2026 年 3 月 31 日,来自南京农业大学的Huibing Chi等人在Journal of Agricultural and Food Chemistry上发表了一篇题为Structure-Guided Design Unlocks the Coupled Improvement of Thermostability and Activity in Lipoxygenase的研究性论文。Abstract
脂氧合酶(LOX)广泛应用于食品和制药行业,但其工业应用受限于催化活性低和热稳定性差。本研究采用B因子引导的理性设计策略,对球状念珠藻(Nostoc sphaeroides)脂氧合酶(NsLOX)进行工程改造。通过定点饱和突变和组合突变,获得了多个热稳定性提高的突变体。其中,双突变体T2C/S36P在50 °C下的半衰期达到264.36 ± 21.66 min,是野生型酶的30.78倍;其熔解温度也从59.81 ± 0.17 °C提高至65.34 ± 1.74 °C,增加了5.53 °C。值得注意的是,T2C/S36P的比活性达到197832.16 ± 3860.89 U/mg,较野生型提高了1.53倍。分子动力学和结构分析表明,热稳定性的增强源于整体构象刚性的增加和表面性质的优化。在此基础上,通过底物结合口袋的重新设计——在保留关键静电相互作用的同时重塑疏水拓扑结构,实现静电-疏水平衡——协同提升了催化活性。据我们所知,这是首批通过B因子引导设计同时提高脂氧合酶热稳定性和活性的报道之一,为工业相关酶的工程改造提供了一种高效策略。
Introduction
脂氧合酶(EC 1.13.11.x,LOX)催化含有1,4-戊二烯结构的不饱和脂肪酸氧化为氢过氧化物。这些反应支撑了其在面粉强化与漂白、洗涤剂配方以及药物中间体合成等不同领域的应用。因此,LOX在食品、化工和制药行业中展现出巨大的应用潜力。该酶广泛存在于植物、动物和微生物中。目前,商品化LOX制剂主要来源于大豆提取物,但受限于季节波动、成分复杂且催化活性较低等因素,制约了其工业适用性。相比之下,微生物来源的LOX具有不受季节影响、活性高、成本低廉且易于规模化生产等优势,成为当前研究的热点。然而,微生物LOX的实际应用常受限于催化活性不足,尤其是热稳定性差的问题。低热稳定性和低活性不仅增加了加工过程中微生物污染的风险,也提高了生产成本,从而限制了其更广泛的利用。因此,开发热稳定性和活性更优的LOX变体对于其有效工业应用至关重要。
LOX的催化功能与其保守的结构特征和独特的自由基机制密切相关。尽管不同来源的LOX在序列上存在差异,但其催化核心结构高度保守。典型的LOX由两个主要结构域组成:直接参与催化的C端结构域和功能多样的N端结构域。C端催化结构域包含一个非血红素铁(或锰)活性中心,由一组保守残基配位(通常为三个组氨酸和一个异亮氨酸/天冬酰胺,形成HxxxxH-HxxxN(H)-I(V)基序),并位于两个长α-螺旋之间。催化过程遵循经典的“自由基”机制:Fe³⁺-OH⁻从底物脂肪酸的顺,顺-1,4-戊二烯单元的中心碳上夺取一个氢原子,形成碳中心自由基和Fe²⁺-H₂O;随后分子氧插入,最终从Fe²⁺获得一个质子,生成具有共轭二烯结构的氢过氧化物产物,同时恢复Fe³⁺状态。N端结构域具有多样性,在植物和动物的LOX中通常呈现为C2样β折叠的“PLAT”结构域,而在微生物LOX中则可能由α-螺旋组成“盖”状结构,或完全缺失。研究表明,尽管N端结构域对催化并非必需,但它通过影响酶的整体构象、稳定性和膜结合能力,在催化活性调控中发挥着重要作用。此外,底物结合口袋的形状(如“靴形”、“U形”或“T形”)及其关键残基(如底部的带电荷His或中性Leu/Gln)决定了反应的区域选择性和立体选择性,这些主要位于C端结构域内。因此,C端催化结构域的精确几何构型、底物通道的特性以及N端结构域对整体构象稳定性的贡献,共同决定了LOX的催化效率和热稳定性。这为通过蛋白质工程理性改造这些性质提供了明确的分子靶点。
