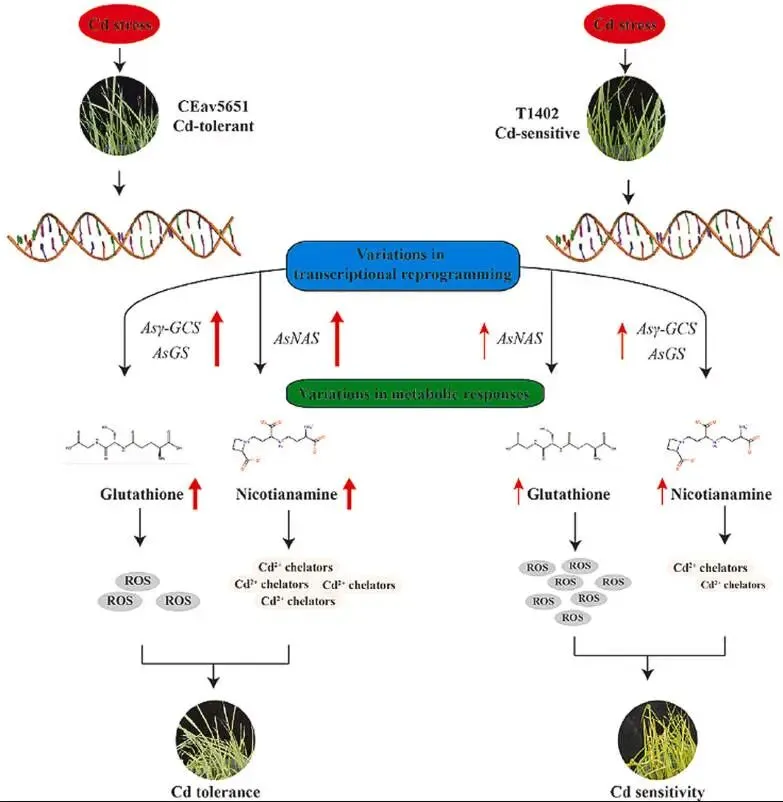
2.1 实验材料、培养与胁迫处理
实验选用两个对镉(Cd)耐受性差异显著的燕麦品种:敏感型T1402与抗性型CEav5651。种子经标准化程序消毒后,在黑暗中萌发72小时,随后移栽至蛭石与草炭土(1:1, w/w)的混合基质中。幼苗在生长室(相对湿度80%,光/暗周期16 h/8 h,温度22 °C/18 °C)中培养,并以霍格兰营养液灌溉。生长14天后,长势一致的幼苗被分为两组:对照组(CK)继续使用霍格兰营养液,处理组(Cd)则在营养液中添加终浓度为50 μM的CdCl₂。该浓度足以诱导胁迫响应并区分两品种的表型差异,但非致死。处理7天后,记录表型并采集叶片用于后续分析。为进行基因功能验证,野生型及转基因烟草(Nicotiana tabacum)先在1/2MS培养基上生长,后移栽至温室,并对30日龄的烟草进行类似的50 μM CdCl₂胁迫处理。植物组织中的Cd含量通过酸(HNO₃-HClO₄)消解后,采用电感耦合等离子体光谱法(ICP-OES)进行测定。
2.2 生理生化指标测定及多组学分析
为评估植物的生理状态,采用标准化方案测定了过氧化氢(H₂O₂)、超氧阴离子(O₂˙⁻)、丙二醛(MDA)的含量,以及超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性。同时,利用商业试剂盒和超高效液相色谱-串联质谱(UPLC-MS/MS)技术分别对谷胱甘肽和烟酰胺的含量进行精确定量。光合性能通过分光光度法测定叶绿素含量(Chl a, Chl b)和使用便携式荧光仪测定PSII最大光化学效率(Fᵥ/Fₘ)来评估。在分子层面,通过Illumina平台对四组样品(T1402-CK, T1402-Cd, CEav5651-CK, CEav5651-Cd)进行转录组测序(RNA-seq),并基于|log₂(倍比变化)| > 1和p-adj < 0.05的标准筛选差异表达基因(DEGs)。代谢组学分析则通过UPLC-ESI-MS/MS系统进行,并依据VIP ≥ 1和|Log₂(倍比变化)| ≥ 1的准则鉴定差异累积代谢物(DAMs)。
