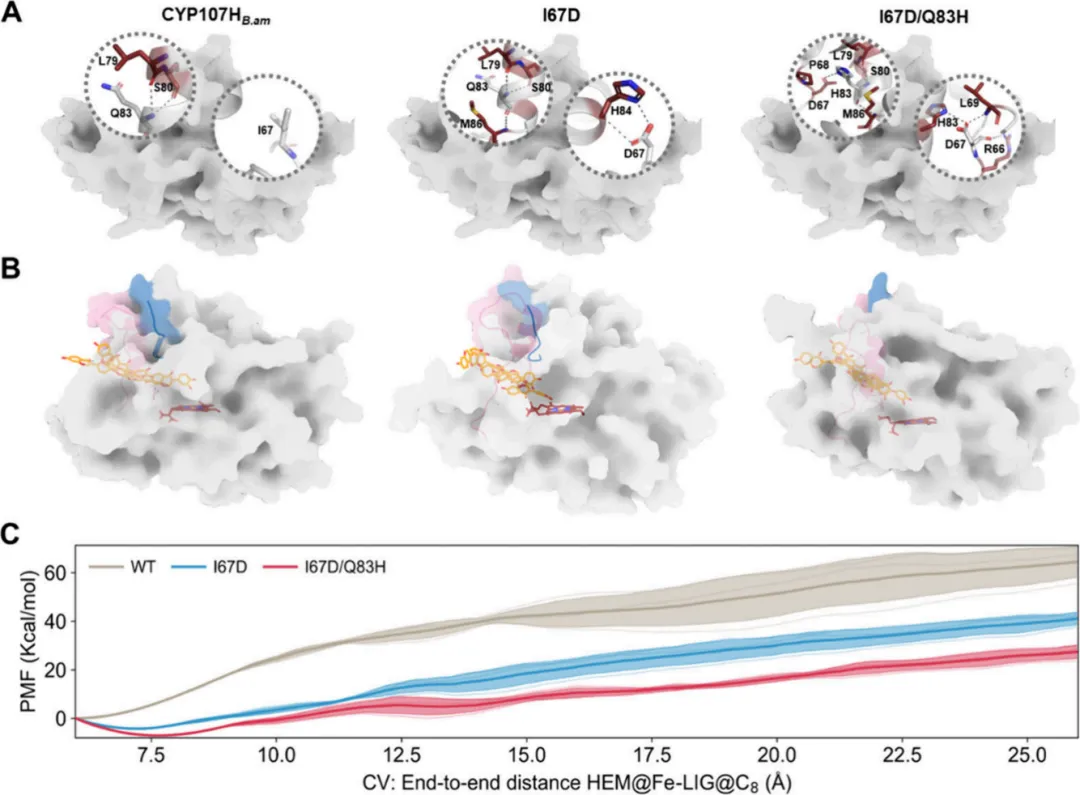
5. CYP107HB.amWT 及其变体中静态通道特性与配体输运效应的研究为了阐明在变体中观察到的催化活性增强的分子机制,采用了结合结构与计算分析的综合方法,包括分子对接和扩展的自适应引导分子动力学模拟。在 CYP107HB.amWT 中,未观察到与残基 Ile67 的相互作用。然而,当 Ile67 突变为 Asp 后,与 B' 螺旋上的 His84 形成了氢键和烷基疏水相互作用。此外,Gln83 与 Met86 形成氢键,并与 Leu79 和 Ser80 产生相互作用。当 Gln83 进一步突变为 His 时,Asp67 的相互作用得到进一步加强。此外,His83 还与 Pro68 形成了烷基-π 疏水接触(图 4A)。这些额外的相互作用有助于稳定 B-C 环的正确构象。
为探究底物或产物的运输机制,基于酶的几何特性进行了详细的通道分析。值得注意的是,与 CYP107HB.amWT 相比,变体中通道瓶颈的半径逐渐增加(从 1.210 Å 增至 1.769 Å)。此外,变体通道的长度缩短,平均曲率减小。这些发现表明,通过适当的构象重排,底物进入通道被成功改造,形成了一种更短、更直、更宽的新通道结构。此类修饰很可能提高了底物或产物的运输效率,为观察到的催化效率提升提供了一个合理的解释。
ASMD 模拟通过评估 CYP107HB.amWT、I67D 和 I67D/Q83H 中产物从催化活性中心解离到蛋白质表面的平均力势,进一步验证了假设。模拟结果显示,CYP107HB.amWT 沿解离路径表现出最高的自由能垒,表明显著的空间位阻阻碍了产物释放。相比之下,I67D 和 I67D/Q83H 的 PMF 曲线均显示出相较于野生型大幅降低的能垒,特别是在 12.5−22.5 Å 区域内,这表明空间位阻减小。值得注意的是,在整个解离过程中,I67D/Q83H 的 PMF 最小值几乎比野生型低 20 kcal/mol(图 4B,C)。这一观察结果与实验动力学数据一致(后者揭示了升高的 kcat 值),证实了引入的突变减轻了空间位阻并提高了催化性能。