目前,获取高耐热性酶主要采用两种策略。第一种是从极端微生物中筛选酶;然而,这类酶在中低温下往往表现出很低甚至可忽略的催化活性,且筛选过程通常成本高昂且耗时。第二种策略是通过蛋白质工程技术对现有脂氧合酶进行改造,以提升其催化性能。B因子(也称温度因子)可用于量化蛋白质结构中原子运动及局部柔性,从而为靶向稳定性工程提供理论依据。然而,其准确性可能受结构分辨率和精修误差的影响。因此,在比较不同结构的B值时通常需要进行归一化处理。为解决这些问题并促进稳定性工程,已开发了多种计算工具,如RONN、Maranas、SCHEMA、FireProt、CAN、Rosetta Design、FoldX、FRESCO和B-FITTER,用于识别柔性区域并建议稳定化突变。其中,B-FITTER被认为是一种普遍适用的方法,能够平衡结构柔性与刚性,为蛋白质的理性稳定化提供有价值的指导。迄今为止,基于B因子的重新设计已成功应用于提高多种酶的热稳定性和催化活性。然而,其在脂氧合酶中的应用仍相对较少,这既是一个研究空白,也是一个机遇。值得注意的是,蛋白质的热稳定性和活性之间往往存在平衡。通过增加蛋白质刚性来提高热稳定性有时会导致活性下降。因此,在保持催化活性的同时实现蛋白质稳定的方法仍是一个挑战。目前,针对脂氧合酶稳定性-活性权衡问题的研究进展有限。此外,阐明此类权衡分子机制的机理研究也仍然匮乏。
在前期工作中,一种来自球状念珠藻(N. sphaeroides)的新型脂氧合酶(NsLOX)被鉴定和表征,显示出高催化活性和宽pH耐受性;但其有限的热稳定性制约了其在食品、化工和制药行业的潜在应用。在本研究中,利用NsLOX的晶体结构,通过基于B因子的策略指导突变酶的设计,旨在大幅提高其热稳定性和活性。此外,通过三维结构模拟和分子动力学分析,探究了突变体热稳定性和活性耦合提升的分子机制。本工作也为其他酶的热稳定性提升策略提供了基础。
1. NsLOX热稳定性关键残基的筛选
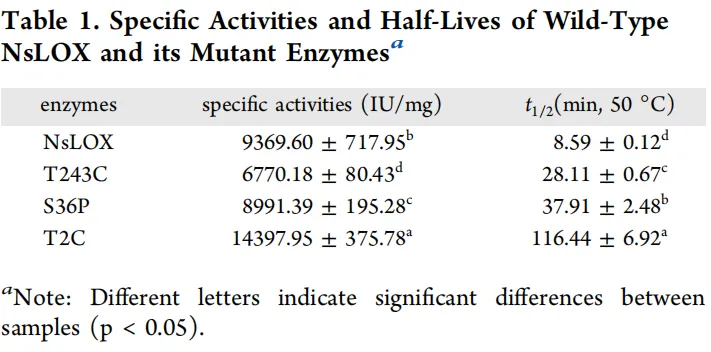
利用B-FITTER对NsLOX的晶体结构进行分析,计算每个氨基酸残基的B值。分析结果显示,B值较高(表明柔性较大)的氨基酸残基对酶的热稳定性影响更为显著。具体而言,结果表明有20个氨基酸残基的平均B值超过120。为提高研究效率,筛选出B值最高的前10个氨基酸残基,这些残基均位于柔性环区结构上(图1)。因此,Thr243、Glu242、Ser241、Lys244、Ser36、Thr2、Lys38、Ile37、Pro35和Val3很可能是影响NsLOX热稳定性的关键氨基酸残基。
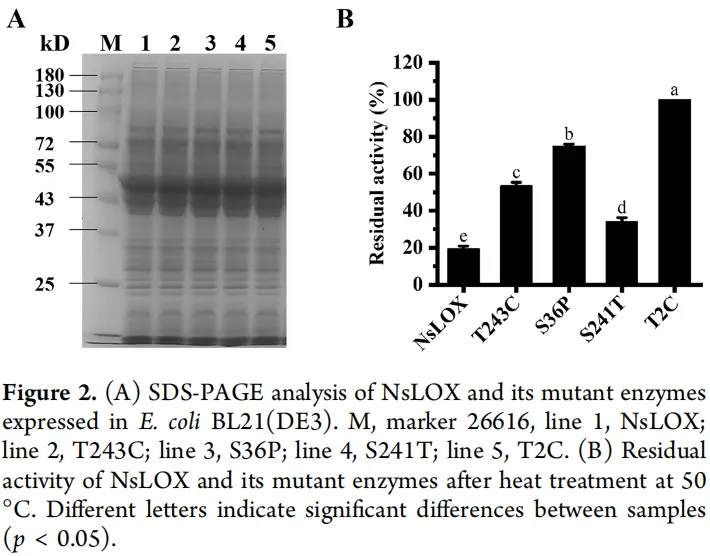
为增强NsLOX的热稳定性,对筛选出的十个氨基酸残基进行了定点饱和突变。从所得突变体中,初步鉴定出四个热稳定性显著提高的突变体,它们与野生型酶具有相似的分子量和表达水平(图2A)。在50 °C热处理后,突变体T243C、S36P和S241T的残余活性分别为53.54 ± 1.75%、74.83 ± 1.18%和33.97 ± 2.22%,均显著高于野生型。其中,T2C突变体表现尤为突出,在50 °C热处理后仍保留100%的初始活性,显著高于野生型及其他单突变体,表明其热稳定性大幅提高(图2B)。
为尽量减少粗酶液中杂蛋白对热稳定性测定的干扰,采用Ni²⁺亲和层析对野生型NsLOX及其突变酶T2C、S36P和T243C分别进行分离纯化。纯化后的突变酶与野生型具有相同的纯度和分子量。随后对野生型NsLOX及其突变酶的比活性和50 °C半衰期进行了测定,结果显示显著差异,详见表1。值得注意的是,所有突变酶的热稳定性均优于野生型。尤其引人关注的是突变酶T2C热稳定性的显著提升,其50 °C半衰期从野生型的8.59 ± 0.12 min延长至116.44 ± 6.92 min,提高了13.56倍。此外,该位点的比活性测定为14397.95 ± 375.78 IU/mg,是野生型NsLOX(9369.60 ± 717.95 IU/mg)的1.54倍。突变酶S36P和T243C在50 °C的半衰期分别为37.91 ± 2.48 min和28.11 ± 0.67 min,分别是野生型的4.41倍和3.27倍。尽管这些突变酶的比活性较野生型略有降低(分别为8991.39 ± 195.28 IU/mg和6770.18 ± 80.43 IU/mg),但仍保留了96%和72%的残余活性。
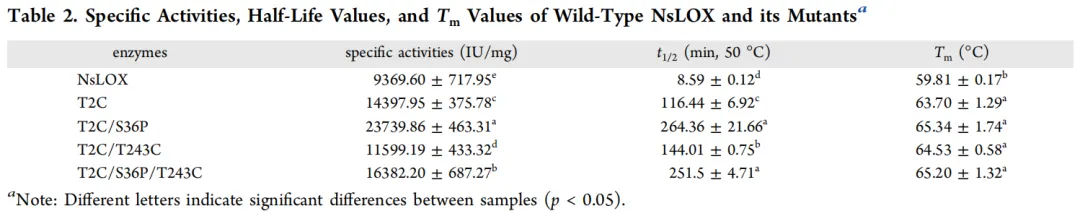
