优化肌纤维类型分布是提升牛肉品质的重要策略,为探讨白藜芦醇对牛肌管细胞肌纤维类型转化的作用及其与Akt/FoxO信号通路的介导机制,山东农业大学的逯莹,孙静,朱立贤,杨啸吟,刘昀阁,梁荣蓉,毛衍伟,张一敏,左惠心等学者采用MTT比色法检测不同浓度白藜芦醇、渥曼青霉素(PI3K抑制剂)、SC79(Akt激活剂)对牛肌管细胞活力的影响及其作用浓度。以400 μmol/L H2O2处理牛肌管细胞24 h建立氧化应激模型。采用实时荧光定量PCR(qPCR)技术检测肌纤维类型的标志基因(MyHC Ⅰ、MyHC Ⅱa、MyHC Ⅱx、MyHC Ⅱb)以及Akt/FoxO信号通路关键基因(PI3K、Akt、FoxO1、FoxO3)的mRNA表达水平。浓度梯度实验表明,10 μmol/L白藜芦醇对氧化应激下肌纤维类型的调控效果较优,可显著逆转这些基因表达趋势(P<0.05)。qPCR检测显示,白藜芦醇处理可显著上调Akt/FoxO通路正调控因子(PI3Kα、PI3Kβ、Akt1、Akt2)及氧化代谢相关基因(MEF2C、PGC-1α)的mRNA表达,抑制负调控因子(FoxO1、FoxO3)转录(P<0.05)。Western blot实验进一步证实,白藜芦醇能提高PI3K蛋白表达及Akt、FoxO1、FoxO3的磷酸化水平,降低FoxO1、FoxO3总蛋白含量(P<0.05);而渥曼青霉素可显著阻断该调控效应,同时削弱白藜芦醇对慢肌纤维蛋白的促进作用和对快肌纤维蛋白的抑制作用(P<0.05)。此外,H2O2处理可抑制PI3K/Akt通路活性,白藜芦醇与SC79协同作用可恢复通路正常功能。研究表明,白藜芦醇通过激活Akt/FoxO信号通路,有效逆转氧化应激诱导的牛肌管细胞快肌化趋势,促进慢肌纤维基因表达,抑制快肌纤维基因表达。研究旨在探讨基于Akt/FoxO信号通路的白藜芦醇对氧化应激诱导的肌纤维类型转化的影响机制,阐明其改善牛肉食用品质的作用路径,为天然活性物质在畜禽肉品质调控中的应用提供理论支撑。
肌纤维是肌肉组织的基本构成单位,其类型与组成在很大程度上影响着肉品的品质[1],根据肌球蛋白重链及代谢特征分为慢速氧化型(Ⅰ型)、快速氧化-酵解型(Ⅱa型)、中间型(Ⅱx型)和快速酵解型(Ⅱb型)4种类型[2]。Ⅰ型与Ⅱa型纤维线粒体丰富,以有氧代谢为主[3],其占比高,通常对应更好的肉品质[4];Ⅱx型和Ⅱb型依靠糖酵解,其比例升高往往伴随嫩度下降、风味变差及滴水损失增加等肉品质降低的表现[5]。肌纤维类型具有可塑性[6],受遗传、环境、营养等因素影响[7]。氧化应激作为动物机体中最普遍的应激类型,会对肉质产生一系列负面影响,包括引起脂质与蛋白质的氧化、诱发细胞凋亡、提高肌肉剪切力以及增加滴水损失等。肌肉中的纤维类型在氧化代谢能力上存在差异,通常慢肌纤维富含更多线粒体,其氧化酶活性也较高;相比之下,快肌纤维中线粒体数量较少,氧化酶活性偏低。氧化应激导致活性氧异常增加,损害线粒体功能[8],是导致肌肉萎缩[9]、代谢紊乱的重要因素[10]。因此,氧化应激与肌纤维类型之间存在重要的内在联系,尚需进一步研究。白藜芦醇(resveratrol,RES)是一种天然多酚物质,具备抗氧化、抗衰老和代谢调节等功能[11]。有研究显示,RES可改变肌纤维类型[12],进而改善肌肉功能和品质[13]。