01
—
文章信息
作者单位:中国科学院应用生态研究所保护性耕作与生态农业重点实验室植物残留物最终是土壤有机物质的来源,但从相对新鲜的植物组织到高度进化的组分是一个连续体。然而,在保护性农业系统中,作物残留物作为植物组织与微生物尸体的累积程度仍然不确定。在东北地区进行了为期12年的玉米零收获、半收获和全收获保护性耕作试验。每隔1-4年采集0–10cm深度的土壤样本。对微生物残余物生物标志物氨基糖和植物成分生物标志物木质素苯酚进行了定量,以追踪微生物尸体和植物碎片的动态。与初始值相比,玉米秸秆覆盖12年后,氨基糖含量呈指数增长至21-45%,接近稳态平衡,表明在重复添加植物残渣的情况下存在“微生物承载能力”。随着保护性耕作持续时间的延长和玉米秸秆还田率的提高,氨基糖与木质素酚的比率从6.2降至1.7,同时木质素侧链氧化程度降低,表明有机输入在更大程度上增强了新鲜植物木质素的选择性保留。保护性耕作下土壤有机碳库的积累受到微生物和植物衍生成分功能互补的关键控制。玉米秸秆投入能够通过增加真菌坏死物质积累来提高微生物坏死物质对SOC稳定的贡献,而木质素类植物碎屑的保留扩大了SOC池容量,并可能长期保持SOC的可分解性。
—
引言
土壤有机碳(SOC)库在陆地碳(C)循环中起着关键作用,SOC积累决定了全球碳汇的增加潜力。从根本上说,土壤有机质(SOM)是通过复杂的分解过程从植物中提取出来的,被认为是从完整的植物碎片到高度进化的微生物C的连续体。植物残体输入的变化可能会改变土壤微生物的底物利用策略,从而影响植物和微生物来源的成分的保留,并相应地决定SOC的积累和稳定。因此,区分植物和微生物来源成分的动态变化对于阐明SOC池的建立过程和可持续功能至关重要。