为进一步提高NsLOX的热稳定性,进行了双位点和三位点的组合突变。以突变体T2C为模板,分别与突变体S36P和T243C进行双点和三点组合突变。随后采用Ni²⁺亲和层析对组合突变体T2C/S36P、T2C/T243C和T2C/S36P/T243C进行分离纯化。纯化后的突变体与野生型NsLOX具有相同的纯度和分子量。测定了NsLOX及其突变酶T2C、T2C/S36P、T2C/T243C和T2C/S36P/T243C的比活性、50 °C半衰期和熔解温度(Tm),结果见表2。突变酶T2C/S36P和T2C/T243C的半衰期分别为264.36 ± 21.66 min和144.01 ± 0.75 min,显著高于突变酶T2C(116.44 ± 6.92 min)(*p* < 0.05),分别是野生型(8.59 ± 0.12 min)的30.78倍和16.76倍。此外,它们的比活性分别达到23739.86 ± 463.31 IU/mg和11599.19 ± 433.32 IU/mg,较野生型(9369.60 ± 717.95 IU/mg)分别提高了1.53倍和0.24倍。值得注意的是,突变酶T2C/S36P/T243C的半衰期和比活性分别为251.5 ± 4.71 min和16382.20 ± 687.27 IU/mg,分别是野生型的29.28倍和1.75倍。与突变酶T2C/S36P相比,其比活性略有下降,半衰期无显著变化(*p* > 0.05)。
酶的熱力学稳定性也可通过其熔解温度(Tm)来评估。为进一步探究其热稳定性,测定了NsLOX及其突变酶的Tm值。如表2所示,所有突变酶的Tm值均较野生型有所提高,表明突变酶的热稳定性得到改善。T2C的Tm值为63.70 ± 1.29 °C,显著高于野生型(59.81 ± 0.17 °C)。T2C/S36P的Tm值更高,达65.34 ± 1.74 °C,与野生型相比展现出优异的热稳定性。尽管T2C/S36P/T243C的Tm值(65.20 ± 1.32 °C)高于T2C/T243C(64.53 ± 0.58 °C),但与T2C/S36P相比无显著变化(*p* > 0.05),这与半衰期实验结果的变化趋势一致。综上所述,T2C在影响酶的热稳定性和催化活性方面起着关键作用,且与S36P组合时表现出协同效应。
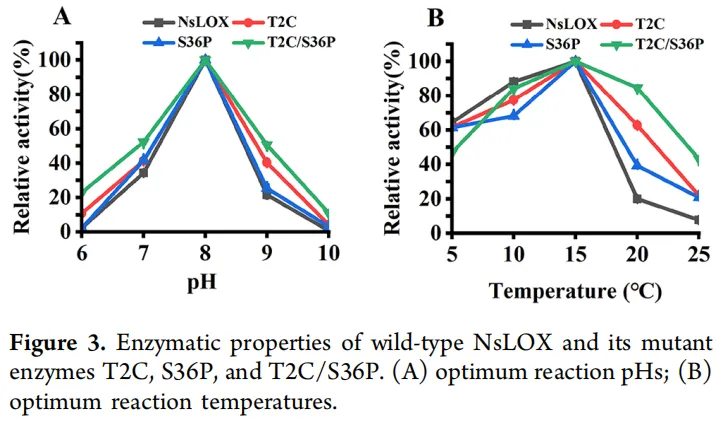
为进一步探究氨基酸残基对NsLOX酶学特性的影响,考察了NsLOX及其突变酶T2C、S36P和T2C/S36P的最适反应pH和温度。突变酶与野生型NsLOX的最适反应pH均为8.0。突变酶T2C/S36P的pH耐受范围较野生型略有拓宽,在pH 7−9范围内维持50%以上的相对活性,表明其结构稳定性增强(图3A)。NsLOX及其突变酶的最适反应温度均为15 °C。值得注意的是,在15 °C以上温度时,突变酶表现出高于野生型NsLOX的相对活性。具体而言,突变酶T2C在20 °C时保留60%以上的相对活性,而突变酶T2C/S36P在20 °C时保留80%以上,在25 °C时保留40%以上(图3B)。这些结果表明突变酶的热稳定性有所提高。
动力学参数在评估酶结合底物和加速反应速率的能力方面起着至关重要的作用。测定了野生型NsLOX及其突变酶T2C、S36P和T2C/S36P的动力学参数,结果见表3。突变酶T2C、S36P和T2C/S36P的Km值分别为7.31 ± 0.39 μM、7.37 ± 0.63 μM和17.21 ± 0.62 μM,均显著低于野生型NsLOX(19.46 ± 1.24 μM)。此外,突变酶T2C、S36P和T2C/S36P的kcat/Km值范围在472.85 μM⁻¹·s⁻¹至648.96 μM⁻¹·s⁻¹、739.88 μM⁻¹·s⁻¹、651.78 μM⁻¹·s⁻¹之间,催化效率较野生型有所提高。这些结果表明,所有突变酶对底物亚油酸的亲和力和催化效率均显著增强。
值得注意的是,不同突变体在比活性和催化效率(kcat/Km)方面表现出不同的优化侧重点。T2C和S36P主要通过大幅增强底物亲和力(Km降低约60%)实现最高的催化效率,而双突变体T2C/S36P则在显著提高催化转化速率(kcat提高约22%)的同时协同优化了亲和力,从而实现了比活性最大程度的提升。这表明本研究采用的理性设计能够差异化调控酶催化循环中的不同步骤。
4. 突变体热稳定性与催化活性协同增强的分子机制
4.1. NsLOX及其突变酶的二级结构与序列分析