2.3 基因克隆、功能验证与数据统计
为验证关键代谢物的功能,对敏感型燕麦T1402进行了外源施用谷胱甘肽(1000 μM)和烟酰胺(500 μM)的保护性实验。同时,从抗性型燕麦CEav5651的cDNA中克隆了关键基因AsGS3和AsNAS17的开放阅读框(ORFs),并构建了由CaMV 35S启动子驱动的pCAMBIA1303表达载体。通过农杆菌介导的叶盘法将重组载体转化至野生型烟草中,以验证这两个基因在提升镉耐受性中的作用。此外,还利用MEGA-X软件构建了燕麦AsNAS基因家族的系统发育树。所有实验数据均采用SPSS 19.0软件进行统计分析,通过单因素方差分析(ANOVA)和Duncan新复极差法检验组间差异,以p < 0.05作为统计学显著性判断标准。
3.1 CEav5651的镉耐受性强于T1402
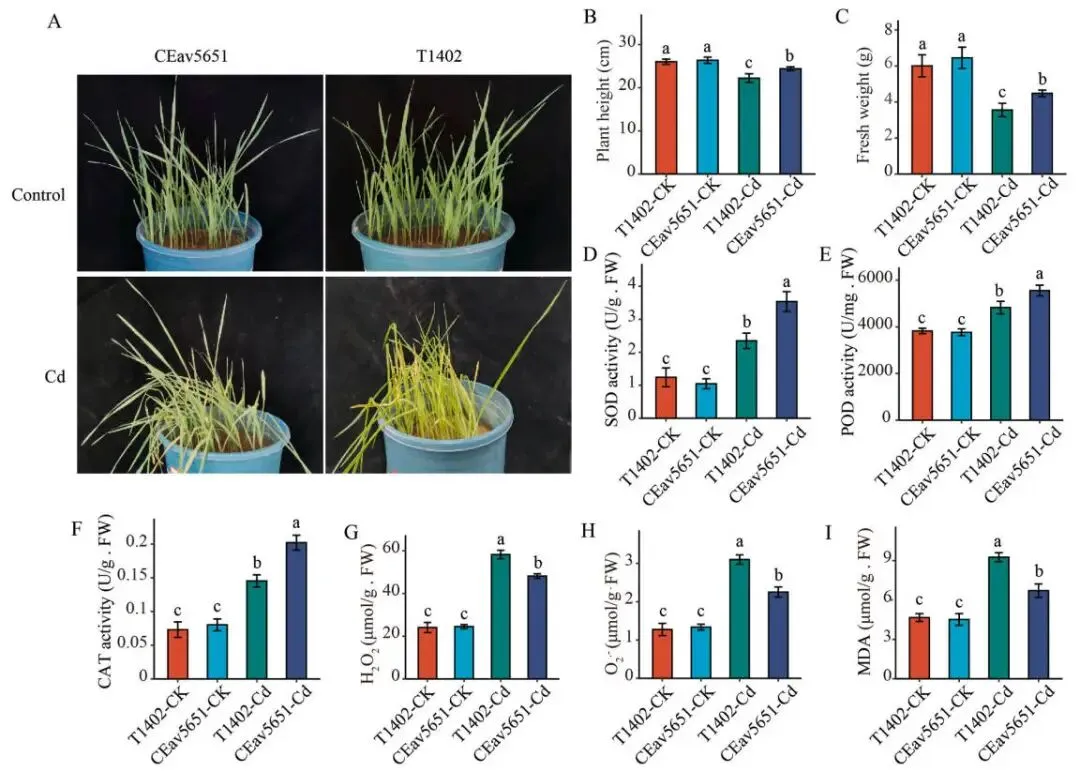
表型差异: 在Cd胁迫下,T1402表现出更严重的黄化、萎蔫、矮化和倒伏。
生长指标: T1402的株高和叶片鲜重显著低于CEav5651。
氧化损伤: T1402叶片中积累了更多的H₂O₂、O₂˙⁻和MDA。
抗氧酶活: CEav5651的CAT、POD和SOD活性显著高于T1402。
为比较对镉毒性的敏感性,对CEav5651和T1402幼苗进行了为期一周的50 μM CdCl₂处理。在无胁迫条件下,二者生长无明显差异。然而,在镉胁迫下,表型变化显著,敏感型T1402表现出更严重的黄化、萎蔫、矮化和倒伏症状。与表型一致,T1402的株高和叶片鲜重显著低于耐受型CEav5651。此外,T1402叶片中积累了更高水平的H₂O₂、O₂˙⁻和MDA,表明其遭受了严重的氧化损伤。而在抗氧化酶方面,CEav5651的CAT、POD和SOD活性均显著高于T1402。这些结果共同表明,镉胁迫抑制了燕麦的正常生长,且CEav5651相较于T1402具有更强的镉耐受能力 (Fig. 1 A–I)。
