2.1 植物材料培养与生物信息学分析
为探究紫花苜蓿中R2R3-MYB转录因子 MsMYB17 的功能及其系统发育关系,本研究选用紫花苜蓿"中苜1号"品种作为实验材料,所有植株均在人工气候室中于25°C、长日照条件(16 h光照/8 h黑暗)下进行培养。在生物信息学分析方面,分别从TAIR数据库和Figshare数据库获取拟南芥126个R2R3-MYB蛋白序列及紫花苜蓿的R2R3-MYB蛋白序列,利用MEGA7软件结合ClustalW进行多序列比对,并通过最大似然法构建系统发育树,以明确 MsMYB17 的进化地位。此外,利用PlantCARE在线工具对目标基因启动子区域的顺式作用元件进行预测分析,为后续功能研究提供序列信息基础。
2.2 转基因材料构建与分子功能验证
为验证 MsMYB17 的生物学功能及其调控靶基因的分子机制,构建了过表达(OE)和RNAi干扰两类转基因紫花苜蓿材料。将 MsMYB17 的CDS克隆至pBI121载体用于过表达,将其200 bp片段克隆至pK7GWIWG2D (II)载体用于基因沉默,并对两月龄转基因植株进行株高、节间数、节间长度和茎直径等表型指标测定。在分子功能验证层面,通过将 MsMYB17-GFP融合蛋白与核标记mRFP-AHL22共表达于本氏烟草叶片,利用激光共聚焦显微镜确定其亚细胞定位;通过酵母双杂交系统,将 MsMYB17 全长及截短片段与GAL4 DNA结合域融合,在选择性培养基上检测其转录激活活性;通过双荧光素酶报告实验,将 MsDFR 和 MsANS 启动子与报告基因融合,验证 MsMYB17 对下游靶基因的转录激活作用;通过EMSA实验,利用生物素标记的含MBS基序探针与纯化的GST-MsMYB17蛋白进行体外结合反应,证实 MsMYB17 与靶基因启动子的直接结合能力。
2.3 转录组分析、蛋白互作检测与生化指标测定
为系统解析 MsMYB17 介导的基因调控网络及其蛋白稳定性调控机制,开展了转录组测序、蛋白互作检测及相关生化指标测定。RNA-seq分析采用两月龄WT和 MsMYB17-OE植株叶片,在Illumina NovaSeq 6000平台完成测序,以HISAT2比对参考基因组,以|log₂(fold-change)| ≥ 1且校正后 p 值 < 0.05为阈值筛选差异表达基因,并结合RT-qPCR对关键基因表达进行验证。在蛋白稳定性研究中,通过无细胞降解实验,将纯化的GST-MsMYB17融合蛋白与经高光或对照光照处理的紫花苜蓿幼苗总蛋白共孵育,利用免疫印迹检测蛋白降解动态;通过Co-IP实验,在本氏烟草中共表达MsMYB17-GFP和MsCOP1-Myc,利用GFP-Trap磁珠进行免疫沉淀,验证两者的蛋白互作关系。此外,采用1% HCl-甲醇法提取组织花青素,经氯仿去除叶绿素后,通过测定530 nm吸光值进行花青素含量的定量分析。所有实验均设置至少三次生物学重复,数据以平均值±标准差表示,组间差异采用Student's t检验进行统计分析(p < 0.05)。