为探究突变如何影响NsLOX的二级结构特征,测定了野生型NsLOX及其突变酶的远紫外圆二色(CD)光谱,并基于野生型和突变酶的CD图谱计算了二级结构含量。与野生型NsLOX相比,所有突变酶的二级结构均未发生显著变化。研究表明,氨基酸的疏水性反映了蛋白质的折叠情况,并在维持蛋白质三级结构方面发挥重要作用。疏水性指数较大的氨基酸更疏水,而疏水性指数较小的氨基酸更亲水。此外,柔性通常与蛋白质构象的柔韧性和适应性相关。当蛋白质柔性降低时,其构象刚性相对增加,从而可提高蛋白质的稳定性。因此,利用在线软件ProtScale对野生型NsLOX及其突变酶的疏水性和柔性进行分析。将氨基酸残基Thr2突变为Cys2后,与野生型相比,环区Cys2-Val10的疏水性增强、柔性降低。因此,突变酶T2C热稳定性的提高可归因于结构疏水性的增加和柔性的降低。
4.2. 野生型NsLOX及其突变酶的分子动力学分析
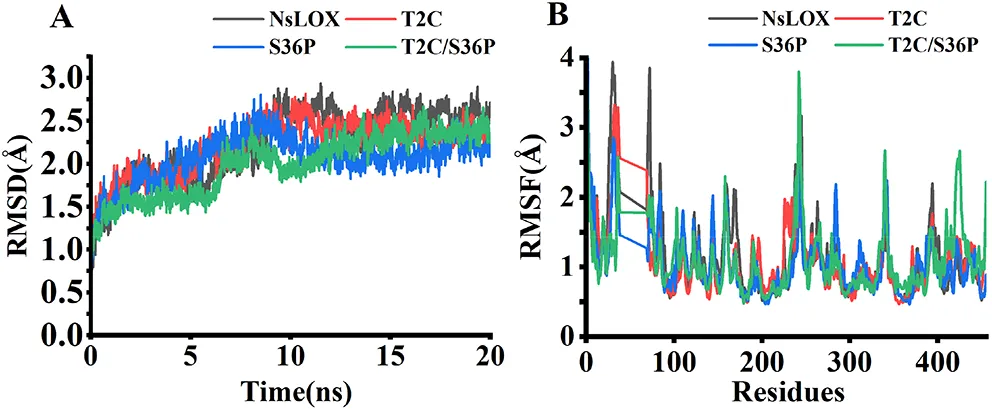
为从动力学角度分析突变酶热稳定性增强的分子机制,以野生型NsLOX的晶体结构为模板,构建了突变酶的三维结构模型,并利用AMBER 18软件对野生型及突变酶T2C、S36P和T2C/S36P的结构模型进行了分子动力学模拟。考虑到野生型的熔解温度为59.81 ± 0.17 °C,模拟温度选择为330 K(56.85 °C)。均方根偏差(RMSD)是评估蛋白质构象热波动的重要参数,与蛋白质刚性和热稳定性呈负相关。经分子动力学模拟分析,野生型NsLOX及其突变酶的RMSD结果如图4A所示,反映了模拟过程中蛋白质主链Cα的位置偏移。野生型的RMSD值上升更为显著,在9 ns时从0.2 nm跃升至约0.25 nm,表明野生型NsLOX在该模拟温度下极不稳定。突变酶T2C、S36P和T2C/S36P的平均RMSD值分别为2.067 Å、2.207 Å和1.997 Å,均低于野生型(2.239 Å),表明突变酶均比野生型更刚性,整体结构更稳定。
均方根波动(RMSF)值反映了MD模拟中蛋白质主碳链Cα的运动情况,RMSF值较大的残基具有更多的自由度,而RMSF值较小的残基自由度较低。野生型NsLOX及其突变酶在20 ns模拟期间的RMSF值如图4B所示。与野生型相比,突变酶T2C、S36P和T2C/S36P的某些区域(包括残基Asn10-Ile15、Asp24-Leu75、Pro84-Glu90、Gly123-Phe135、Gln146-Val185、His223-Leu228、Leu270-Asn274、Glu325-Asp330和Met388-Gln403)均具有较低的RMSF值。突变酶热波动区域的分布大幅减少,表明突变酶柔性降低,从而显著增强了酶的热稳定性。综上所述,结构刚性增加和柔性降低是突变酶T2C、S36P和T2C/S36P热稳定性提高的分子机制。
4.3 野生型NsLOX及其突变酶的三维结构分析
4.3.1. 突变酶热稳定性增强的机制解析
为进一步阐明突变影响酶热稳定性的机制,分析了关键位点(Thr2/Cys2、Ser36/Pro36、Thr243/Cys243)周围残基相互作用网络的变化及各突变体的表面静电势。结合50 °C半衰期测定结果(表1、2),发现局部相互作用的重塑与整体热稳定性之间存在明确的构效关系。
(1)突变酶相互作用网络分析
在T2C突变体中,野生型中Cys2与Ser4之间的疏水相互作用转变为氢键(3.09 Å),同时与Glu5和Gln6的氢键网络得以维持和优化。这种“氢键替代疏水”策略显著增强了2号位点区域的局部刚性和极性相互作用,被认为是其热稳定性大幅提高的主要原因。S36P突变遵循了不同的路径:在保留与Lys38原有疏水相互作用的基础上,Pro36新形成了与Glu225的强疏水相互作用,同时与Trp39和Gln40的氢键距离进行了适应性调整。这种“疏水增强与氢键重塑”策略在酶分子远端构建了更稳定的疏水核心,从而适度提高了稳定性。在T243C突变体中,Thr243被Cys取代后与Ser241形成了新的氢键(3.31 Å),取代了原有的疏水相互作用。这种“局部氢键增强”策略同样带来了稳定性的提高。
