
 论文摘要先前研究表明,梨黑斑病菌(Alternaria alternata)能够利用添加了外源酚酸单体(浓度与梨果皮中酚酸浓度相当)的培养基作为唯一碳源,并以物种特异性和浓度依赖性方式降解这些化合物。然而,梨果皮中酚酸降解的详细分子机制仍未完全明确。转录组分析显示,在 1 mM 外源绿原酸和阿魏酸处理后,梨黑斑病菌中与芳香族化合物代谢相关的基因显著上调。儿茶酚 1,2 - 双加氧酶(AaCHD)作为酚类代谢中的关键酶,通过靶向基因敲除技术进行进一步研究。尽管 AaCHD 突变体与野生型菌株在生长、孢子萌发或致病性方面未观察到显著差异,但 AaCHD1 和 AaCHD2 基因的缺失导致病菌对外源酚酸和渗透胁迫的敏感性增加。此外,超高效液相色谱分析表明,与外源酚酸孵育 48 小时后,ΔAaCHD1 突变体仍保持较高的绿原酸残留量,而 ΔAaCHD1 和 ΔAaCHD2 突变体的对香豆酸残留量均高于野生型。这些发现表明,尽管 AaCHD 酶对梨黑斑病菌的生长发育并非必需,但它可能在该病菌降解梨果皮中的绿原酸和对香豆酸过程中发挥关键作用。
论文摘要先前研究表明,梨黑斑病菌(Alternaria alternata)能够利用添加了外源酚酸单体(浓度与梨果皮中酚酸浓度相当)的培养基作为唯一碳源,并以物种特异性和浓度依赖性方式降解这些化合物。然而,梨果皮中酚酸降解的详细分子机制仍未完全明确。转录组分析显示,在 1 mM 外源绿原酸和阿魏酸处理后,梨黑斑病菌中与芳香族化合物代谢相关的基因显著上调。