3.1 MsMYB17是一个定位于细胞核的转录激活因子
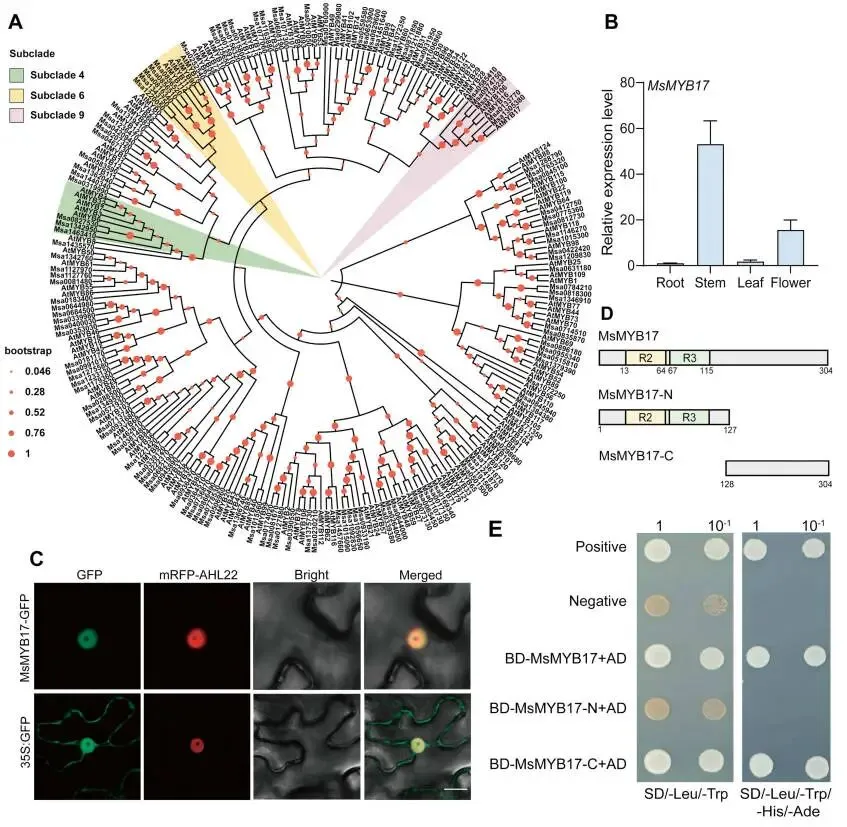
系统发育分析:基于前期研究发现第9亚群的MYB106可增强植物光合能力和生物量,本研究对紫花苜蓿基因组中的MYB蛋白进行鉴定和系统发育分析,筛选出与AtMYB17聚类且研究较少的Msa1305380,命名为 MsMYB17,其属于第9亚群并与MsMYB106亲缘关系较近(Fig.1A)。
序列与表达特征:MsMYB17 的开放阅读框(ORF)为915 bp,编码304个氨基酸;组织特异性表达分析显示,MsMYB17 在茎中表达量最高,在根中最低(Fig.1B)。
亚细胞定位:在本氏烟草叶片中瞬时表达MsMYB17-GFP融合蛋白,激光共聚焦显微镜观察结果表明MsMYB17定位于细胞核(Fig.1C)。
转录激活活性:将全长MsMYB17与GAL4 DNA结合域融合并与pGADT7共转化酵母,结果显示全长MsMYB17在缺乏Leu-Trp-His-Ade的SD培养基上生长良好,表明MsMYB17是转录激活因子;序列分析发现MsMYB17含有两个保守的MYB结构域(R2:13−64 aa;R3:67−115 aa)(Fig.1D);进一步将N端区域(1−127 aa)和C端区域(128−304 aa)分别与GAL4 DNA结合域融合进行转录激活活性检测,结果显示MsMYB17-C能在选择性培养基上生长而MsMYB17-N不能,表明MsMYB17的C端负责其转录激活功能(Fig.1E)。
本研究通过系统发育分析从紫花苜蓿中鉴定出属于第9亚群的R2R3-MYB转录因子 MsMYB17,其ORF为915 bp,编码304个氨基酸。组织特异性表达分析表明 MsMYB17 在茎中表达量最高,在根中最低。亚细胞定位实验证实MsMYB17定位于细胞核。酵母转录激活实验表明MsMYB17是一个转录激活因子,且其C端区域(128−304 aa)负责转录激活功能,而含有R2和R3保守结构域的N端区域(1−127 aa)不具有转录激活活性。
图1. MsMYB17的特征分析。(A)使用MEGA7.0软件通过最大似然法(1,000次bootstrap重复)构建的拟南芥和紫花苜蓿R2R3-MYB蛋白系统发育树。(B)MsMYB17 在根、茎、叶和花中的表达水平,以 Msactin 作为内参,数据以平均值±SD表示(n = 3)。(C)35S:MsMYB17-GFP和35S:GFP(对照)在烟草表皮细胞中的亚细胞定位,mRFP-AHL22作为细胞核标记,标尺 = 10 μm。(D)MsMYB17蛋白结构示意图。(E)MsMYB17全长及截短版本的自激活实验,SD/-Leu/-Trp为缺乏亮氨酸和色氨酸的SD培养基,SD/-Leu/-Trp/-His/-Ade为缺乏亮氨酸、色氨酸、组氨酸和腺嘌呤的SD培养基,BD-53 + AD-T作为阳性对照,BD-Lam + AD-T作为阴性对照。