由于绝对量化SOM连续体中微生物和植物衍生成分的浓度极其困难和耗时,因此可以使用不同来源的生物标志物来评估异质成分的相对变化。氨基糖是微生物细胞壁的重要组成部分,可作为土壤中微生物死亡质量的指标。与具有高周转率的易感微生物生物量相比,微生物坏死块的积累基本上与微生物在增殖和死亡过程中能够维持的最大C量有关,因此可以推断“微生物携带能力”。在可检测的氨基糖中,胞壁酸(MurN)来源于土壤细菌肽聚糖,而真菌几丁质主要贡献葡萄糖胺(GluN),尽管细菌细胞壁也有助于土壤中GluN的积累。因此,GluN与MurN的比率可以反映真菌和细菌坏死块的相对保留。木质素的酚类成分已被证明与植物组织分解的程度有关,植物组织可分为酚香草基(V)、丁香基(S)和肉桂基(C)化合物。V(Ac/Al)V和S单元(Ac/Al)S的酸与醛的质量比用于反映木质素苯酚中侧链的氧化程度。
在农业生态系统中,作物残体分解是SOM维持和积累的主要途径。通过追踪同位素标记的凋落物分解,Cotrufo等人(2015)认为,植物残体中的碳水化合物等不稳定底物可以被土壤微生物迅速利用,并以微生物尸体的形式保留,而木质素酚等结构化合物则通过物理转移途径在土壤中积累。尽管存在化学抗性,但植物木质素在一定程度上可以被微生物吸收,并且在贫碳地块而不是富碳地块中发现了更大的木质素生物降解。因此,碳可用性随植物残基输入的变化调节了微生物底物利用策略,但目前尚不清楚微生物底物偏好如何影响微生物残基的产生和木质素苯酚在土壤中的保留。
正如一些长期田间定位实验中普遍发现的那样,作物秸秆还田可以增加微生物尸体堆积,而独立实验报告称,有机物质输入后也可以促进木质素苯酚的保留。当将它们的浓度归一化为SOC时,微生物死亡质量对SOC积累的相对贡献随着有机输入的增加而降低,这意味着微生物和植物来源的成分在促进SOC积累方面存在权衡。然而,目前尚不清楚作物残茬投入是如何影响这种效应的。特别是,与SOC池的可用性和稳定性相关的微生物和植物衍生成分的动态仍然不确定。
在此,基于保护性耕作下正在进行的长期玉米秸秆覆盖田间试验,通过分析氨基糖和木质素酚浓度,我们旨在量化微生物和植物来源成分的动态,并阐明它们在长期玉米秸秆还田影响下对有机碳的固存和稳定中的不同作用。我们假设:(1)添加玉米渣后,微生物尸体在早期迅速积累;(2)由于微生物对不稳定C的偏好,重复添加作物渣可以显著增强土壤中植物木质素的保留;(3)玉米渣归还后SOC积累的改善主要归因于新鲜植物来源的成分保留,这可以长期保持SOC的可分解性。