本团队前期研究发现,RES可通过AMPK/PGC-1α通路促进牛肌管细胞向慢肌纤维转化[14]。然而,RES是否能通过其他信号通路协同调控肌纤维类型转化,尤其是在氧化应激环境下的作用及其机制,仍有待深入阐明。在调控肌纤维转化的过程中,Akt/FoxO信号通路发挥了重要的作用。Akt激活可抑制FoxO转录活性[15],阻断其核定位,从而促进慢肌纤维特征维持[16];相反,FoxO激活则可能诱导快肌纤维相关基因表达[17]。但目前尚不清楚H2O2诱导的氧化应激是否影响肌纤维快肌化过程,以及RES能否影响牛肌管细胞Akt/FoxO通路。
本研究拟建立牛肌管细胞模型,探究H2O2诱导的氧化应激和RES对肌纤维类型及Akt/FoxO通路关键基因表达的影响,建立一种稳定可靠的氧化应激诱导肌纤维快肌化转化的细胞模型,为后续深入研究RES通过Akt/FoxO信号通路拮抗氧化应激的作用机制提供依据。
1.1 材料与试剂
牛成肌细胞,上海一研生物科技有限公司;RES(纯度>99%,细胞用),美国MCE公司;渥曼青霉素、Akt激活剂(SC79)、过氧化氢(H2O2),美国Selleck生物科技有限公司;马血清、胎牛血清、最低必需培养基(MEM)、MTT噻唑蓝,北京索莱宝科技有限公司;Evo M-MLV反转录预混型试剂盒、SYBR Green Pro Taq HS 预混qPCR 试剂盒,湖南艾科瑞生物工程有限公司;BCA蛋白定量试剂盒,北京康为世纪生物科技有限公司;PMSF溶液、磷酸酶抑制剂混合物,上海碧云天生物技术股份有限公司;PVDF膜,Millipore公司。
1.2 仪器与设备
AB104-S型分析天平,瑞士Mettler Toledo公司;BD-145HDE型冷藏冷冻转换柜,青岛海尔特种电冰柜有限公司;Nikon Eclipse E100型光学显微镜,日本Nikon公司;3111型二氧化碳培养箱,美国赛默飞世尔科技公司;JXFSTPRP-2型冷冻研磨机,上海净信实业发展有限公司;P330型核酸蛋白测定仪,德国慕尼黑Implen公司;5840R型高速冷冻离心机,德国Eppendorf生物科学公司;BioTekEpoch2型酶标仪,美国伯腾仪器有限公司;T100型PCR仪、1658029型电泳仪、CFX96型实时荧光定量PCR仪、ChemiDoc MP型凝胶成像仪,美国Bio-Rad有限公司。
1.3 实验方法
1.3.1 培养基及试剂制备
取89 mL MEM培养基、10 mL胎牛血清、1 mL双抗(青霉素-链霉素)制备100 mL细胞增殖培养基;取97 mL MEM培养基、2 mL马血清、1 mL双抗,制备100 mL细胞分化培养基,配制完成后于4 ℃保存。取34 μL H2O2溶液、2 966 mL细胞分化培养基制备10 mmol/L过氧化氢母液。取11.412 mg RES溶于1 mL DMSO中配制50 mmol/L RES母液,于-80 ℃下可保存6个月,在-20 ℃下可保存1个月。取5 mg渥曼青霉素粉末溶于1.167 1 mL DMSO中配制10 mmol/L渥曼青霉素母液,于-80 ℃下保存。取10 mg SC79粉末溶于1.370 7 mL DMSO中配制20 mmol/L SC79母液,于-80 ℃下保存。
1.3.2 牛肌管细胞培养