3.2 过表达 MsMYB17 降低紫花苜蓿的株高和茎直径
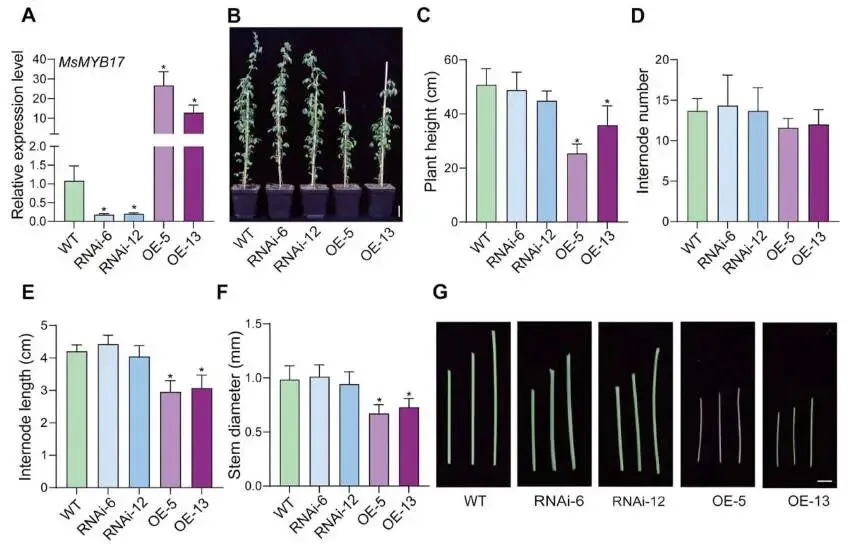
转基因材料获得:构建 MsMYB17 过表达(OE)和RNAi干扰转基因紫花苜蓿;RT-qPCR检测显示OE-5和OE-13株系中 MsMYB17 表达量分别为野生型(WT)的25倍和12倍,而RNAi-6和RNAi-12株系中表达量较WT降低约80%(Fig.2A)。
生长表型分析:MsMYB17-OE植株表现出矮化表型,株高较WT降低30%−50%,而 MsMYB17-RNAi植株与WT之间无明显生长表型差异(Fig.2B,C)。
节间特征测定:MsMYB17-OE与WT植株的节间数无显著差异,但 MsMYB17-OE的节间长度显著短于WT,表明株高降低源于节间缩短(Fig.2D,E)。
茎直径测定:MsMYB17-OE转基因紫花苜蓿的茎显著细于WT植株;WT与 MsMYB17-RNAi植株在株高、节间数、节间长度和茎直径方面均无显著差异(Fig.2F,G)。
为探究 MsMYB17 的功能,本研究构建了 MsMYB17 过表达和RNAi干扰转基因紫花苜蓿。RT-qPCR检测证实OE株系中 MsMYB17 表达量显著上调,RNAi株系中表达量显著下调。表型分析显示,MsMYB17-OE植株呈现矮化表型,株高较WT降低30%−50%,且茎直径显著变细,而节间数无变化,表明株高降低源于节间长度缩短。相比之下,MsMYB17-RNAi植株与WT在各项生长指标上均无显著差异。综上,过表达 MsMYB17 导致紫花苜蓿株高和茎直径降低。
图2. MsMYB17 转基因紫花苜蓿的表型。(A)转基因紫花苜蓿中 MsMYB17 的相对表达水平。(B)WT、MsMYB17-RNAi和 MsMYB17-OE植株的代表性图像,标尺 = 5 cm。(C, D, E, F)WT、MsMYB17-RNAi和 MsMYB17-OE紫花苜蓿的株高(C)、节间数(D)、节间长度(E)和茎直径(F),数据以平均值±SD表示(n = 3),星号表示经Student's t检验确定的显著性差异,p < 0.05。(G)WT、MsMYB17-RNAi和 MsMYB17-OE紫花苜蓿茎的代表性图像,标尺 = 5 mm。
3.3 MsMYB17通过激活 MsDFR 和 MsANS 促进花青素积累
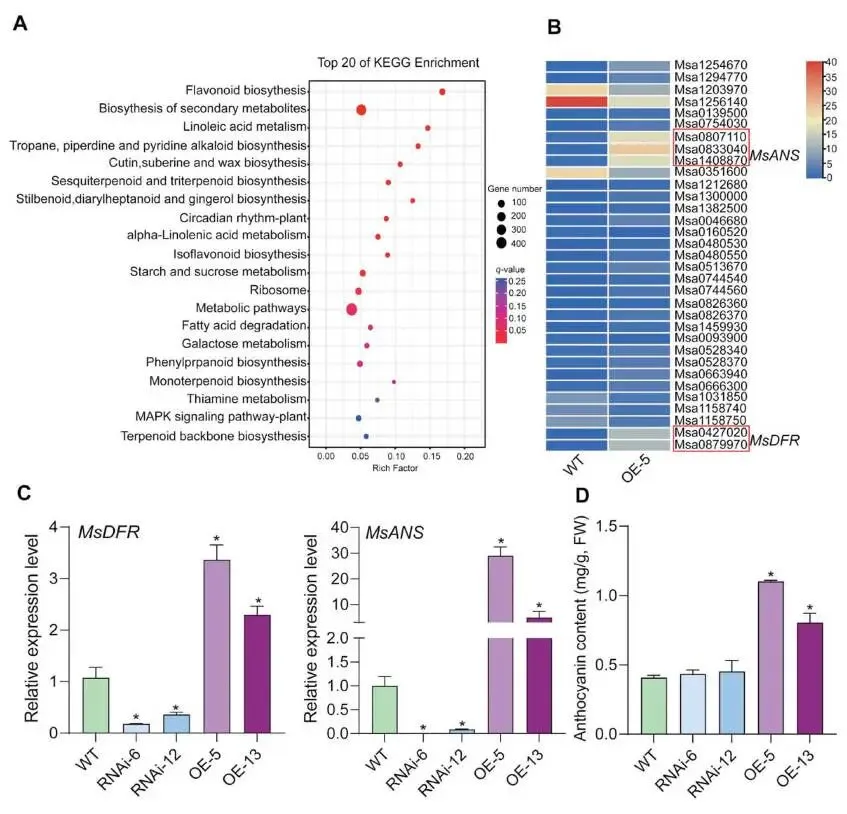