儿茶酚 1,2 - 双加氧酶(AaCHD)作为酚类代谢中的关键酶,通过靶向基因敲除技术进行进一步研究。尽管 AaCHD 突变体与野生型菌株在生长、孢子萌发或致病性方面未观察到显著差异,但 AaCHD1 和 AaCHD2 基因的缺失导致病菌对外源酚酸和渗透胁迫的敏感性增加。此外,超高效液相色谱分析表明,与外源酚酸孵育 48 小时后,ΔAaCHD1 突变体仍保持较高的绿原酸残留量,而 ΔAaCHD1 和 ΔAaCHD2 突变体的对香豆酸残留量均高于野生型。这些发现表明,尽管 AaCHD 酶对梨黑斑病菌的生长发育并非必需,但它可能在该病菌降解梨果皮中的绿原酸和对香豆酸过程中发挥关键作用。 研究背景
研究背景梨黑斑病菌(Alternaria alternata)是引发梨黑斑病的致病菌,可导致梨树叶斑、腐烂和枯萎,造成严重的采前采后产量与品质损失,该病菌兼具内生菌和强致病菌特性,能在梨生长阶段潜伏,待采后果实代谢变化、防御化合物减少时重新激活;为抵御该病菌,梨植株会通过产生酚类化合物构建防御体系,梨果皮中富含绿原酸、咖啡酸、对香豆酸、阿魏酸等酚酸类物质,这些物质凭借抗氧化和抗菌特性,可通过抑制真菌孢子萌发、菌丝生长、干扰代谢关键酶活性或强化细胞壁等方式抵御病菌侵染。