—
结果
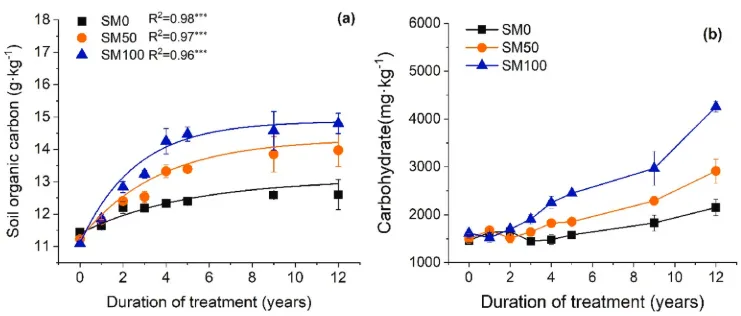
图1. (a)土壤有机碳(SOC)的时间动态;(b) 2007-2019年期间的土壤碳水化合物(中性糖)(n=3,误差条=SE)。星号表示单指数回归的显著性水平:*P<0.05、**P<0.01和***P<0.001。SM0,0%秸秆覆盖;SM50,50%秸秆覆盖和SM100,100%秸秆覆盖。
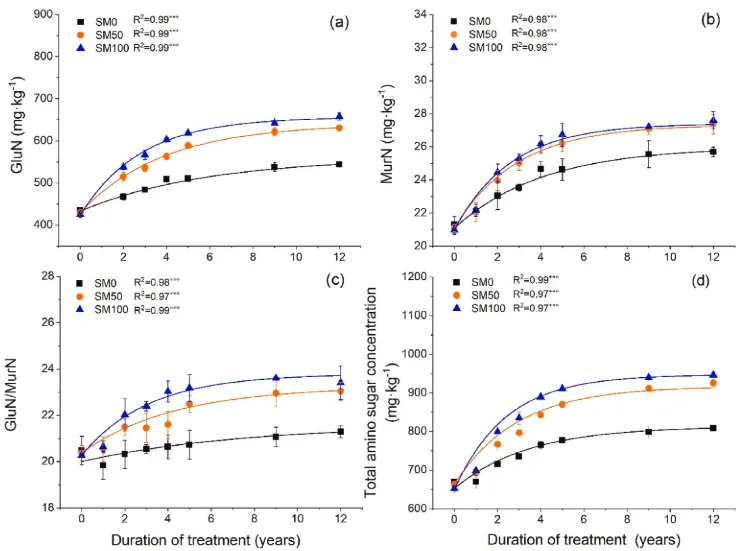
图2.(a)葡萄糖胺浓度的时间动态;(b) 胞壁酸浓度;(c) 葡萄糖胺与胞壁酸的比例(GluN/MurN);以及(d)2007-2019年期间的总氨基糖浓度(n=3,误差条=SE)。星号表示单指数回归的显著性水平:*P<0.05、**P<0.01和***P<0.001。SM0,0%秸秆覆盖;SM50,50%秸秆覆盖和SM100,100%秸秆覆盖。
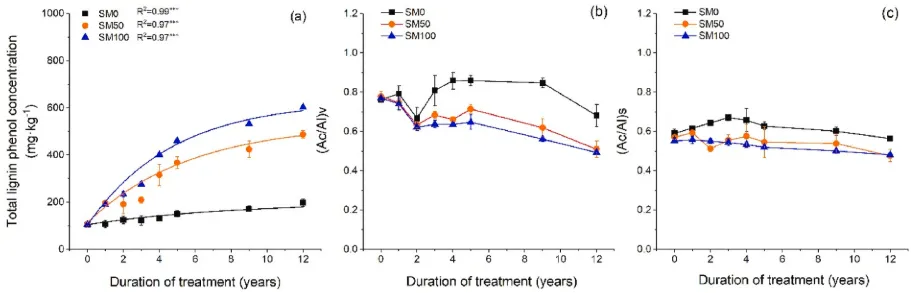
图3. (a)木质素酚总浓度的时间动态;(b) (Ac/Al)V和(c)(Ac/Al)S在2007年至2019年期间的变化(n=3,误差条=SE)。星号表示单指数回归的显著性水平:*P<0.05、**P<0.01和***P<0.001。SM0,0%秸秆覆盖;SM50,50%秸秆覆盖和SM100,100%秸秆覆盖。
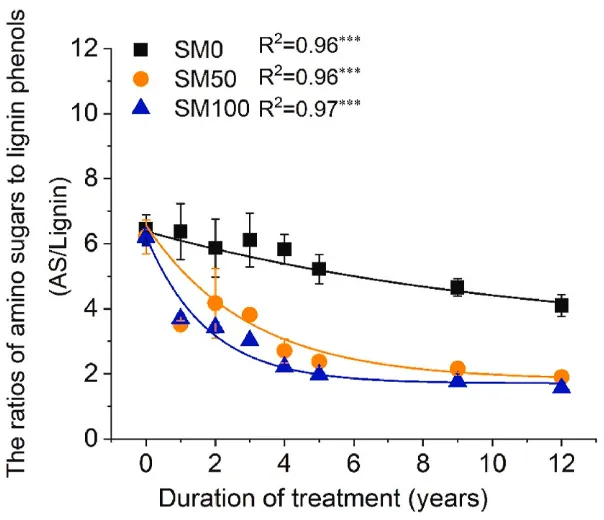
图4. 2007-2019年氨基糖与木质素酚比率的时间动态(n=3,误差条=SE)。星号表示单指数回归的显著性水平:*P<0.05、**P<0.01和***P<0.001。SM0,0%秸秆覆盖;SM50,50%秸秆覆盖和SM100,100%秸秆覆盖。