图1. 镉胁迫下T1402和CEav5651的表型及生理指标。 (A) 经50 μM CdCl₂处理一周后T1402和CEav5651的表型。(B–I) 在对照(CK)和CdCl₂处理(Cd)条件下的株高(B)、鲜重(C)、超氧化物歧化酶(SOD)活性(D)、过氧化物酶(POD)活性(E)、过氧化氢酶(CAT)活性(F)、过氧化氢(H₂O₂)含量(G)、超氧阴离子(O₂˙⁻)含量(H)以及丙二醛(MDA)含量(I)。误差棒代表标准差(SD; n = 6)。不同小写字母表示经单因素方差分析和Duncan新复极差法检验后差异显著(P < 0.05)。
3.2 镉胁迫下燕麦幼苗的转录组分析
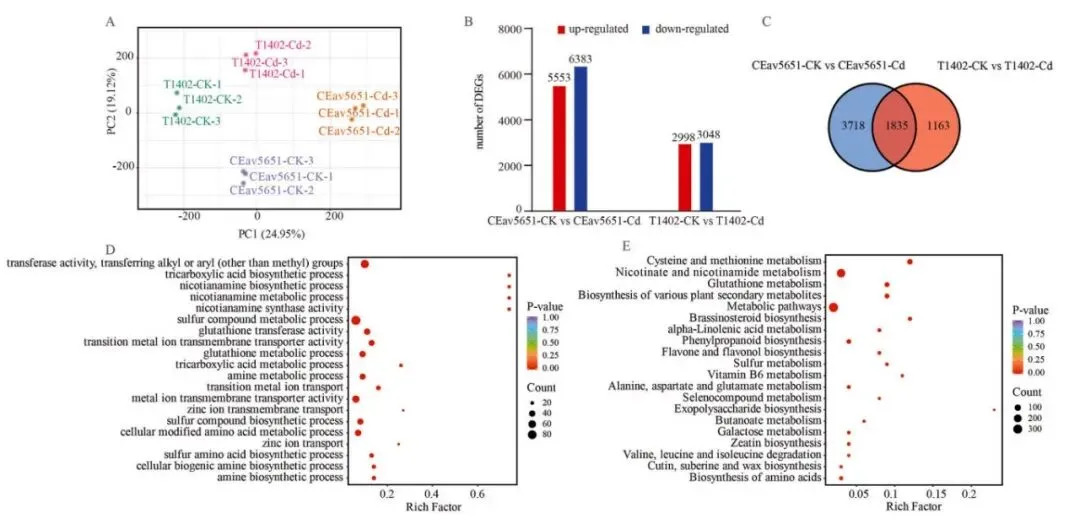
整体差异: PCA分析显示,Cd胁迫下两品种的转录谱显著分离。
基因响应: CEav5651中差异表达基因(DEGs)的数量远多于T1402。
共调基因: 1835个DEGs在两品种中共同上调,参与Cd响应。
功能富集: GO和KEGG分析共同指向“谷胱甘肽代谢”和“烟酰胺代谢”。
特异调控: CEav5651特异上调的DEGs富集于次级代谢和信号转导通路。
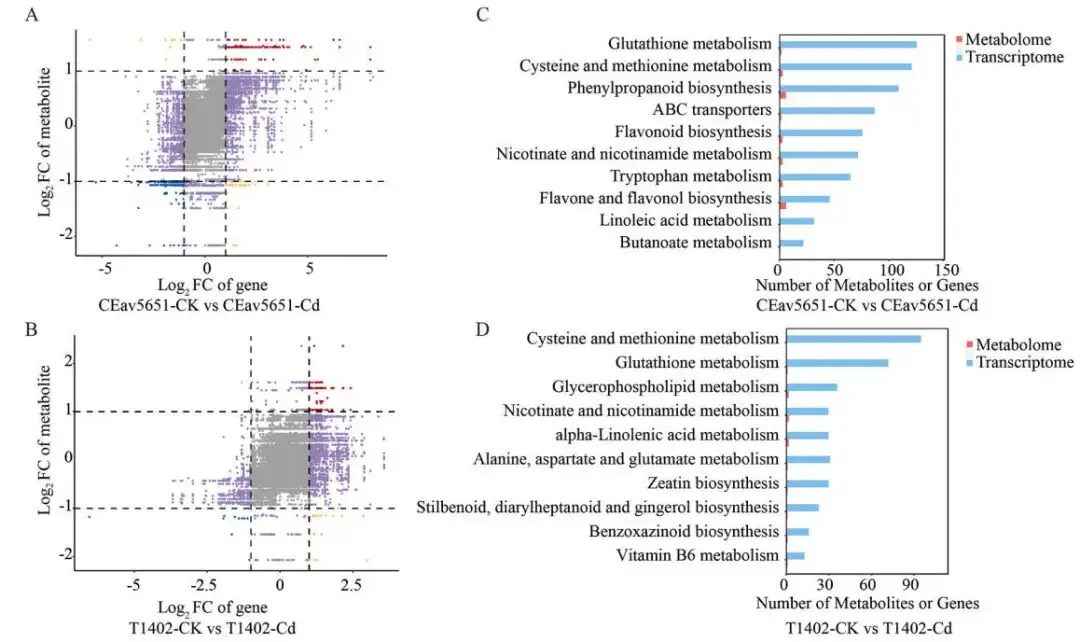
为深入理解镉胁迫响应的分子基础,对有无CdCl₂处理的CEav5651和T1402幼苗进行了转录组分析。主成分分析(PCA)显示,在镉胁迫下,两品种的转录组清晰分离,与表型差异一致。镉暴露在CEav5651中诱导了5533个基因上调和6383个基因下调,而在T1402中则分别为2998个和3048个。CEav5651中更多的差异表达基因(DEGs)表明,活跃的转录响应对于镉耐受性至关重要。交集分析发现1835个DEGs在两品种中共同上调,GO富集分析显示这些基因显著富集于“烟酰胺代谢过程”、“谷胱甘肽代谢过程”以及金属离子转运相关条目。KEGG通路分析进一步强调了“谷胱甘肽代谢”、“烟酸和烟酰胺代谢”和“半胱氨酸和甲硫氨酸代谢”的富集。此外,KEGG分析揭示,CEav5651中特异上调的DEGs主要参与“苯丙烷生物合成”、“黄酮生物合成”、“MAPK信号通路”和“植物激素信号转导”等,表明其优越的耐受性是通过胁迫信号和次级代谢的协同调控实现的 (Fig. 2 A–E, Fig. S1–S3)。