值得注意的是,T2C/S36P双突变体的相互作用网络并非两个单点突变的简单叠加,而是表现出显著的协同和调整效应。首先,在2号位点,T2C单点突变中形成的与Ser4的氢键被Cys2放弃,恢复了疏水相互作用,而与Glu5和Gln6的优化氢键得以保留。这表明在S36P创造的构象背景下,“疏水接触+精简氢键”的组合比全氢键网络更为有利。其次,在36号位点,S36P建立的疏水网络(与Lys38和Gln225)及重塑的氢键被完全继承。最关键的是,在243号位点,Thr243明确地维持了与Ser241的疏水相互作用,完全避免了T243C的氢键模式(图S4C、F、I)。这三个位点的变化共同定义了一个新颖、高度协同且无冲突的稳定网络,使其热稳定性达到所有突变体中的峰值(约为野生型的30.8倍),远超单点突变体之和,表现出强烈的正协同效应。
然而,在T2C/S36P/T243C三突变体中,T243C的引入触发了优化网络的系统扰动,导致两个关键变化:首先,36号位点的疏水网络发生重构。Pro36的疏水相互作用伙伴从Lys38/Glu225变为Leu34/Lys38,导致与Glu225的重要相互作用丢失。其次,2号位点的最优模式发生退化。Cys2被迫与Ser4重新形成氢键(3.13 Å),从双突变体的最优“疏水+氢键”模式退化为类似于T2C单点突变的次优“氢键替代”模式。这一系列为迁就T243C而做出的能量不利的链式调整,最终抵消了T243C可能带来的有限增益。因此,三突变体的热稳定性(251.5 min)略低于双突变体(264.36 min),催化活性顺序为:T2C/S36P > T2C/S36P/T243C > T2C。这清楚地表明,T243C的稳定化机制与T2C/S36P的协同网络不相容,其引入特异地扰乱了Pro36的疏水网络,并迫使Cys2氢键模式发生退化,最终引发了性能的“天花板效应”。
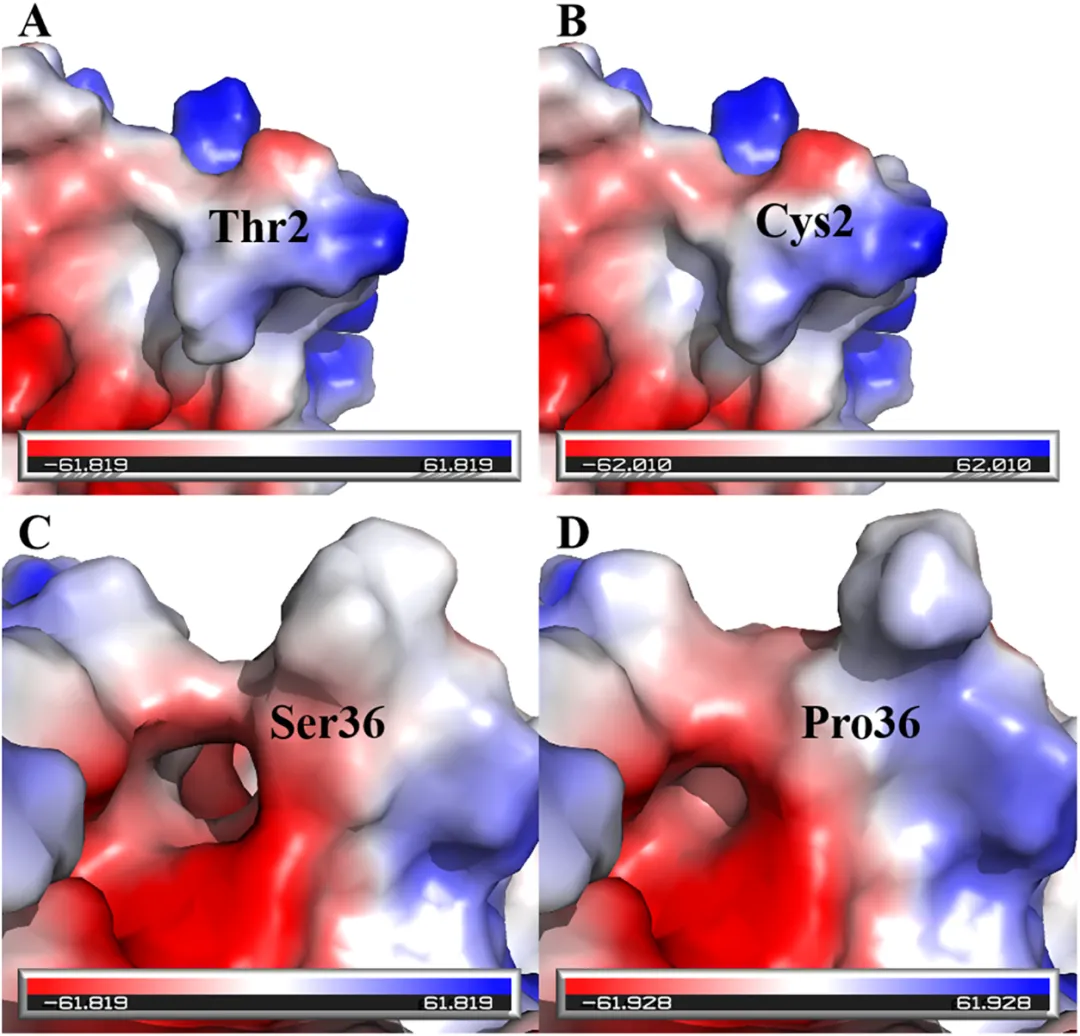
(2)突变酶表面静电势分析
表面静电势的分布被认为是影响蛋白质折叠稳定性和功能的关键理化性质之一。已有研究表明,表面静电势的优化,尤其是通过增强其对称性和正电性,可通过改善分子内相互作用以及蛋白质与溶剂之间的相互作用显著提高酶的热稳定性。为探究T2C和S36P突变体热稳定性增强的结构基础,计算并可视化了它们的表面静电势。如图5所示,与野生型(图5A)相比,T2C(图5B)和S36P(图5D)突变体整体表面上观察到更广泛且连续的正电势区域分布。定量分析表明,两种突变体的“对称正电势”值(T2C:62.010;S36P:61.928)均高于野生型(61.819)。特别值得注意的是,在野生型结构中,位于活性中心附近的Ser36残基处于局部负电势“口袋”中(图5C),这种与周围整体正电势环境的不匹配被认为是潜在的局部结构不稳定因素。相比之下,S36P突变成功将该区域转变为与周围环境一致的正电势分布(图5D)。因此,本研究提出表面静电势的优化可能也是T2C和S36P突变体实现显著热稳定性的重要结构贡献因素之一。
4.3.2. 突变酶比活性提高的机制解析
为进一步分析突变如何影响酶的催化活性,利用Discovery Studio Client对野生型及其突变体复合物结构进行了酶-底物相互作用网络分析。在野生型中,底物LA与His192、His197、Leu193、Ile230、Leu238、Leu386和Leu410之间形成烷基相互作用。LA羧基的O⁻与Thr406形成氢键,与Arg54和Arg402形成π-阴离子相互作用(图6A)。在T2C突变体中,底物LA与Phe43、His192、His197、Leu193、Ile230、Leu238、Val248和Leu410之间形成烷基相互作用。LA羧基的O⁻与Thr406和Asn409形成氢键,与Arg54和Arg402形成π-阴离子相互作用。此外,LA羧基的O与Arg402形成氢键(图6B)。在S36P突变体中,底物LA与Phe43、His192、His197、Leu193、Ile230、Leu238、Leu386和Leu410之间形成烷基相互作用。LA羧基的O⁻与Thr406形成氢键,与Arg402形成π-阴离子相互作用。此外,LA羧基的O与Arg402之间也形成氢键(图6C)。在双突变体T2C/S36P中,底物LA与Leu47、Val247、His192、His197、Leu193、Ile230、Leu238、Leu386和Leu410之间形成烷基相互作用。LA羧基的O⁻与Arg402形成π-阴离子相互作用,而LA羧基的O与Thr406形成氢键(图6D)。进一步定量分析发现,野生型和S36P中的烷基相互作用数量均为14个,T2C中为15个,T2C/S36P中为16个(图6)。