参考Zhang等[18]的方法并稍加修改。将牛成肌细胞在含有体积分数10%胎牛血清和体积分数1%双抗的最低必需培养基中,于37 ℃、体积分数5% CO2条件下培养到融合度70%左右,更换为含体积分数2%马血清的分化培养基诱导分化,待牛肌管形成后用于实验。
1.3.3 牛肌管细胞的处理
为探索氧化应激建模条件、氧化应激对牛肌管细胞肌纤维类型转化的影响,及探究渥曼青霉素与RES的安全浓度,当细胞融合度达到70%左右时,更换成细胞分化培养基,诱导分化3 d。使用含有不同浓度H2O2溶液(0、100、200、300、400、500 μmol/L)的分化培养基,分别处理不同时间(0、6、12、24 h),收集细胞用于相关指标测定,每批重复处理3次。
为了探究各试剂的安全浓度范围,诱导分化3 d后,换用含有不同浓度渥曼青霉素(0、1、2、4、8、16 μmol/L)和不同浓度RES(0、10、20、30、40、50 μmol/L)的分化培养基,分别处理细胞24 h,收集细胞用于相关指标测定,每批重复处理3次。
1.3.4 牛肌管细胞活力检测
调节细胞浓度至1×105 cells/mL,以每孔200 μL接种于96孔板,并置于37 ℃、体积分数5%的CO2培养箱中进行培养。细胞贴壁后,更换分化培养基继续诱导分化3 d,之后更换为含有不同浓度RES、渥曼青霉素、SC79的培养基继续培养。待处理结束,首先移除各孔内的培养基,随后立即向每孔加入100 μL MTT溶液(0.5 mg/mL),并将细胞板放回培养箱中持续孵育4 h[19]。随后吸弃上清液,每孔加入100 μL DMSO,避光条件下振荡10 min,最后在570 nm测定吸光度,并计算细胞相对活力。
1.3.5 牛肌管细胞mRNA表达水平测定
参考Zhang等[18]的方法并稍加修改。采用RNAiso Plus试剂提取牛肌管细胞中的总RNA,再利用琼脂糖凝胶电泳检测RNA的完整性。以OD260与OD280的比值(OD260/OD280)判断RNA的纯度,OD260/OD280值为1.8~2.0,说明RNA未受到污染,可用于后续实验。根据Evo M-MLV反转录预混型试剂盒合成cDNA。扩增反应在实时荧火定量PCR检测系统上进行,使用2×SYBR®预混试剂。引物采用Beacon Designer软件设计,以甘油醛-3-磷酸脱氢酶(GAPDH)为内参基因,采用2-ΔΔCT分析mRNA相对表达量的变化。所用基因引物序列见表1。
表1 实时荧光定量PCR引物序列
Tab.1 Primer sequences used for real-time quantitative fluorescent PCR
F为上游引物(forward primer),R为下游引物(reverse primer)。
1.3.6 牛肌管细胞蛋白测定
1.3.6.1 蛋白质的提取
研磨管中加入0.5 g肉样、5 mL冰冷提取液、PMSF和磷酸酶抑制剂,加入一大一小钢珠,研磨仪45 s研磨3次。4 ℃、2 789 r/min离心15 min,得上清液。取100 μL上清液稀释50倍,使用BCA蛋白定量试剂盒测定蛋白质含量。取800 μL蛋白质定量后的样品于离心管中,加入等体积的蛋白上样缓冲液(2×loading buffer,质量分数1%溴酚蓝和β-巯基乙醇),金属浴95 ℃变性5 min,制得5 mg/mL的蛋白液。