图2. 镉胁迫下T1402和CEav5651的转录组学分析。 (A) 不同品种和处理下转录组的主成分分析(PCA)。(B) Cd处理后T1402和CEav5651中上调和下调的差异表达基因(DEGs)数量。(C) 两品种在Cd处理后共同上调的基因数量。(D) 共同上调基因的基因本体(GO)富集分析。(E) 共同上调基因的京都基因与基因组百科全书(KEGG)通路富集分析。
3.3 镉胁迫期间的代谢组重编程
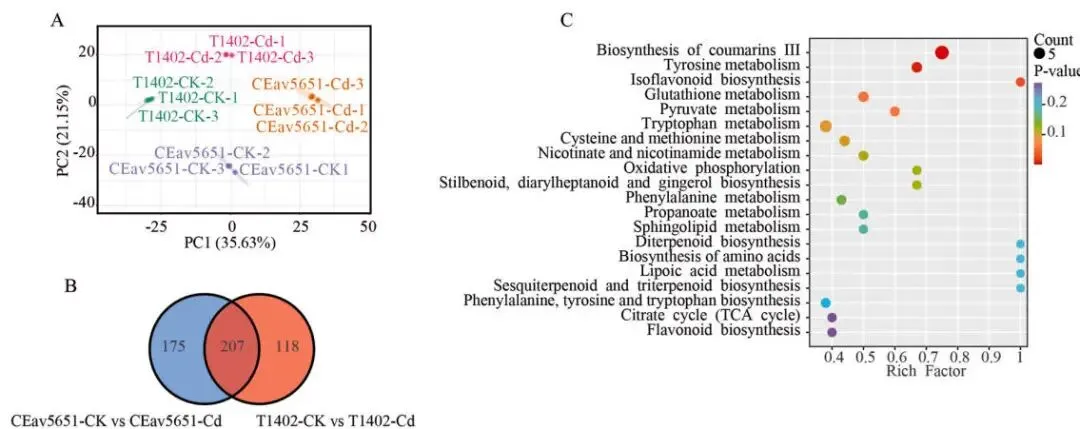
代谢差异: PCA分析显示,两品种的代谢物谱在Cd胁迫下存在明显差异。
差异代谢物: 在Cd暴露下,CEav5651产生了382个DAMs,T1402为325个。
关键物质: CEav5651中烟酰胺和谷胱甘肽的累积量分别比T1402高22.37倍和13.27倍。
通路富集: 共享DAMs的KEGG富集分析再次指向与转录组重叠的代谢通路。
采用UPLC-ESI-MS/MS系统分析了镉胁迫下燕麦幼苗的代谢组,共检测到1342种代谢物。基于代谢物的PCA分析进一步揭示了两品种间Cd耐受性的明显差异。筛选差异累积代谢物(DAMs)后发现,在镉暴露下,T1402和CEav5651中分别有325个和382个DAMs。在207个共享的DAMs中,有57个在CEav5651中的累积量显著高于T1402,其中烟酰胺和谷胱甘肽的含量分别高出22.37倍和13.27倍,这可能与其耐受性差异有关。对这些共享DAMs的KEGG富集分析揭示了与转录组数据重叠的四个通路:“谷胱甘肽代谢”、“半胱氨酸和甲硫氨酸代谢”、“烟酸和烟酰胺代谢”以及“氨基酸生物合成”。这些结果表明,谷胱甘肽和烟酰胺的生物合成是CEav5651增强镉耐受性的关键介导因素 (Fig. 3 A–C, Fig. S4–S5)。