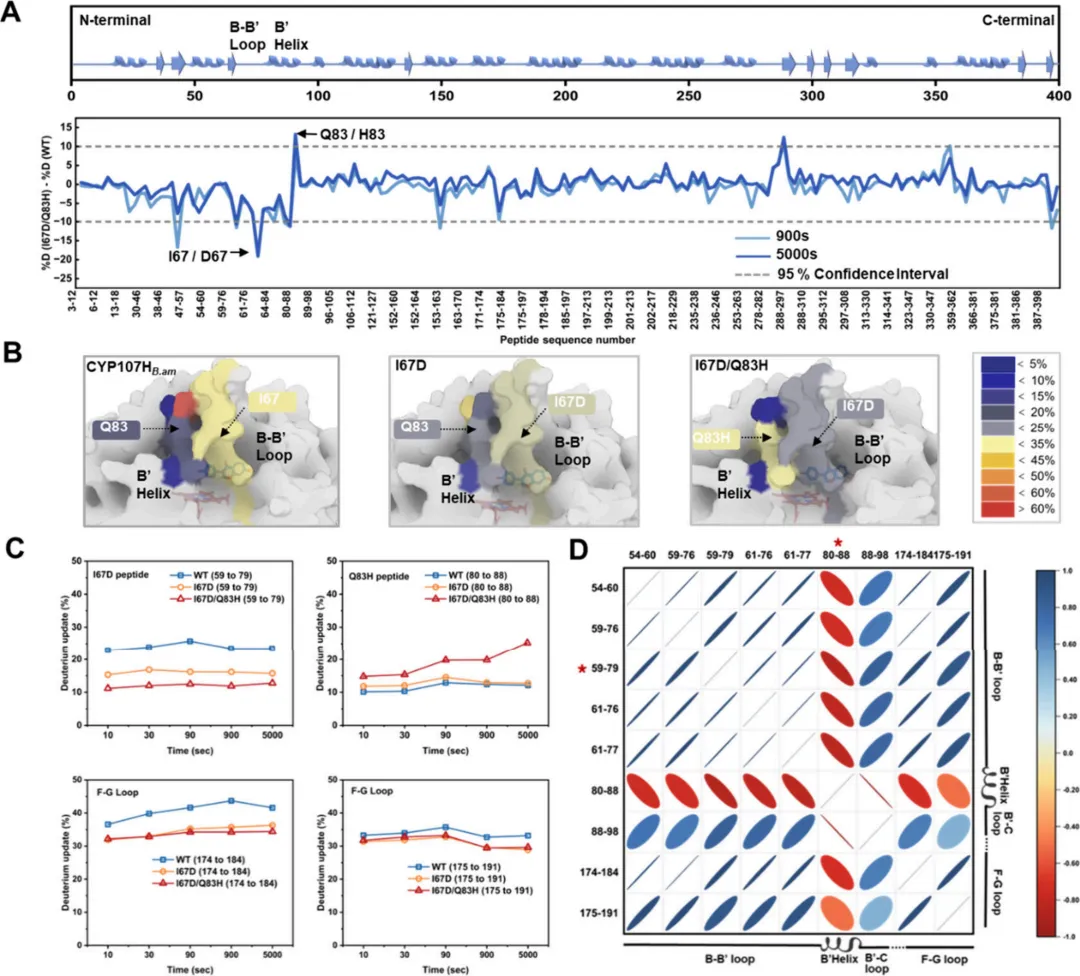
为阐明通道构象动力学对P450s活性的影响,采用HDX-MS研究长时间尺度下的通道构象动力学。HDX-MS技术通过主链酰胺氢与氘的交换,可深入揭示主链周围溶剂的可及性及动力学特征。通过分析数据并比较变体与CYP107HB.amWT特异性肽段的氘代百分比,鉴定了点突变后氘代水平发生统计学显著变化的区域。
HDX-MS谱图显示,在包含I67D和Q83H突变的区域,肽段水平的氘摄取存在显著差异(图5A)。具体而言,在CYP107HB.amWT中,包含I67D突变的肽段(B-B′环:肽段59−79)表现出约24%的高平均氘摄取率。相比之下,I67D和I67D/Q83H变体中该肽段的氘摄取率逐渐下降,分别为16%和12%。这表明B-B′环中的I67D突变导致动力学改变减少和溶剂化程度降低。相反,对于I67D/Q83H变体(19%),包含Q83H突变的肽段(B′螺旋:肽段80−88)的氘摄取显著增加,达到远高于CYP107HB.amWT(12%)和I67D(13%)的水平。这表明包含Q83H残基的B′螺旋具有显著更高的动态性和溶剂可及性(图5B,C)。此外,B-B′环和F-G环共同构成了通道的瓶颈区域。对与F-G环相关肽段(肽段174−184和175−191)氘摄取的比较分析显示,两个变体在该区域的氘摄取均有所下降,这可能归因于远程波动效应。通过HDX-MS观察到的这些氘代模式表明,由B-B′环、B′螺旋和F-G环构成的通道热点区域的动力学特征之间存在潜在关联。
为探究CYP107HB.amWT及其变体通道区域中相关肽段是否表现出协同运动,对0−5000秒内通道内所有肽段的平均氘摄取率进行了相关性分析。值得注意的是,在依次引入两个突变位点后,观察到B-B′环的运动趋势与B′螺旋呈显著负相关,而与F-G环呈显著正相关(图5D)。