转录组分析:对WT和 MsMYB17-OE转基因紫花苜蓿进行RNA-seq分析,共鉴定出4482个差异表达基因(DEGs),其中3144个下调,1338个上调;KEGG分析显示"类黄酮生物合成"、"次生代谢物生物合成"和"亚油酸代谢"等通路显著富集(Fig.3A)。
关键基因鉴定:在富集程度最高的"类黄酮生物合成"通路中,MsDFR 和 MsANS 的表达水平在WT和 MsMYB17-OE植株间变化最为显著(Fig.3B)。
表达验证:RT-qPCR验证显示,与WT相比,MsDFR 和 MsANS 在 MsMYB17-OE植株中表达显著上调,在 MsMYB17-RNAi植株中显著下调(Fig.3C)。
花青素含量测定:MsMYB17-OE植株中花青素含量显著高于WT,而 MsMYB17-RNAi植株中花青素含量无显著变化(Fig.3D)。
其他基因分析:过表达 MsMYB17 导致 MsCHS、MsCHI 和 MsF3H 转录水平显著上调;MsMYB17-RNAi植株中与 MsMYB17 亲缘关系最近的 Msa0133770 表达显著上调,可能补偿了 MsMYB17 的下调。
为解析MsMYB17调控植物生长的机制,本研究对WT和 MsMYB17-OE转基因紫花苜蓿进行了RNA-seq分析。共鉴定出4482个DEGs,KEGG富集分析显示"类黄酮生物合成"通路显著富集,其中 MsDFR 和 MsANS 表达变化最为显著。RT-qPCR验证表明,MsDFR 和 MsANS 在 MsMYB17-OE植株中显著上调,在RNAi植株中显著下调。由于DFR和ANS催化花青素生成的连续步骤,花青素含量测定显示 MsMYB17-OE植株中花青素显著高于WT,而RNAi植株无变化。此外,MsMYB17-RNAi植株中 Msa0133770 表达上调可能对 MsMYB17 下调起补偿作用,这解释了RNAi植株花青素含量未变化的原因。
图3. MsMYB17促进紫花苜蓿花青素积累。(A)与WT相比,MsMYB17-OE植株中DEGs的前20个富集KEGG通路。(B)(A)图中"类黄酮生物合成"通路DEGs的热图。(C)WT、MsMYB17-RNAi和 MsMYB17-OE紫花苜蓿中 MsDFR 和 MsANS 的相对表达水平,数据以平均值±SD表示(n = 3)。(D)两月龄WT、MsMYB17-RNAi和 MsMYB17-OE紫花苜蓿叶片中的花青素含量,数据以平均值±SD表示(n = 3),(C)和(D)图中星号表示经Student's t检验确定的显著性差异,p < 0.05。
3.4 MsMYB17结合 MsDFR 和 MsANS 的启动子
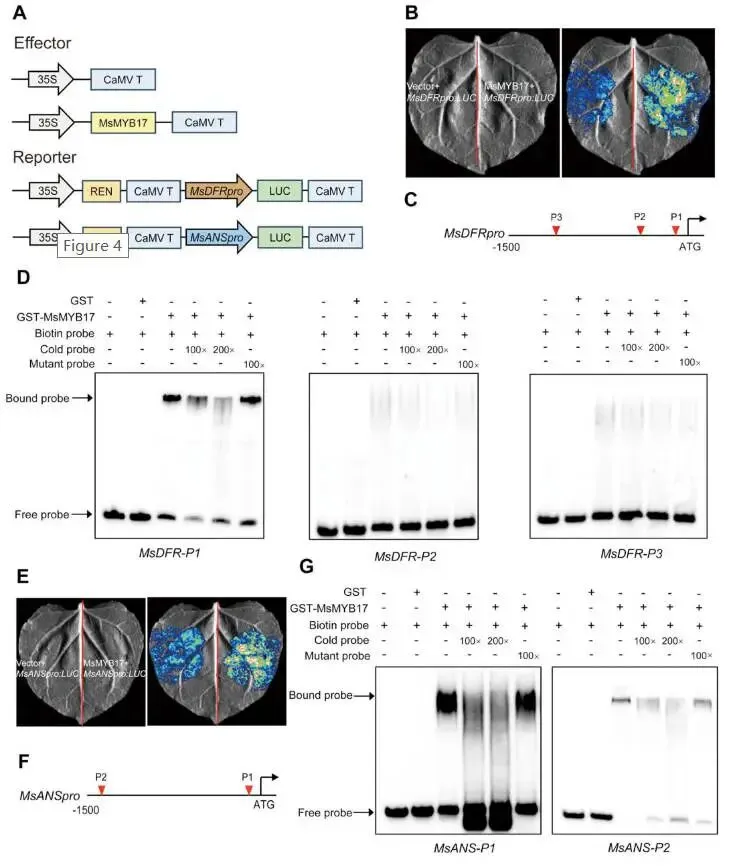
双荧光素酶实验:将 MsDFR 和 MsANS 启动子分别与LUC报告基因融合,与MsMYB17效应载体共注射烟草叶片;结果显示MsMYB17与 MsDFR 启动子共注射的LUC活性显著高于空载体对照,表明MsMYB17激活 MsDFR 表达(Fig.4A,B);同样,MsMYB17也激活 MsANS 的表达(Fig.4E)。