图3. 镉胁迫下T1402和CEav5651的代谢组学分析。 (A) 所有检测到的代谢物的主成分分析(PCA)。(B) 显示两品种在对照(CK)和CdCl₂处理(Cd)条件下代谢物谱的韦恩图。(C) 对两品种共有的207个差异累积代谢物(DAMs)进行的KEGG通路富集分析。
3.4 转录组与代谢组的整合分析
关联分析: 在两品种中,众多DEGs与DAMs均表现出强正相关性。
共富集通路: 对比分析识别出前10个显著共富集的KEGG通路。
核心通路: “谷胱甘肽代谢”、“烟酸和烟酰胺代谢”及“半胱氨酸和甲硫氨酸代谢”这3个通路在两品种中均一致富集。
为探究DEGs和DAMs在镉胁迫下的相互作用,进行了全面的共表达网络分析。结果显示,在T1402和CEav5651中,众多DEGs与DAMs均表现出强的正相关性(Pearson相关系数 > 0.8, P < 0.01)。对转录组和代谢组数据集的KEGG富集分析确定了每个比较组中前10个显著共富集的通路。值得注意的是,“谷胱甘肽代谢”、“烟酸和烟酰胺代谢”以及“半胱氨酸和甲硫氨酸代谢”这三个通路在两品种的胁迫响应中均被一致地富集。这一整合分析揭示了基因表达与代谢物累积协同调控的机制,并确定了燕麦为有效适应镉毒性而采用的关键代谢途径 (Fig. 4 A–D)。
图4. 镉胁迫下转录组和代谢组数据的整合分析。 (A, B) CEav5651-CK vs. CEav5651-Cd (A) 和 T1402-CK vs. T1402-Cd (B) 比较组的基因-代谢物关联九象限图。(C, D) 两个比较组中,在相同通路共富集的DEGs(蓝色条)和DAMs(红色条)的KEGG富集分析。
3.5 外源谷胱甘肽和烟酰胺缓解镉毒性
表型缓解: 外源添加谷胱甘肽和烟酰胺显著减轻了T1402的叶片萎蔫。
氧化减轻: 处理后MDA、O₂˙⁻及H₂O₂含量显著降低。
抗氧增强: CAT、POD和SOD活性显著增强。
生长恢复: 叶片鲜重和总叶绿素含量分别增加了高达60.9%和45.7%。
光合改善: PSII最大光化学效率(Fᵥ/Fₘ)显著提高。
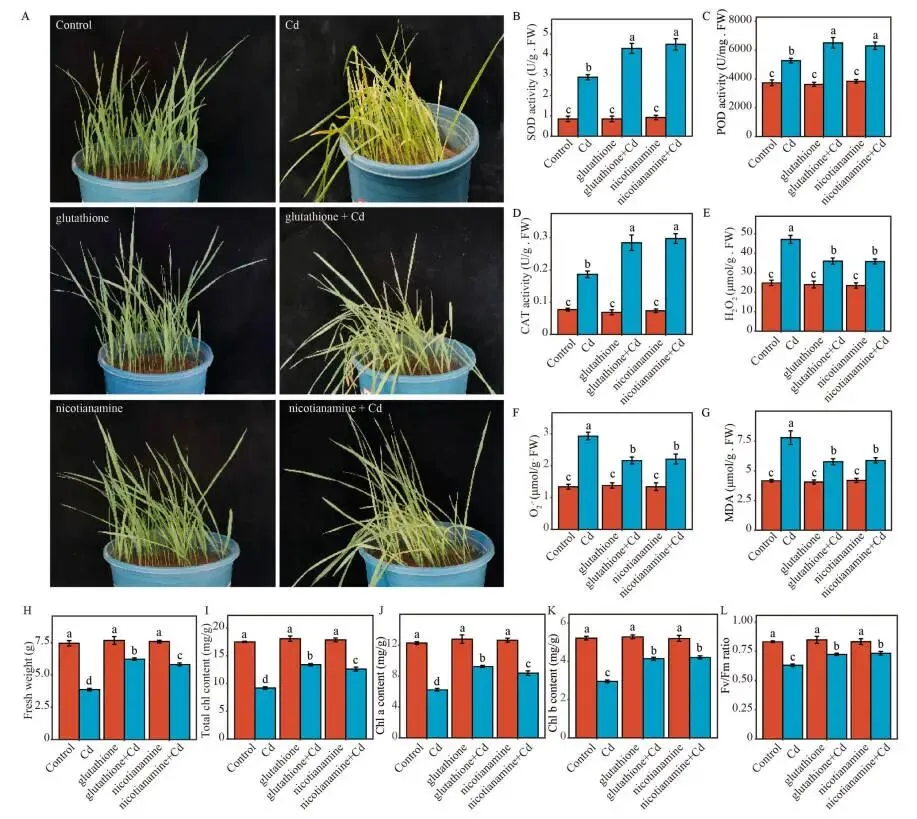