由B-B'环、B'螺旋和F-G环构成的区域是主通道内的关键结构元件,该通道将深埋的活性位点与周围溶剂相连。突变引入该区域的动力学变化,形成了"柔性支架刚性化,刚性支架柔性化"的调节模式(图5B)。柔性支撑结构的刚性增强可减少通道周围环区的无效振荡,更有利于通道的正确形成。反之,B'螺旋柔性的适度增加可能进一步促进B-B'环的精确定位。因此,质谱数据验证了静态通道分析与ASMD模拟的结论,表明突变为通道赋予了显著的几何稳定性。这为通道动力学在P450s催化中的作用提供了切实的实验证据。
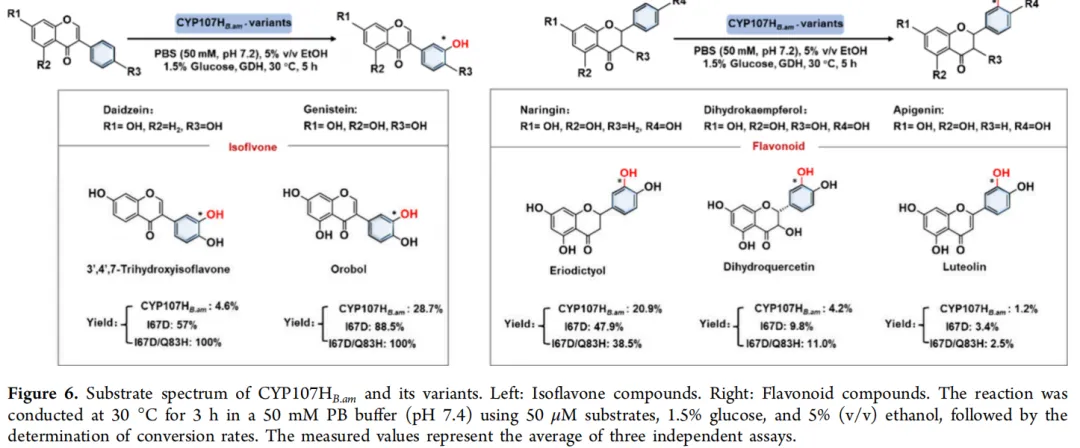
鉴于 I67D 和 I67D/Q83H 变体在大豆苷元 3′-羟基化反应中展现出相较于 CYP107HB.amWT 出乎意料的高转化效率,探究了这些最优变体是否也能对其他黄酮类化合物表现出增强的催化活性。为此,测试了 CYP107HB.amWT、I67D 和 I67D/Q83H 对四种不同底物的催化反应。
如图 6 所示,变体对这四种底物的转化率均有显著提高。值得注意的是,由于染料木素与大豆苷元结构相似,变体对染料木素的转化效率与对大豆苷元的转化效率非常接近。其中,I67D/Q83H 变体表现出最高的羟基化活性,转化效率达到 100%,较仅实现 28.7% 转化率的 CYP107HB.amWT 提高了 2.5 倍。此外,变体对其他三种黄酮类化合物——柚皮素、二氢山奈酚和芹菜苷元的催化活性也提高了 0.83 至 1.7 倍(图 6)。
为深入探究优势变体在调节底物特异性中的作用,对所有四种黄酮类底物进行了动力学参数分析。结果显示,变体对所有底物的催化效率(kcat/Km)均显著增强,提升幅度在1.1倍至6.5倍之间。此外,优势变体对异黄酮类化合物表现出明显的偏好性。四种底物的总转化数和偶联效率也同步反映了催化效率的变化:变体的TTN提升了1.5倍至115倍,偶联效率提高了1.1倍至105倍,证实其整体催化能力显著增强。综上,这些结果表明变体相较于CYP107HB.amWT具有明显优势,其在保持对天然底物活性的同时有效拓宽了底物特异性。
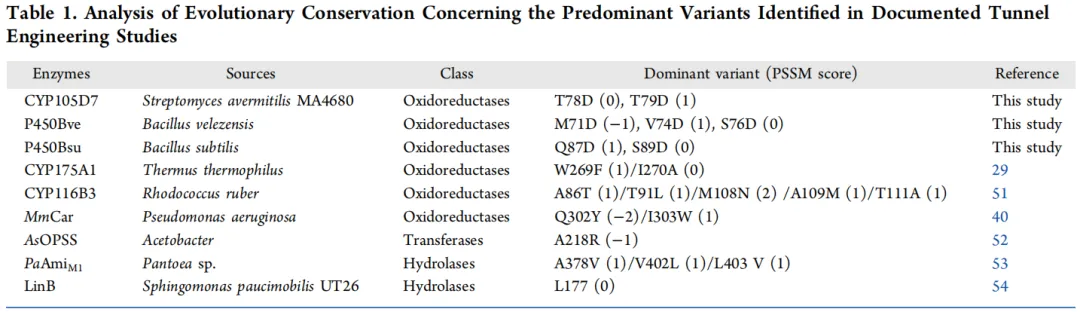