综上所述,基于生化数据分析(表2),催化活性顺序为:T2C/S36P > T2C > 野生型,而S36P的活性与野生型相近。Arg402被确定为关键的“活性开关”,其羧基与底物之间静电/氢键相互作用(π-阴离子或氢键)的增强被认为是正向影响活性的关键共同因素(涉及T2C、S36P和T2C/S36P)。相比之下,Arg54被视为重要的“稳定基础”,其π-阴离子相互作用的缺失会导致活性显著降低(如S36P所示),表明其在维持基础活性网络中的重要作用。值得注意的是,T2C/S36P并非沿T2C中“持续增加氢键”的路径发展,而是采用了类似S36P的“精简氢键”策略。其成功归因于两个关键决策:保留了与Arg402的π-阴离子相互作用,从而维持了静电稳定性的基线;同时大幅重塑了疏水口袋,引入了与Leu47和Val247的两个新颖且强烈的疏水相互作用。这被认为显著优化了底物结合构象,使得仅存的Thr406氢键能够以最高效率发挥作用。
催化活性的优化被视为一项系统工程任务。T2C/S36P突变组合被认为巧妙地平衡了“静电稳定性”和“疏水匹配”:通过保留与Arg402的关键静电相互作用维持了稳定性,同时通过创造新型疏水空间实现了底物结合构象的精确优化,从而达到了活性峰值。这为理性设计提供了比单纯“增加氢键”更深入的依据。研究表明,重塑底物结合口袋的疏水拓扑结构可能比简单地增加极性相互作用更为有效。
基于B因子的分子改造是半理性设计策略中提高酶热稳定性的有效方法,是一种较为通用的分子进化策略。该策略通过分子设计在结构柔性与刚性之间寻求平衡,为突破蛋白质结构多样性和分子进化路径复杂性的限制、重塑和增强蛋白质分子稳定性提供了指导和见解,并已广泛应用于蛋白质工程领域。B因子通过对蛋白质晶体结构的X射线衍射数据进行计算分析获得,反映原子电子层的弥散程度,且受蛋白质晶体质量和结构分辨率的显著影响。B因子的概念源于晶体学研究,在缺乏蛋白质晶体结构的情况下,有研究借助同源建模、分子动力学模拟等计算模拟技术,依赖蛋白质序列和同源相似性信息来预测蛋白质分子的结构。因此,每个氨基酸残基的B因子也可通过这些计算模拟技术进行预测。然而,与使用源自X射线数据的B因子相比,所有计算模拟技术均存在显著的不确定性,其精度不超过10−15%。
在早期阶段,利用不同在线平台(Swiss-Model、i-TASSER和Phyre 2)对NsLOX进行的同源建模产生了显著差异的模拟结构和B值,这初步揭示了依赖计算预测所固有的不确定性。基于这些模型筛选出的突变体I302V,其热稳定性仅获得有限的提高(半衰期仅增加0.38倍),且伴随活性的轻微降低(比活性降至野生型水平的96.23%)。其他LOX也有类似现象的报道;例如,基于同源模型衍生B值对鱼腥藻(Anabaena sp.)脂氧合酶(AnaLOX)进行改造,半衰期仅提高1.18倍。这些结果共同表明,在LOX工程中,依赖预测的B值往往导致热稳定性改进不理想,且常伴随活性损失。其共同的深层原因是模型本身的不准确性可能误导设计方向。因此,本研究转而使用从NsLOX高分辨率晶体结构中提取的精确B值。通过B-FITTER计算每个残基的B值,并采用定点饱和突变结合组合突变技术,成功获得了热稳定性显著增强的突变体——T2C、S36P、T243C和T2C/S36P/T243C。其中,双突变体T2C/S36P在50 °C下的半衰期为264.36 ± 21.66 min,是野生型的30.78倍,ΔTm为+5.53 °C,同时其活性得到良好维持。相比之下,通过定向进化开发的AnaLOX热稳定性突变体在50 °C下的半衰期相对有限,仅约30 min,而共有序列设计的突变体N130D/G260Q的半衰期仅提高5.6倍,ΔTm为+2.45 °C。此外,通过自组装肽融合实现的铜绿假单胞菌(Pseudomonas aeruginosa)LOX热稳定性提高仅限于2.3−4.5倍。本研究结果清楚地表明,基于晶体结构计算精确B值的设计不仅大幅提高了实验可靠性,而且有效克服了LOX热稳定性工程中常见的“精度依赖性”和“功能-稳定性权衡”瓶颈。该方法为同类酶的理性设计提供了一种高度可转移的策略。