为功能性验证谷胱甘肽和烟酰胺的作用,将它们外源施用于镉敏感的T1402。结果显示,这两种代谢物显著减轻了镉诱导的叶片萎蔫,并大幅降低了MDA、O₂˙⁻和H₂O₂的累积,同时显著增强了CAT、POD和SOD的活性。与仅受镉胁迫的植物相比,经谷胱甘肽或烟酰胺处理后,叶片鲜重分别增加了60.9%和50.4%。此外,受镉胁迫严重降低的总叶绿素含量也分别恢复了45.7%和37.2%。光合色素的恢复进一步体现在光合器官完整性的改善上,PSII的最大光化学效率(Fᵥ/Fₘ)相较于单独镉处理组分别提高了约14.8%和16.2%。这些结果证明,谷胱甘肽和烟酰胺不仅通过增强抗氧化系统,还通过维持生长和光合能力来缓解镉毒性 (Fig. 5 A–L)。
图5. 外源谷胱甘肽和烟酰胺对燕麦镉耐受性的影响。 (A) 在指定处理下的燕麦幼苗表型:对照、Cd (50 μM CdCl₂)、GSH (1000 μM 谷胱甘肽)、NA (500 μM 烟酰胺)、GSH + Cd 和 NA + Cd。(B–L) 不同处理下的超氧化物歧化酶(SOD)活性(B)、过氧化物酶(POD)活性(C)、过氧化氢酶(CAT)活性(D)、过氧化氢(H₂O₂)含量(E)、超氧阴离子(O₂˙⁻)含量(F)、丙二醛(MDA)含量(G)、叶片鲜重(H)、总叶绿素含量(I)、叶绿素a (Chl a)含量(J)、叶绿素b (Chl b)含量(K)以及PSII最大光化学效率(Fᵥ/Fₘ)(L)。误差棒代表SD (n = 6)。不同小写字母表示经单因素方差分析和Duncan检验确定的处理间显著差异(P < 0.05)。
3.6 AsGS3介导的谷胱甘肽生物合成赋予镉耐受性
基因表达: 在CEav5651中,AsGS3基因受镉强烈诱导上调。
功能验证: 在烟草中异源过表达AsGS3显著增强了植物的镉耐受性。
生理效应: 转基因株系谷胱甘肽含量更高,氧化损伤指标显著降低。
Cd的分布: 转基因株系根系Cd含量更高,叶片含量平均降低55%。
转运降低: 生物富集系数(BCF)和转运系数(TF)分别降低了53%和65%。
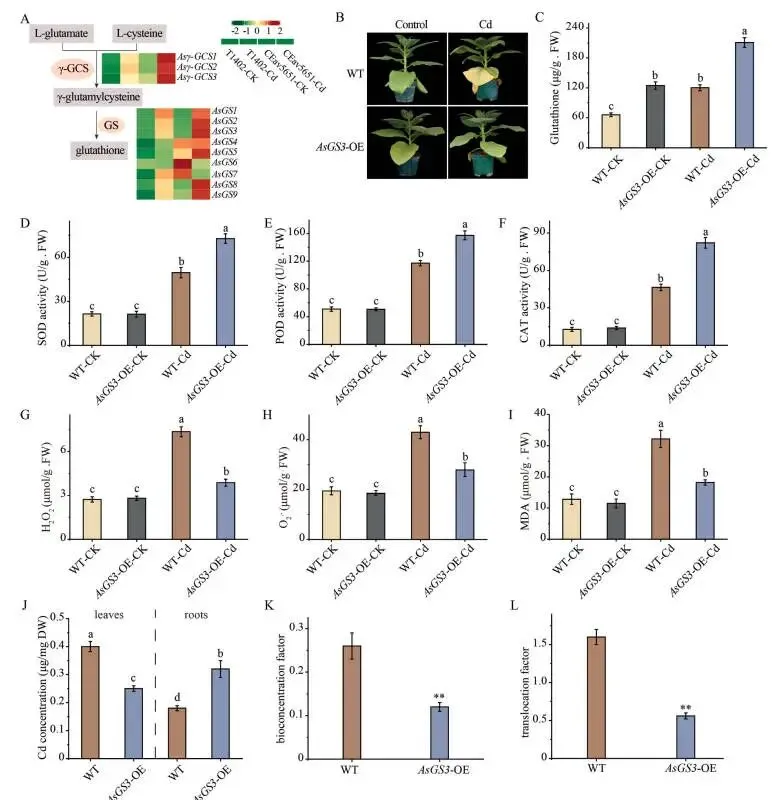