然而,众多真菌病原体已进化出规避植物酚类防御的机制,主要包括通过氧化还原酶、水解酶等进行酶促降解,或借助 ABC 转运蛋白、MFS 转运蛋白等进行主动运输排出酚类化合物,其中儿茶酚 1,2 - 双加氧酶(CHD)作为酚类代谢关键酶,已在黑曲霉、异旋孢腔菌等丝状真菌中被证实可通过催化儿茶酚芳香环氧化裂解实现酚类解毒,助力真菌侵染宿主,但目前尚无直接证据表明梨黑斑病菌中的 CHD 类酶参与梨果皮酚酸防御的中和过程,其相关分子机制仍有待阐明,因此开展本研究以填补这一空白。
 图文赏析
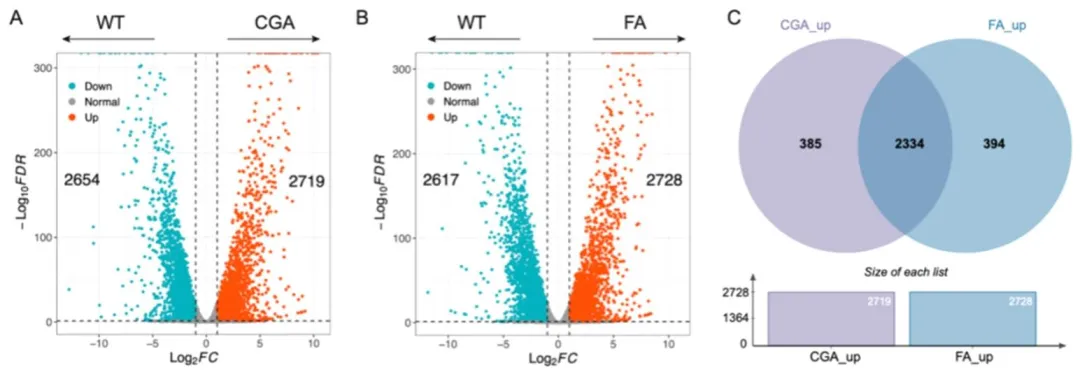
图文赏析图 1(Fig. 1):(A)绿原酸(CGA)处理组、(B)阿魏酸(FA)处理组差异表达基因(DEGs)的火山图(X 轴表示 log₂转换后的折叠变化值,Y 轴表示 - log₁₀转换后的假发现率(FDR)值;红色代表上调基因,蓝色代表下调基因,黑色代表组间无差异的基因);(C)维恩图显示 CGA(紫色圆圈)和 FA(蓝色圆圈)处理组中上调基因的重叠情况(仅 CGA 处理组特有上调基因 385 个,仅 FA 处理组特有上调基因 394 个,两组共有的上调基因 2334 个;CGA 处理组相较于野生型(WT)的上调基因总数为 2719 个,FA 处理组相较于野生型的上调基因总数为 2728 个)
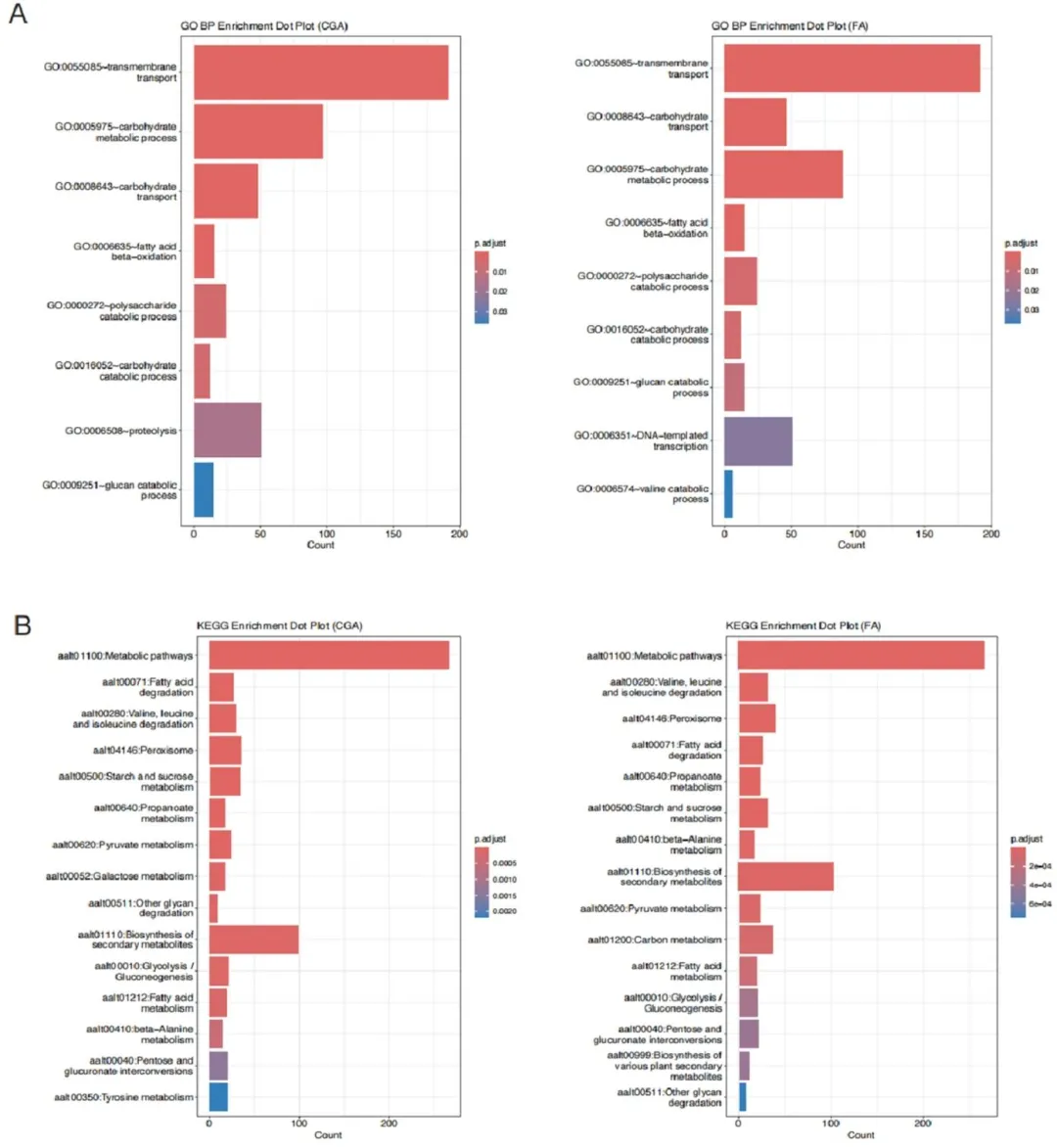
图 2(Fig. 2):(A)CGA 和 FA 处理组差异表达基因(DEGs)的基因本体(GO)生物学过程(BP)富集分析点图;(B)CGA 和 FA 处理组差异表达基因(DEGs)的京都基因与基因组百科全书(KEGG)通路富集分析点图