值得注意的是,尽管稳定性显著提高,所有突变体的最适反应温度仍保持在15 °C不变,这一现象与某些定向进化研究和N端截短修饰中LOX的报道一致,但不同于其他策略(如共有序列设计和融合技术)中最适温度提高5−20 °C的情况。这一对比清晰地说明了热稳定性与催化温度适应性之间的解耦,源于它们不同的分子决定因素。本研究所采用的B因子策略增强了蛋白质主链局部柔性区域,导致热稳定性显著提高,同时维持了最适反应温度不变。然而,催化的最适温度主要由活性位点中限速步骤的温度敏感性决定,该步骤通常涉及一系列协调的构象动力学。该结果表明,仅依赖于增强结构刚性的稳定化策略未必能重新校准催化循环中精细调控的动力学网络。为同时提高酶的最适温度,未来的蛋白质工程工作应进一步整合面向功能动力学的策略,如调控底物通道的柔性或优化催化中间体的能量景观,这可通过分子动力学模拟和定向进化等组合方法实现。
许多结构因素影响蛋白质的热稳定性,如盐桥、二硫键、氢键、二级结构、疏水性、柔性、蛋白质表面静电势和氨基酸组成。本研究利用在线服务器ProtScale对野生型及其突变酶进行了疏水性和柔性分析,结果表明将Thr2突变为Cys2增加了残基Thr2-Val10的疏水性并降低了其柔性,从而增强了结构的紧密性。对于S36P,其热稳定性的提高可归因于脯氨酸侧链的吡咯环赋予其特殊的构象刚性,从而使其酶分子结构更加稳定。Li等人通过将位于EcAII β-转角处的Asp178替换为脯氨酸,提高了酶分子的热稳定性。对南极纤毛虫来源的冷适应α-淀粉酶进行定点诱变,将关键氨基酸残基E166、S185和T350突变为脯氨酸,导致其在高温(40和50 °C)下的热稳定性显著提高,而不影响其在5−35 °C下的催化活性。此外,对野生型及其突变酶进行三维(3D)结构模拟分析和分子动力学模拟分析表明,整体结构刚性增强、部分环区柔性降低以及蛋白质表面有利正电势的改善是突变酶热稳定性增强的分子机制。
值得注意的是,尽管T2C突变在序列水平上增加了Cys2-Val10环区的疏水性并降低了理论柔性,但在分子动力学模拟中该区域的原子波动(RMSF)并未显著降低。这一现象可能解释为:虽然T2C突变部分“刚性化”了环区的起始端,但其主要的稳定化效应并非直接限制环区自身的运动,而是像“杠杆效应”一样将稳定能量传递到蛋白质中其他更易受热扰动的关键区域(如图4B中显示RMSF显著降低的几个区域)。换言之,该突变可能优化了结构的整体刚性分布,优先加固了整体的“薄弱环节”,从而在不显著改变局部环区动力学的前提下实现了整体热稳定性的提高。这表明一个有效的稳定化突变可以通过长程变构效应影响蛋白质的整体能量景观,其作用机制比简单地增加局部刚性更为复杂。
基于静电势分析结果,并结合本研究的结构模型及类似研究中观察到的一般规律,提出了关于稳定性增强的合理机制推论:T2C和S36P突变诱导的局部正电势增强主要通过以下两条途径稳定蛋白质构象。首先,在蛋白质内部,增强的正电势可能促进或加强与邻近带负电残基之间的有利静电相互作用。这种分子内静电网络的巩固被认为有助于“锁定”突变区域的构象,从而降低其热振动柔性。其次,在蛋白质-溶剂界面,更均匀的带正电表面被认为有利于与溶液中带负电的离子或极性水分子形成更有序、更稳定的溶剂化层。这种优化的外部相互作用不仅被认为能减少疏水表面的暴露,从而抑制高温下因疏水聚集引起的失活,还能通过提供额外的溶剂化能来稳定蛋白质的天然折叠状态。这一发现与近期若干工业酶热稳定性工程研究的结论一致,表明表面静电相互作用的理性调控是打破酶活性与稳定性之间权衡关系的有效策略之一。当然,也认识到蛋白质稳定性的增强通常是多因素协同作用的结果。静电势的改善与本研究观察到的其他局部相互作用优化(如特定氢键或疏水相互作用的形成)共同构成了一个整合的稳定化网络。未来可通过分子动力学模拟等方法对上述静电相互作用的具体贡献进行进一步的动态验证。
文献报道,多个有益氨基酸残基的组合突变通常对蛋白质的热稳定性具有累加或协同效应,少量文献表明突变之间也可能存在拮抗效应。本研究中,突变酶T2C/S36P和T2C/T243C的半衰期分别为264.36 ± 21.66 min和144.01 ± 0.75 min,显著高于突变酶T2C(116.44 ± 6.92 min)(p < 0.05),表现出协同效应。相比之下,突变酶T2C/S36P/T243C的半衰期为251.5 ± 4.71 min,与突变酶T2C/S36P相比无显著变化(p > 0.05),未表现出累加或协同效应。