代谢组学显示,Cd胁迫的CEav5651中谷胱甘肽累积量最高。相应地,镉强烈诱导了CEav5651中谷胱甘肽合成酶(GS)基因的表达,其中AsGS3的上调最为显著。为探究其功能,在烟草中异源表达了AsGS3。在镉胁迫下,过表达AsGS3的转基因(AsGS3-OE)株系表现出更优的生长和更轻的萎蔫症状。与野生型(WT)相比,AsGS3-OE株系的SOD、POD和CAT活性显著增加,而MDA、O₂˙⁻和H₂O₂含量显著降低。转基因株系也积累了显著更高的谷胱甘肽。对Cd含量的测定发现,AsGS3-OE株系叶片中的Cd含量比WT平均低55%,而根部Cd含量则显著更高。这导致生物富集系数(BCF)和转运系数(TF)分别降低了53%和65%。这些数据表明,过表达AsGS3通过在根部强化镉的固存,有效减弱了Cd的总吸收及其从根到叶的转运 (Fig. 6 A–L)。
图6. 过表达AsGS3增强烟草的镉耐受性。 (A) 谷胱甘肽生物合成相关基因的表达。(B) 野生型(WT)和AsGS3过表达(AsGS3-OE)株系在对照和Cd (50 μM CdCl₂)处理下的表型。(C–I) WT和AsGS3-OE株系在对照和Cd处理下的谷胱甘肽含量(C)、SOD活性(D)、POD活性(E)、CAT活性(F)、H₂O₂含量(G)、O₂˙⁻含量(H)和MDA含量(I)。(J) 暴露于Cd胁迫的WT和AsGS3-OE植株叶片和根部的Cd浓度。(K, L) WT和AsGS3-OE植株的生物富集系数(BCF)(K)和转运系数(TF)(L)。误差棒代表SD (n = 6)。不同小写字母(C–I)表示组间的显著差异(P < 0.05)。星号(J–L)表示WT和AsGS3-OE株系间的显著差异(**P ≤ 0.01)。
3.7 AsNAS17的基因组鉴定与功能验证
基因鉴定: 在燕麦基因组中鉴定出23个AsNAS基因,分为两个进化枝。
表达模式: 进化枝I中的12个AsNAS基因受镉诱导上调,其中AsNAS17诱导倍数最高。
功能验证: 在烟草中异源表达AsNAS17显著减轻了镉胁迫下的叶片萎蔫。
生理效应: 转基因株系烟酰胺含量更高,氧化损伤指标显著降低。