为验证ESTEEM理论的普适性,考察了其在其他P450酶中的适用性。基于前期研究中的三种P450酶,设计了突变热点以评估ESTEEM策略的有效性。具体而言,选取位于通道瓶颈区域的进化非保守氨基酸位点(PSSM评分≤1)进行实验验证。对于CYP105D7,单点变体的催化活性较野生型提高了1.83至2.9倍。P450Bve的三个变体表现出更宽的催化活性范围,为野生型的1.64至5.7倍。在P450Bsu中,单点变体Q87D的活性提高了1.5倍。这些发现共同凸显了ESTEEM策略在预测和调控多种同源P450酶方面的有效性。值得注意的是,不同酶的催化改进效果差异显著。为阐明这种差异的结构基础,我们比较了通道特性和活性口袋构型。对于CYP105D7变体,尽管平均半径和宽度增加,但通道曲率(平均值:1.062,接近线性)基本保持不变,限制了底物运输的整体增强。在P450Bve和P450Bsu变体中,通道长度的缩短部分优化了底物路径。然而,由于两种野生型的整体通道较宽(平均通道半径分别为1.789 Å和1.654 Å),这可能导致底物运输路径的自由度较高,从而削弱了突变带来的优化效果。因此,在具有宽或近线性通道的同源酶中,局部突变的几何扰动对底物迁移轨迹的改变极小,从而限制了催化活性的提升。该发现强调了在应用ESTEEM策略时,需评估目标酶的通道可调性——整合长度、半径和曲率等参数——以指导精确工程改造的必要性。
在现代生物催化快速发展的领域中,合理设计高效酶仍然是一项艰巨的挑战。通过借鉴自然的进化智慧,可以深入了解蛋白质的功能和结构。在此基础上,整合蛋白质结构数据以指导诱变策略,能够构建智能突变库,从而显著提高酶进化的精度和效率。本研究中开发的ESTEEM设计策略证明,识别并改造关键结构支架(尤其是底物通道)内的进化非保守位点,可以显著增强酶活性,且这种增强与通道动力学密切相关。成功实施该策略(将进化保守性分析与关键结构特征相结合),强调了将蛋白质进化见解融入酶工程的重要性。该策略在目标区域系统性地引入突变,大幅减少了实验工作量,同时保持了高成功率。
以大豆苷元3′-羟化酶CYP107HB.am为模型系统,在ESTEEM策略指导下,仅通过两轮进化就成功改造出一个双突变变体,其总转化数显著提高了106倍,且稳定性未受影响。研究结果表明,基于进化保守性的工程改造方法在调节通道特性方面比活性位点修饰更有效。