图 3(Fig. 3):17 个关键代谢基因、应激响应基因和转运蛋白基因在野生型(灰色)、CGA 处理组(红色)和 FA 处理组(蓝色)中的 log 转换表达量箱线图(每个图上方括号内分别标注 CGA 处理组与野生型、FA 处理组与野生型比较的 P 值)
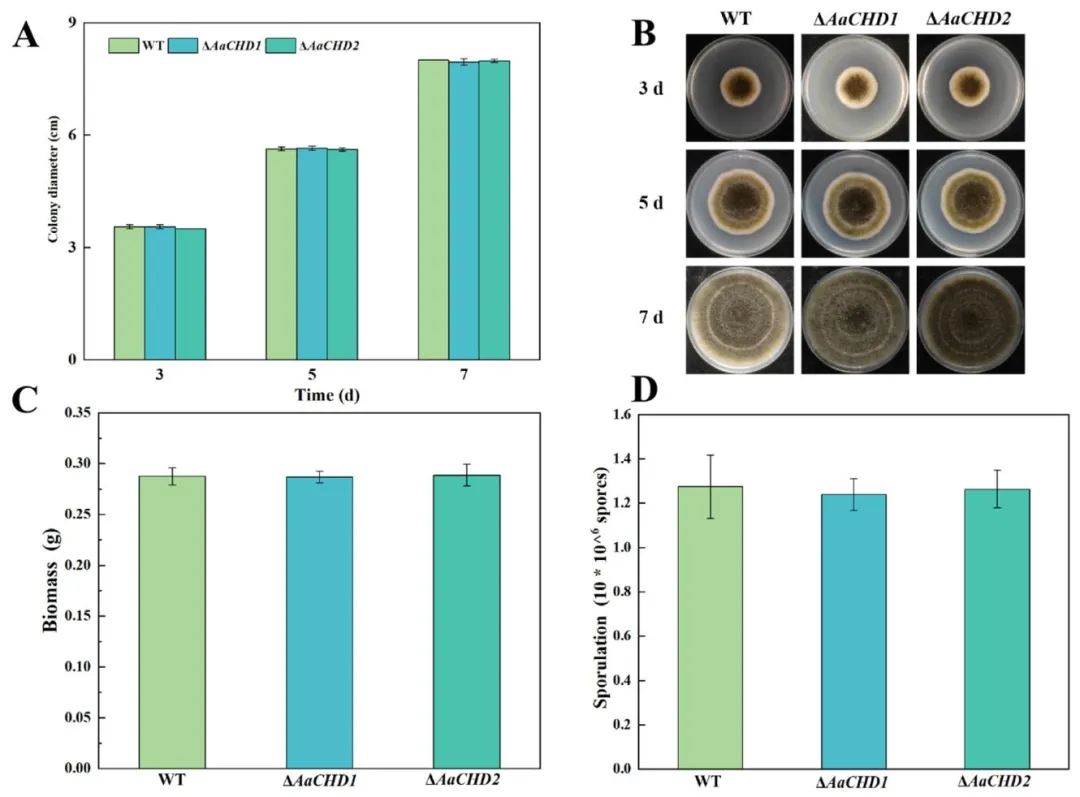
图 4(Fig. 4):AaCHD 基因对梨黑斑病菌(A. alternata)菌落形态(A)和菌落直径(B)的影响(误差线表示标准误 ±SE);AaCHD 基因对梨黑斑病菌生物量(C)和产孢量(D)的影响(误差线表示标准误 ±SE)
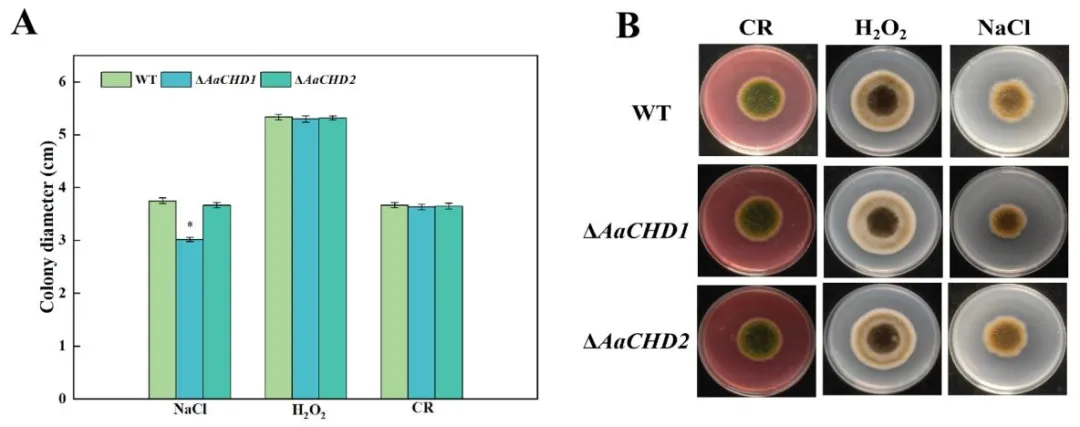
图 5(Fig. 5):不同外源胁迫下 AaCHD 基因对梨黑斑病菌菌落直径(A)和菌落形态(B)的影响(误差线表示标准误 ±SE;星号(*)表示差异显著(P<0.05);NaCl 模拟渗透胁迫,H₂O₂模拟氧化胁迫,刚果红(CR)干扰细胞壁完整性)
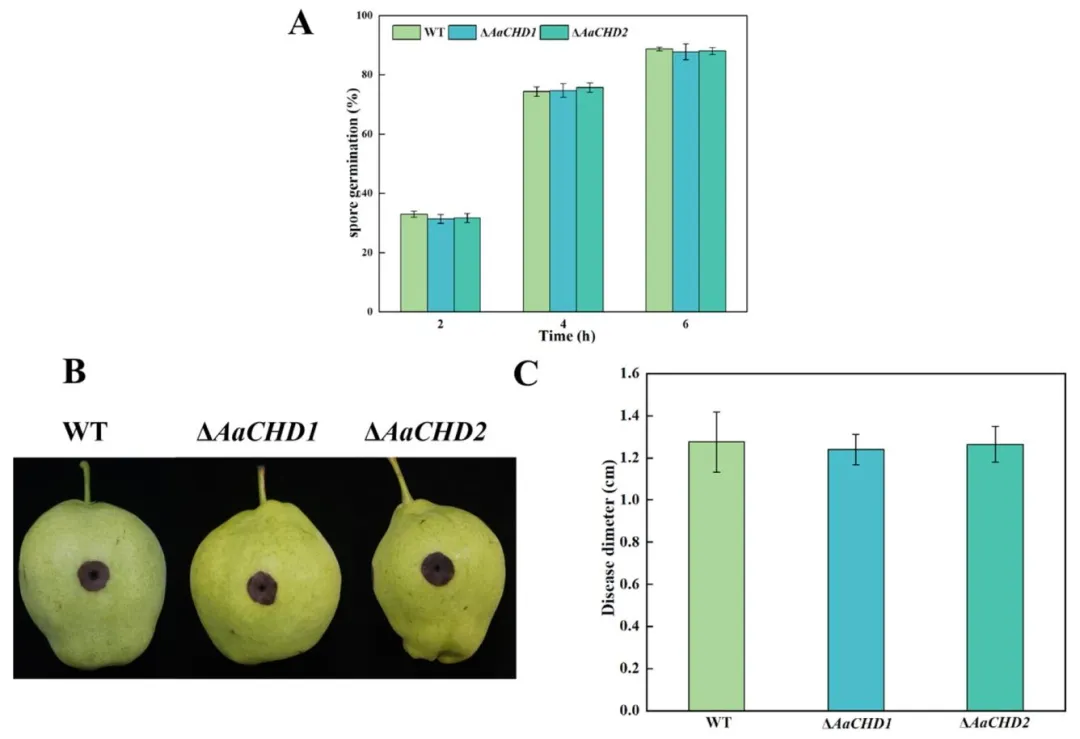
图 6(Fig. 6):AaCHD 基因对梨黑斑病菌孢子萌发(A)的影响(误差线表示标准误 ±SE);AaCHD 基因对梨果实病害扩展(B)和病斑直径(C)的影响(误差线表示标准误 ±SE)