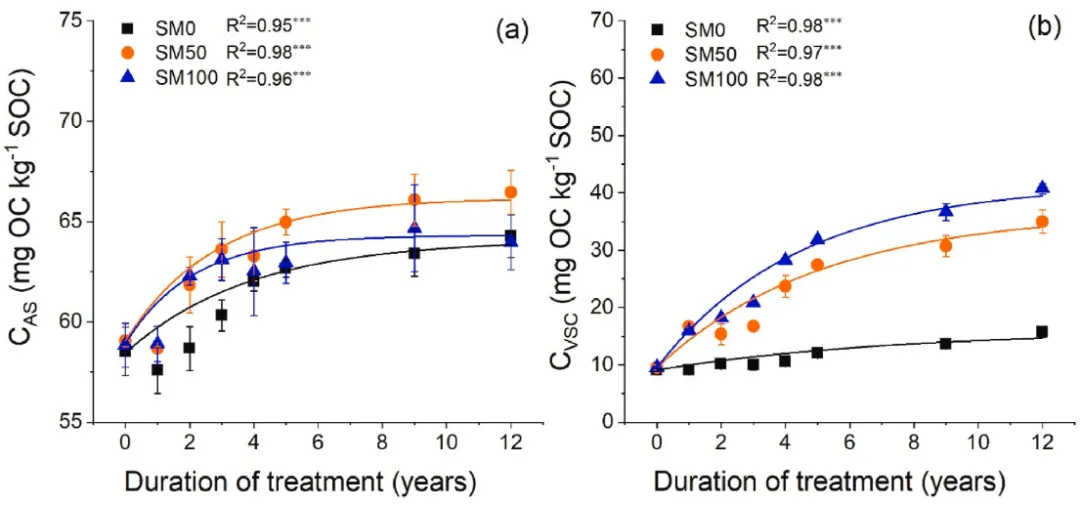
图5. 2007-2019年间(a)SOC中氨基糖的比例(CAS)和(b)SOC中木质素酚的比例(CVSC)的时间动态(n=3,误差条=SE)。星号表示单指数回归的显著性水平:*P<0.05、**P<0.01和***P<0.001。SM0,0%秸秆覆盖;SM50,50%秸秆覆盖和SM100,100%秸秆覆盖。
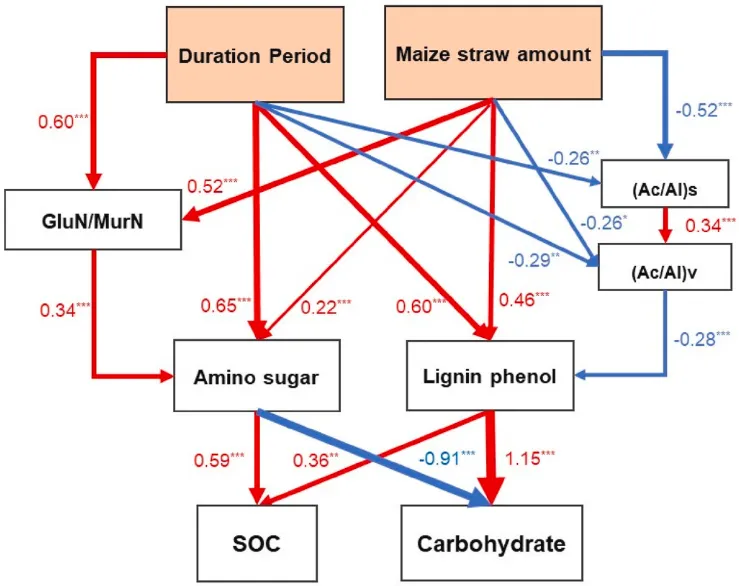
图6. 不同秸秆覆盖时间和数量下SOC组成来源(植物和微生物来源的成分)与SOC和碳水化合物含量之间假设因果关系的结构方程模型(SEM)。模型拟合:χ2=5.625,P=0.897,CFI=1.000,GFI=0.983,RMSEA=0.000。标准化路径系数(r,显著效应系数)列在每条路径旁边(红线表示正路径,蓝线表示负路径;线宽表示因子贡献的比例)。显著性影响:*P<0.05,**P<0.01,***P<0.001。
05
—
结论
根据不同的微生物和植物来源生物标志物的指示,基于12年的保护性耕作实验,探索了微生物尸体和植物木质素在介导长期SOC积累中的不同作用。玉米秸秆的添加有利于微生物的增殖和SOC的封存,但经过十年的非线性增加,微生物的死质量接近“微生物承载能力”。相比之下,玉米残茬归还率较高时SOC积累的饱和延迟主要归因于顽固木质素样植物残体的选择性保留,导致微生物死亡质量对扩大的SOC库容量的贡献被稀释。因此,保护性耕作下可持续SOC库的建立受到微生物和植物来源成分功能分化的严格控制。玉米秸秆覆盖可能提高了微生物死亡质量对SOC稳定的贡献,而植物木质素的增加,伴随着氧化的减少,从长远来看,主要与SOC对微生物代谢的可分解性提高有关。
论文链接:https://linkinghub.elsevier.com/retrieve/pii/S0038071723000251