启动子元件分析:利用PlantCARE分析 MsDFR 启动子序列,鉴定出三个MYB结合顺式元件(MBS)(Fig.4C);MsANS 启动子中鉴定出两个MBS(Fig.4F)。
EMSA验证:设计各MBS元件的探针与纯化的GST-MsMYB17重组蛋白进行EMSA实验;结果表明MsMYB17蛋白直接结合 MsDFR-P1位点,而不结合 MsDFR-P2或 MsDFR-P3(Fig.4D);MsMYB17直接结合 MsANS-P1和 MsANS-P2两个位点(Fig.4G)。
为探究MsMYB17是否直接激活 MsDFR 和 MsANS 的表达,本研究进行了双荧光素酶报告实验和EMSA分析。双荧光素酶实验表明MsMYB17能够激活 MsDFR 和 MsANS 的表达。启动子序列分析在 MsDFR 启动子中鉴定出三个MBS元件,在 MsANS 启动子中鉴定出两个MBS元件。EMSA实验进一步证实MsMYB17蛋白直接结合 MsDFR 启动子的P1位点以及 MsANS 启动子的P1和P2位点。综上,MsMYB17通过直接结合 MsDFR 和 MsANS 启动子并激活其表达来促进花青素生物合成。
图4. MsMYB17结合并激活 MsDFR 和 MsANS 的表达。(A)双荧光素酶实验所用载体示意图。(B)和(E)双荧光素酶实验分别显示MsMYB17对 MsDFR 和 MsANS 的激活作用。(C)和(F)分别为 MsDFR 和 MsANS 启动子中MYB结合顺式元件的示意图。(D)和(G)EMSA实验分别显示MsMYB17结合 MsDFR 启动子的P1位点以及 MsANS 启动子的P1和P2位点,"+"和"-"分别表示所示探针或蛋白的存在和缺失。
3.5 MsMYB17-OE植株中木质素含量降低
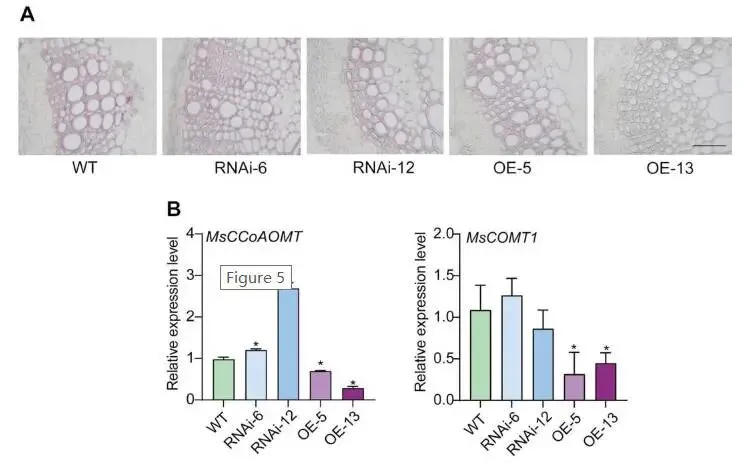
组织化学染色:采用Wiesner染色(间苯三酚-HCl)检测WT、MsMYB17-OE和 MsMYB17-RNAi植株的木质素积累;组织化学分析显示 MsMYB17-OE植株的木质化程度显著低于WT,而WT与 MsMYB17-RNAi植株之间无差异(Fig.5A)。
基因表达验证:RT-qPCR检测关键木质素生物合成基因 MsCCoAOMT 和 MsCOMT1 的表达;结果证实 MsMYB17-OE植株中这两个基因的表达显著低于WT,而WT与 MsMYB17-RNAi植株之间无差异(Fig.5B)。