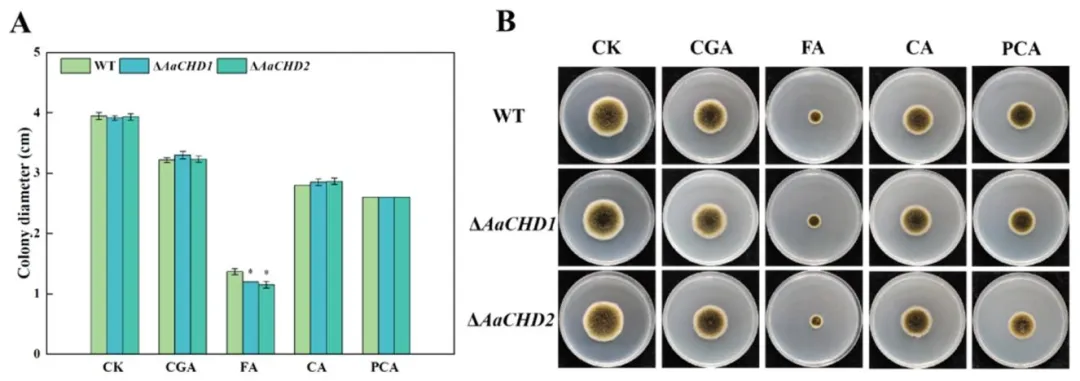 图 7(Fig. 7):外源酚酸对 AaCHD 基因敲除突变体(ΔAaCHDs)和野生型(WT)菌落直径(A)和菌落形态(B)的影响(误差线表示标准误 ±SE;星号(*)表示差异显著(P<0.05);CK 为对照组,CGA 为绿原酸,FA 为阿魏酸,CA 为咖啡酸,pCA 为对香豆酸)
图 7(Fig. 7):外源酚酸对 AaCHD 基因敲除突变体(ΔAaCHDs)和野生型(WT)菌落直径(A)和菌落形态(B)的影响(误差线表示标准误 ±SE;星号(*)表示差异显著(P<0.05);CK 为对照组,CGA 为绿原酸,FA 为阿魏酸,CA 为咖啡酸,pCA 为对香豆酸)
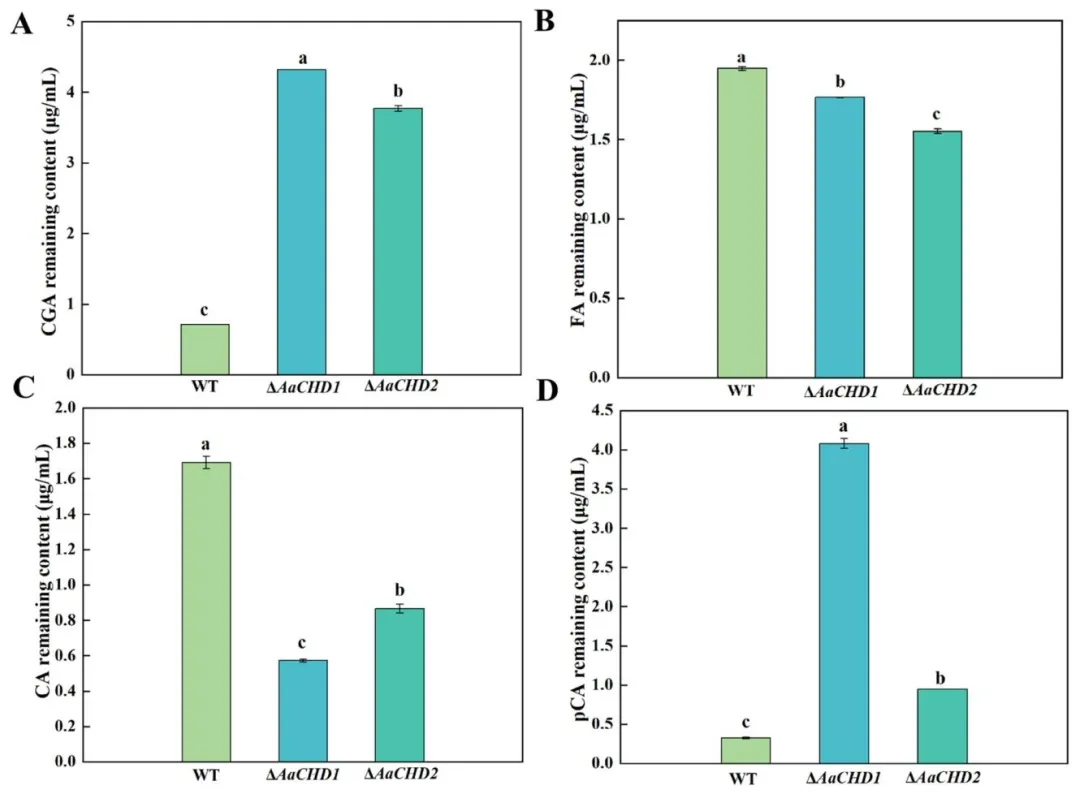
图 8(Fig. 8):AaCHD 基因对梨黑斑病菌降解绿原酸(A)、阿魏酸(B)、咖啡酸(C)和对香豆酸(D)的影响
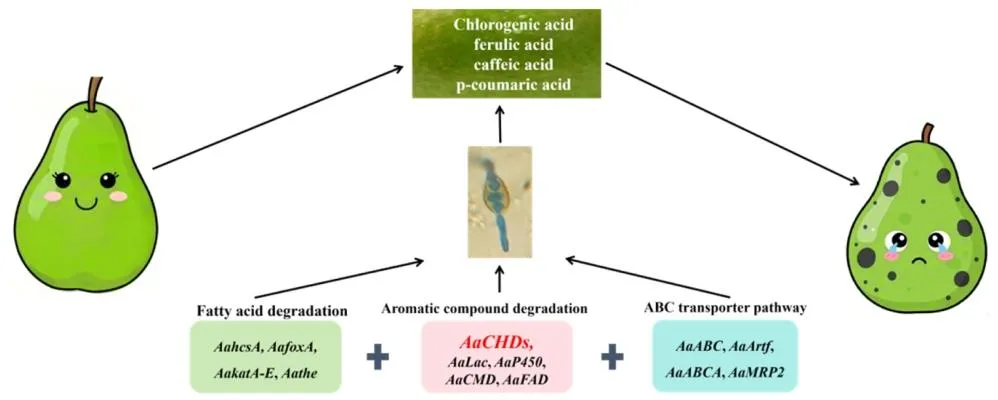
图 9(Fig. 9):梨黑斑病菌降解梨果皮酚酸类物质的功能及分子机制示意图(儿茶酚 1,2 - 双加氧酶是降解梨果皮酚酸类物质的关键途径之一,可有效降解绿原酸和对香豆酸,助力真菌完成定殖;涉及脂肪酸降解、芳香族化合物降解、ABC 转运蛋白通路等,相关基因包括 AaP450、AaKatA-E、AaABCA、AaMRP2、AaABC、AaArf 等)