Cd的积累: 转基因株系根和叶的Cd含量均升高,但转运系数无显著变化。
耐受机制: 其增强的耐受性归因于ROS清除能力的提升,而非Cd吸收的减少。
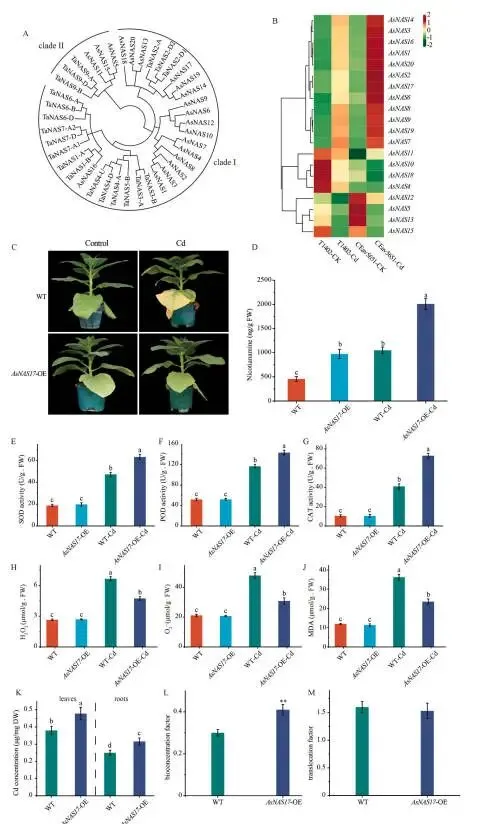
在燕麦基因组中鉴定出23个烟酰胺合酶(NAS)基因,系统发育分析将其分为进化枝I和II。转录组学数据显示,进化枝I的12个AsNAS基因在两品种中均受镉诱导上调,其中AsNAS17的诱导倍数最高。在烟草中异源表达AsNAS17后,转基因株系(AsNAS17-OE)在镉胁迫下表现出显著减轻的叶片萎蔫。与WT相比,AsNAS17-OE株系积累了更低的MDA、O₂˙⁻和H₂O₂,以及更高的SOD、POD和CAT活性。此外,转基因株系积累了更高的烟酰胺,表明AsNAS17在燕麦烟酰胺生物合成中起关键作用。对Cd含量的测定显示,AsNAS17-OE植株的叶片和根部Cd累积量均高于WT,其生物富集系数显著增加,而转运系数无显著变化。这些结果表明,AsNAS17赋予的镉耐受性并非源于减少了Cd的吸收,而是归因于ROS清除能力的提高 (Fig. 7 A–M)。
图7. 过表达AsNAS17增强烟草的镉耐受性。 (A) 小麦和燕麦NAS蛋白的邻接法系统发育树。(B) 不同条件下AsNAS基因表达谱的热图。(C) 野生型(WT)和AsNAS17过表达(AsNAS17-OE)株系在对照和Cd (50 μM CdCl₂)处理下的表型。(D–J) WT和AsNAS17-OE株系在对照和Cd处理下的烟酰胺含量(D)、SOD活性(E)、POD活性(F)、CAT活性(G)、H₂O₂含量(H)、O₂˙⁻含量(I)和MDA含量(J)。(K) 暴露于Cd胁迫的WT和AsNAS17-OE植株叶片和根部的Cd浓度。(L, M) WT和AsNAS17-OE植株的生物富集系数(BCF)(L)和转运系数(TF)(M)。误差棒代表SD (n = 6)。不同小写字母(D–J)表示组间的显著差异(P < 0.05)。星号(K–M)表示WT和AsNAS17-OE株系间的显著差异(**P ≤ 0.01)。