这是因为通道突变位点与活性中心在空间上分离,避免了对底物结合口袋几何结构的直接干扰。因此,该策略能够协同优化酶的多种特性,同时最大限度地减少权衡效应。在多种P450同源酶上的实验结果表明,该方法可以有效识别高性能的单点变体。为进一步验证我们策略设计的合理性,我们对先前报道的有益通道工程改造突变进行了回顾性保守性分析。结果显示,活性增强的变体通常位于进化保守性低到中等的位点(表1),这与提出的筛选原则高度吻合,并为ESTEEM策略提供了有力支持。这种创新方法将蛋白质通道结构信息与先进算法相结合,能够精确操控蛋白质结构和功能以实现预期的催化效果,为未来的应用提供了巨大潜力。
“钥匙孔-锁-钥匙”模型为理解具有深埋活性位点和入口通道(称为钥匙孔)的酶的催化机制提供了一个可靠的框架。通过对催化增强因子的系统分析,确定优势变体主要调控 CYP107HB.am 通道的动态特性。这种调控类似于通过降低通道构象灵活性来维持最佳的“钥匙孔”构型。静态结构分析显示,突变增强了 B-C 环与 B' 螺旋之间的相互作用,从而改善了通道几何结构。然而,必须认识到酶通道是动态实体。采用 ASMD 模拟分析了配体通过蛋白质通道的运输,证实变体活性的增强源于有利于产物高效释放的最佳通道定位。这一发现代表了超越传统几何分析的重大进展。通过回顾性阐明,尽管大量计算研究预测了通道工程在改善酶功能方面的潜力,但实验验证仍然有限。HDX-MS 捕获了这些通道关键结构组件在长时间尺度上的动态变化,为蛋白质通道动力学在酶催化中的关键作用提供了经验证据。
总之,研究提出了一种综合策略,该策略整合了进化见解与结构信息以指导酶设计,并通过成功的工程应用得到了验证。通过对优势变体的详细分析,阐明了酶催化中进化门控的概念。发现不仅验证了所提出的概念框架,而且为未来的实验方法提供了明确的方向。该策略对于改造具有深埋活性位点的酶尤其有价值,并对不断增长的高效生物催化剂开发方法库做出了重要贡献。ESTEEM 方法强调进化保守性和通道动力学,为合成生物学和工业生物催化中的理性酶设计和优化开辟了有前景的途径。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