 研究结论总之,转录组分析表明,梨黑斑病菌(A. alternata)在暴露于外源酚酸后,与芳香族化合物降解相关的基因显著上调。值得注意的是,我们鉴定出儿茶酚 1,2 - 双加氧酶(AaCHD)是参与该过程的关键差异表达基因。利用 AaCHD 基因敲除突变体进行的功能鉴定显示,尽管 AaCHD 对真菌的生长发育并非必需,但它在梨黑斑病菌降解绿原酸(CGA)和对香豆酸(pCA)过程中发挥着重要作用(图 9)。这些发现加深了我们对梨黑斑病菌解毒梨果皮酚酸分子机制的理解,为开发梨黑斑病的靶向防控策略提供了参考。李永才:甘肃农业大学食品科学与工程学院教授,博士生导师。研究领域:从事食品质量安全控制、风险评价及甘肃特色果蔬马铃薯、甜瓜、梨等的品质形成及采后品质劣变发生机制、采后侵染性病害发生规律及病原物与寄主互作机理、果蔬采后安全防腐保鲜技术等方面的研究工作。本研究得到国家自然科学基金(项目编号:32372411)和国家重点研发计划(项目编号:2021YFD2100502-3)的资助。https://doi.org/10.1016/j.postharvbio.2026.114194
研究结论总之,转录组分析表明,梨黑斑病菌(A. alternata)在暴露于外源酚酸后,与芳香族化合物降解相关的基因显著上调。值得注意的是,我们鉴定出儿茶酚 1,2 - 双加氧酶(AaCHD)是参与该过程的关键差异表达基因。