如第3.4.3.1节所述,T2C/S36P通过精细的“氢键-疏水相互作用模式重构”——具体而言,Cys2恢复了疏水接触,Pro36构建了远端疏水核心,Thr243维持了疏水沉默——形成了一个内部协同性高、冲突最小化的稳定网络。这种网络层面的优化,而非孤立位点效应的简单叠加,构成了稳定性和活性同时突破的结构基础。在三突变体T2C/S36P/T243C中未获得进一步增益,并观察到了“潜在负面影响”。结构分析表明,尽管T243C本身的引入能够增强稳定性,但在T2C/S36P已经高度优化的网络背景下,其微妙的局部改变触发了远端位点的非适应性重构(Pro36的疏水网络从Lys38/Glu225变为Leu34/Lys38),并迫使关键位点发生退化(Cys2重新形成氢键)。因此,协同增益被部分抵消。这种“上位效应”深刻表明,在多点工程中,任何新添加的突变都必须评估其与现有优化网络的整体兼容性;否则可能诱发性能倒退。双突变酶T2C/S36P的比活性是野生型的1.53倍。众所周知,酶的高催化活性与高热稳定性之间存在长期存在的权衡问题,本研究基于结构解析计算的B值策略权衡了酶的催化活性与热稳定性之间的关系,为其他酶的分子改造提供了依据。
酶的催化活性和热稳定性是评估其工业适用性的两个关键性能指标。具有高催化活性和良好热稳定性的酶在工业环境中更具竞争力和应用潜力。研究表明,酶的催化效率主要取决于其活性位点的结构柔性,而酶分子的整体稳定性与其三维结构的刚性密切相关。这种对结构和功能的双重需求导致了酶工程中普遍存在的“稳定性-活性”权衡效应。这一科学挑战已成为限制工业酶性能提升的关键瓶颈。本研究基于结构指导的B因子策略,对NsLOX进行了分子设计,获得了催化活性和热稳定性协同增强的双突变体。T2C突变主要增强了N端区域的局部稳定性和刚性(对热稳定性有贡献),并可能通过引入额外的氢键(例如与Asn409)略微优化了活性。S36P突变通过优化表面静电势提高了整体稳定性(热稳定性),其“精简氢键”模式为双突变体中的活性优化提供了概念性方法。两者结合后,在获得累加的热稳定性增益的同时,引入的局部变化(如T2C/S36P联合作用可能诱导的构象调整)意外而有效地重塑了远端底物结合疏水口袋。热稳定性通过增强整体结构刚性和优化表面性质实现。
稳定的蛋白质骨架为高效催化提供了必要的基础。增强的整体刚性(作为热稳定化机制)导致结构更加刚性和波动更小。这一稳定化的平台进而确保了底物结合口袋精确而稳定的三维几何结构。在此基础上,进一步实施了口袋的精确工程,以实现最佳的“静电-疏水”平衡。这涉及重塑疏水拓扑结构以优化底物结合构象,同时保留关键的静电相互作用。整体稳定化与局部精细调控的协同整合,最终实现了热稳定性和催化活性的双重提升。这些发现表明,在酶工程中,将整体折叠的稳定化与活性位点微环境的精确调控相结合,是开发高性能突变体的有效策略。
尽管本研究通过体外酶动力学分析充分验证了突变体的催化性能,但需要认识到,全细胞生物转化和长期酶反应实验对于全面评估酶的工业应用潜力具有不可替代的价值。这些实验能够提供酶在实际应用条件下的催化效率、操作稳定性和产物积累能力等关键信息。因此,在未来的研究中将进行全细胞生物转化实验,以评估NsLOX及其突变体在复杂反应体系中的催化性能;同时,通过长期酶反应和连续操作实验研究酶的操作稳定性和产物抑制效应。这些研究将进一步揭示NsLOX在工业生物催化中的应用前景,并为其实际应用开发奠定基础。
综上所述,基于X射线晶体学的B因子分析,成功工程化了一种热稳定性和催化活性均增强的NsLOX变体。这是首批利用B值理性设计球状念珠藻脂氧合酶以同时提高热稳定性和活性的报道之一。通过二级结构、序列比对、分子动力学模拟和三维结构分析的进一步研究,揭示了双突变体T2C/S36P协同改进的潜在机制:增强的整体结构刚性和优化的表面性质有助于热稳定性的提高。在此稳定化框架的基础上,底物结合口袋内实现了重新平衡的“静电-疏水”环境——具体而言,通过重塑疏水拓扑结构优化底物结合构象,同时保留关键静电相互作用——从而协同提升了热稳定性和催化活性。本研究为提高NsLOX的热稳定性提供了一种有效的工程策略和机制见解,这在临床和食品工业应用中具有广阔的前景。




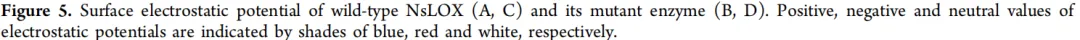
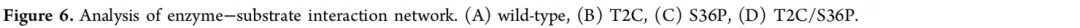
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