鉴于花青素和木质素共享类黄酮代谢途径的共同底物,且 MsMYB17-OE紫花苜蓿表现出茎细和矮化表型,本研究检测了木质素积累情况。Wiesner组织化学染色显示 MsMYB17-OE植株的木质化程度显著降低。RT-qPCR证实关键木质素生物合成基因 MsCCoAOMT 和 MsCOMT1 在 MsMYB17-OE植株中表达显著下调,而RNAi植株与WT无差异。因此,木质素含量降低是 MsMYB17-OE紫花苜蓿矮化表型的原因。
图5. 过表达 MsMYB17 降低紫花苜蓿木质素含量。(A)使用间苯三酚-HCl对WT、MsMYB17-RNAi和 MsMYB17-OE紫花苜蓿茎横切面进行组织化学染色,标尺 = 50 μm。(B)WT、MsMYB17-RNAi和 MsMYB17-OE紫花苜蓿中 MsCCoAOMT 和 MsCOMT1 的相对表达水平,数据以平均值±SD表示(n = 3),星号表示经Student's t检验确定的显著性差异,p < 0.05。
3.6 高光通过激活 MsDFR 和 MsANS 表达诱导紫花苜蓿花青素生物合成
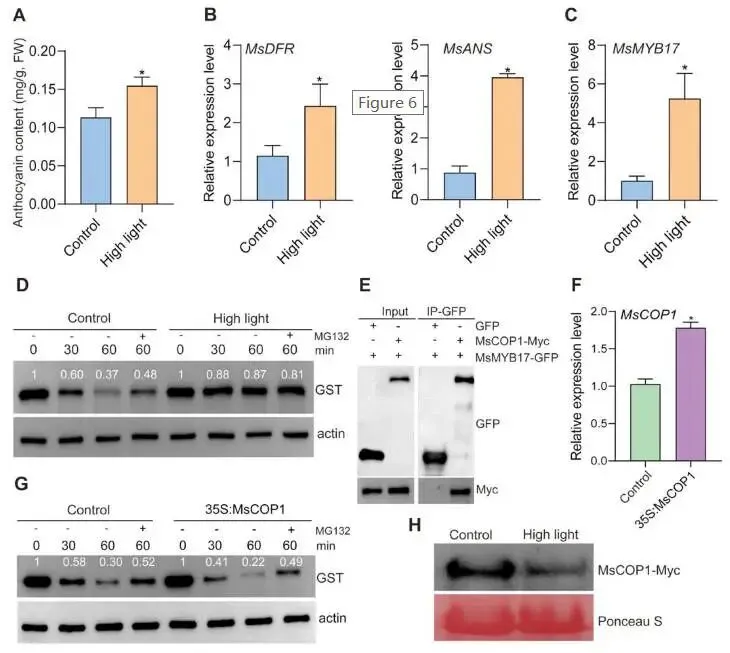
花青素含量测定:检测正常生长光照(对照)和高光处理紫花苜蓿的花青素含量;结果显示高光处理显著增加了紫花苜蓿的花青素含量(Fig.6A)。
基因表达分析:高光条件下 MsDFR 和 MsANS 的表达水平也显著上调,表明这两个基因负责紫花苜蓿高光条件下的花青素生物合成(Fig.6B)。
前期研究表明高光条件下花青素会积累,本研究检测了高光是否也诱导紫花苜蓿花青素积累。结果显示,与对照相比,高光处理显著增加了紫花苜蓿的花青素含量。同时,MsDFR 和 MsANS 的表达水平在高光条件下也显著上调,表明这两个基因负责紫花苜蓿高光条件下的花青素生物合成。
3.7 高光诱导 MsMYB17 表达并通过26S蛋白酶体途径抑制MsCOP1介导的MsMYB17降解
表达水平检测:检测对照和高光条件下 MsMYB17 的表达;结果显示高光条件下 MsMYB17 表达水平显著上调(Fig.6C)。
蛋白降解分析:无细胞降解实验显示,在对照条件下MsMYB17蛋白随孵育时间延长快速降解,添加26S蛋白酶体抑制剂MG132可阻断降解;而在高光条件下MsMYB17几乎不降解,添加MG132对蛋白丰度无显著影响(Fig.6D)。
蛋白互作验证:Co-IP实验显示MsCOP1-Myc能与MsMYB17-GFP共沉淀,而不能与单独的GFP共沉淀,表明MsCOP1与MsMYB17存在相互作用(Fig.6E)。
MsCOP1功能验证:在紫花苜蓿毛状根中过表达 MsCOP1(Fig.6F),无细胞降解实验显示 MsCOP1 过表达植株中MsMYB17降解速率加快,MG132可有效阻断降解,表明MsCOP1通过26S蛋白酶体途径介导MsMYB17降解(Fig.6G)。
高光对MsCOP1的影响:Western blot检测显示,与对照条件相比,高光显著降低了MsCOP1蛋白丰度(Fig.6H)。