利用 AaCHD 基因敲除突变体进行的功能鉴定显示,尽管 AaCHD 对真菌的生长发育并非必需,但它在梨黑斑病菌降解绿原酸(CGA)和对香豆酸(pCA)过程中发挥着重要作用(图 9)。这些发现加深了我们对梨黑斑病菌解毒梨果皮酚酸分子机制的理解,为开发梨黑斑病的靶向防控策略提供了参考。李永才:甘肃农业大学食品科学与工程学院教授,博士生导师。研究领域:从事食品质量安全控制、风险评价及甘肃特色果蔬马铃薯、甜瓜、梨等的品质形成及采后品质劣变发生机制、采后侵染性病害发生规律及病原物与寄主互作机理、果蔬采后安全防腐保鲜技术等方面的研究工作。本研究得到国家自然科学基金(项目编号:32372411)和国家重点研发计划(项目编号:2021YFD2100502-3)的资助。https://doi.org/10.1016/j.postharvbio.2026.114194
标注 “原创” 仅表明为原创编译,本平台不主张对原文享有版权。本平台转载内容仅出于学术交流与信息传播之目的,并不代表本平台观点,亦不意味着证实内容的真实性。转载文章的版权归原作者所有,若原作者不希望作品被转载或存在侵权行为,请联系本平台删除。由于编译水平有限,推文或简历中若有不妥之处,我们深表歉意,请联系本平台进行修改或删除。













 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