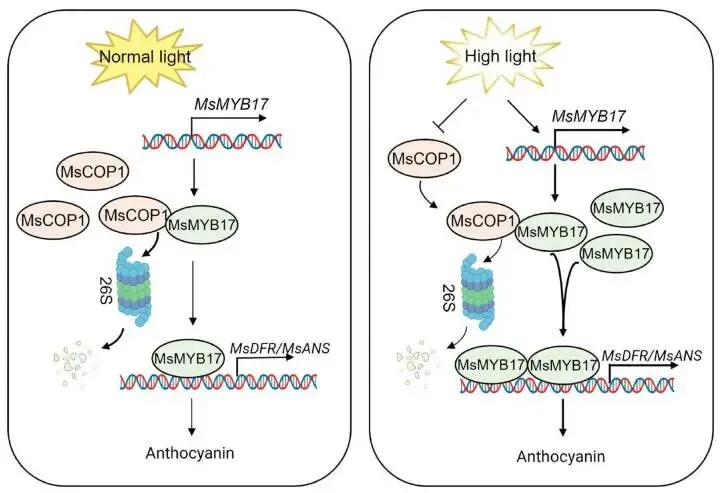
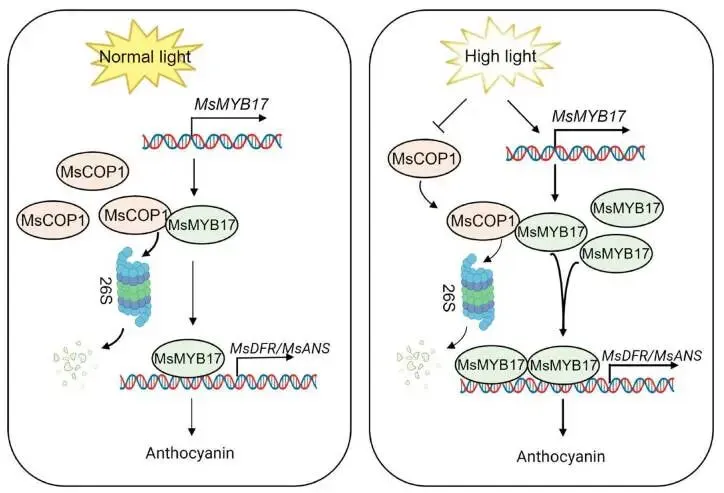
为探究 MsMYB17 是否参与高光诱导的花青素积累,本研究检测了对照和高光条件下 MsMYB17 的表达及蛋白稳定性。结果显示高光显著上调 MsMYB17 表达水平。无细胞降解实验表明,对照条件下MsMYB17蛋白通过26S蛋白酶体途径快速降解,而高光条件下降解被抑制。Co-IP实验证实MsCOP1与MsMYB17存在蛋白互作。在毛状根中过表达 MsCOP1 加速了MsMYB17的降解,且该降解可被MG132阻断,表明MsCOP1通过26S蛋白酶体途径介导MsMYB17降解。此外,高光显著降低MsCOP1蛋白丰度。这些发现提供了高光通过26S蛋白酶体途径抑制MsCOP1介导的MsMYB17降解的直接证据。综上,高光激活 MsMYB17 表达并抑制MsCOP1介导的MsMYB17降解,积累的MsMYB17直接结合 MsDFR 和 MsANS 启动子以激活其表达和花青素生成(Fig.7)。
图6. 高光通过MsCOP1-MsMYB17-MsDFR/MsANS模块促进紫花苜蓿花青素生成。(A)高光和正常光照(对照)处理紫花苜蓿的花青素含量。(B)和(C)高光和对照条件下 MsDFR、MsANS 和 MsMYB17 的相对表达水平,数据以平均值±SD表示(n = 3),星号表示经Student's t检验确定的显著性差异,p < 0.05。(D)无细胞降解实验显示与对照相比高光条件下MsMYB17降解速率降低,Actin作为上样对照。(E)Co-IP实验显示MsCOP1与MsMYB17的相互作用。(F)过表达 MsCOP1 的紫花苜蓿毛状根中 MsCOP1 的相对表达水平,数据以平均值±SD表示(n = 3),星号表示经Student's t检验确定的显著性差异,p < 0.05。(G)无细胞降解实验显示与对照相比过表达 MsCOP1 的紫花苜蓿毛状根中GST-MsMYB17蛋白降解速率加快,Actin作为上样对照。(H)对照和高光条件下MsCOP1蛋白水平的Western blot分析,丽春红S染色作为上样对照。
图7. 高光通过MsCOP1-MsMYB17-MsDFR/MsANS模块诱导花青素生物合成的模型。 在正常光照条件下(左侧),MsCOP1蛋白积累并促进MsMYB17的降解,低水平的残余MsMYB17蛋白维持花青素生物合成结构基因 MsDFR 和 MsANS 的基础表达,导致有限的花青素生成。在高光条件下(右侧),发生两个关键事件:(1)MsMYB17 表达的转录激活,以及(2)MsCOP1介导的MsMYB17降解被抑制,二者共同导致MsMYB17蛋白积累。MsMYB17随后直接结合 MsDFR 和 MsANS 的启动子,激活其表达并增强花青素生物合成。
首次发现第9亚群R2R3-MYB转录因子正向调控花青素合成。传统研究普遍认为,R2R3-MYB家族中第6亚群成员促进花青素合成,而第4亚群成员则起抑制作用。本研究突破了这一认知,首次鉴定并证实了属于第9亚群的转录因子 MsMYB17 是紫花苜蓿花青素生物合成的正向调控因子,拓展了对MYB转录因子家族功能多样性的理解。
首次解析了紫花苜蓿中花青素合成的MYB调控通路。尽管花青素对紫花苜蓿的品质和抗逆性至关重要,但其合成调控机制此前一直未被报道。本研究填补了这一空白,明确了 MsMYB17 通过直接结合下游关键基因 MsDFR 和 MsANS 的启动子,并激活其表达,从而促进花青素积累的分子通路,为紫花苜蓿这一重要牧草的品质改良提供了新的理论依据和基因靶点。
创新性地构建了高光诱导花青素合成的“MsCOP1-MsMYB17-MsDFR/MsANS”调控模块。本研究系统地揭示了一个全新的、整合了转录和转录后调控的高光响应分子模块。该模块阐明了高光通过双重机制调控 MsMYB17:一方面在转录水平上调其表达;另一方面在转录后水平通过抑制E3泛素连接酶MsCOP1的活性,从而稳定MsMYB17蛋白。这一完整调控链的发现,为理解植物如何精确响应光信号、调控次生代谢产物积累提供了新的范式。
揭示了MsCOP1通过26S蛋白酶体途径介导MsMYB17降解的新机制。研究通过免疫共沉淀(Co-IP)和无细胞降解实验,首次证实了MsCOP1是调控MsMYB17蛋白稳定性的关键E3泛素连接酶。更重要的是,研究发现高光通过降低MsCOP1蛋白的丰度来抑制其对MsMYB17的降解作用。这不仅拓展了对COP1功能的新认知,也深化了对光信号如何通过蛋白降解途径调控转录因子活性的理解。
阐明了MsMYB17调控下花青素与木质素合成的代谢权衡关系。研究发现,过表达 MsMYB17 在促进花青素积累的同时,显著抑制了木质素的合成,并导致植株矮化和茎秆变细。这表明 MsMYB17 在调控代谢流向、即在花青素与木质素两条分支途径之间扮演了“开关”角色,为理解次生代谢网络调控及植物表型建成的分子基础提供了直接证据。
发现了第9亚群MYB蛋白独特的结构与功能特征。通过序列分析,本研究指出 MsMYB17 虽与第6亚群激活子同样拥有保守的R2R3结构域,但其缺乏第6亚群特有的激活基序(KPRPR[S/T]),而是拥有一个不同的保守酸性基序。这一结构上的差异可能导致其通过不同的蛋白互作方式或辅因子招募机制发挥功能,为研究MYB转录因子结构与功能多样性提供了新的线索。
1. MsMYB17 的功能特性:
2. 调控花色苷和木质素合成:
3. 高光对 MsMYB17 的双重调控机制:
审核|林蘖亙
阅读科研文献,荟萃前沿进展
欢迎关注、转发、投稿、点赞





 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