1.3.6.2 蛋白质表达水平的测定
配制质量分数8%分离胶,试剂混匀后打入玻璃板间隙,用纯水将胶压平,30 min后倒掉纯水并用滤纸吸干。加入配制好的5%浓缩胶,插好梳子待胶凝固。浓缩胶凝固后,将其放入电泳槽并加入电泳液,拔梳子上样,依次将marker和蛋白质样品打入浓缩胶梳子孔中进行电泳,上样量为30 μg。电泳条件90 V、30 min,120 V、55 min。电泳结束后,采用湿转法将胶上的蛋白质转印到PVDF膜。裁下所需条带,用甲醇激活PVDF膜,转印夹板依次放入海绵、3张滤纸、凝胶、PVDF膜、3张滤纸、海绵中,然后将其放入转印槽中,倒入快速转膜液,转印条件为恒流400 mA转膜30 min。转印结束后,用质量分数5%脱脂奶粉或者质量分数5% BSA将PVDF膜在摇床上室温封闭90 min。封闭结束后TBST洗膜,5 min洗3次。将对应分子质量的PVDF膜置于对应稀释过的一抗中,摇床4 ℃孵育12 h左右。TBST洗膜3次,每次洗5 min。将PVDF膜置于稀释好的二抗中,在摇床上室温孵育90 min。TBST洗膜3次,每次洗5 min。每张膜滴加700 μL显影液反应2 min,利用凝胶成像仪进行拍照成像。
1.4 数据处理
采用SPSS 26.0软件进行统计分析。利用独立样本t检验和单因素方差分析(one-way ANOVA)进行比较分析。当分析结果显示存在显著性差异时,进一步利用最小显著差异法(least significant difference,LSD)进行事后两两对比检验,P<0.05表示差异具有统计学意义。所有实验均独立重复3次,结果以平均值±标准偏差(mean±SD)表示。相关图表的绘制使用Origin 2022软件。
2.1 牛肌管细胞氧化应激模型的建立
2.1.1 H2O2处理对牛肌管细胞活力的影响
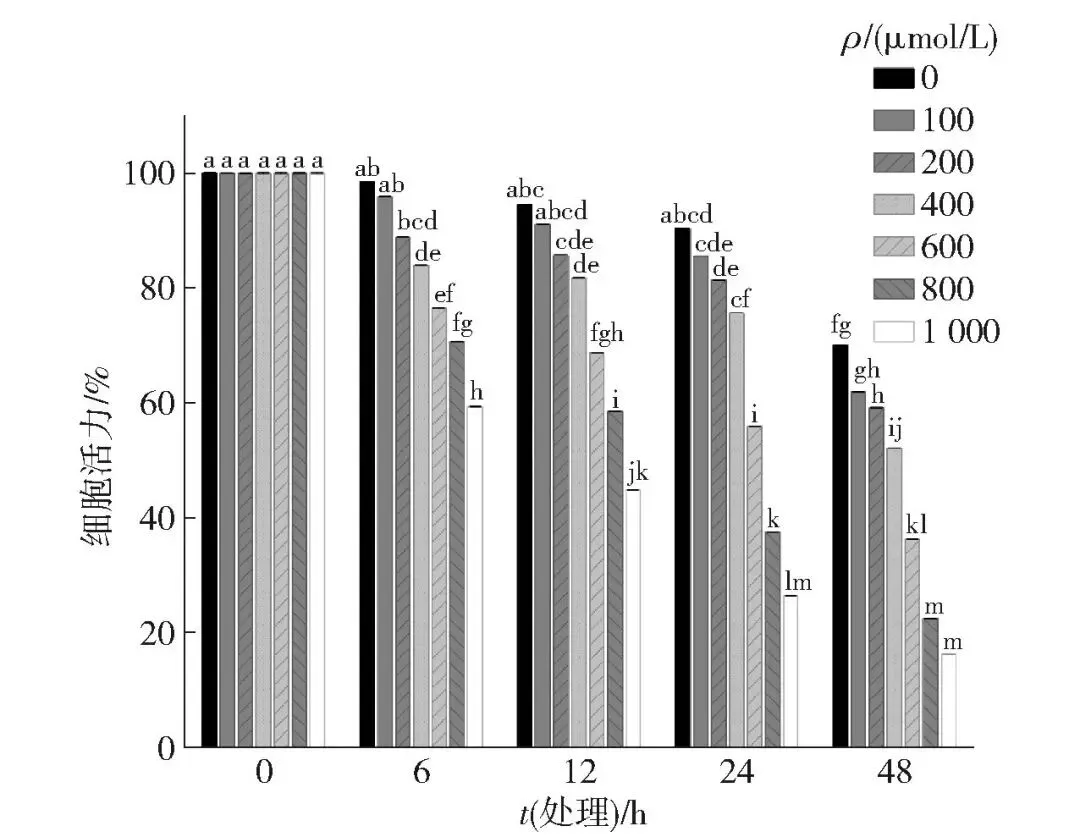
为模拟体内氧化应激环境,本研究采用H2O2处理牛肌管细胞以建立氧化应激模型。为评估氧化应激对牛肌管细胞活力的影响,采用不同浓度H2O2(0、100、200、400、600、800、1 000 μmol/L)分别处理牛肌管细胞0、6、12、24、48 h,采用MTT法检测细胞活力变化,结果如图1。随着H2O2浓度的升高和处理时间的延长,牛肌管细胞活力呈逐渐下降趋势。400 μmol/L H2O2诱导处理在表征牛肌管细胞代谢抗氧化反应中具有显著作用。有研究表明,特定浓度的H2O2可触发谷胱甘肽调控途径,这表明所选H2O2浓度能在可控范围内诱导氧化应激,避免过度损伤,保证了模型的稳定性和可重复性[20]。400 μmol/L H2O2处理6~24 h后,细胞活力维持在80%左右,符合适度损伤的建模原则。后续实验选择400 μmol/L H2O2作为氧化应激模型的诱导条件。
不同字母代表细胞活力水平差异显著(P<0.05)。
图1 不同H2O2浓度和处理时间对牛肌管细胞活力的影响Fig.1 Effects of different H2O2 concentrations and treatment time on cell viability of bovine myotubues
2.1.2 H2O2处理时间对肌纤维类型相关基因表达的影响
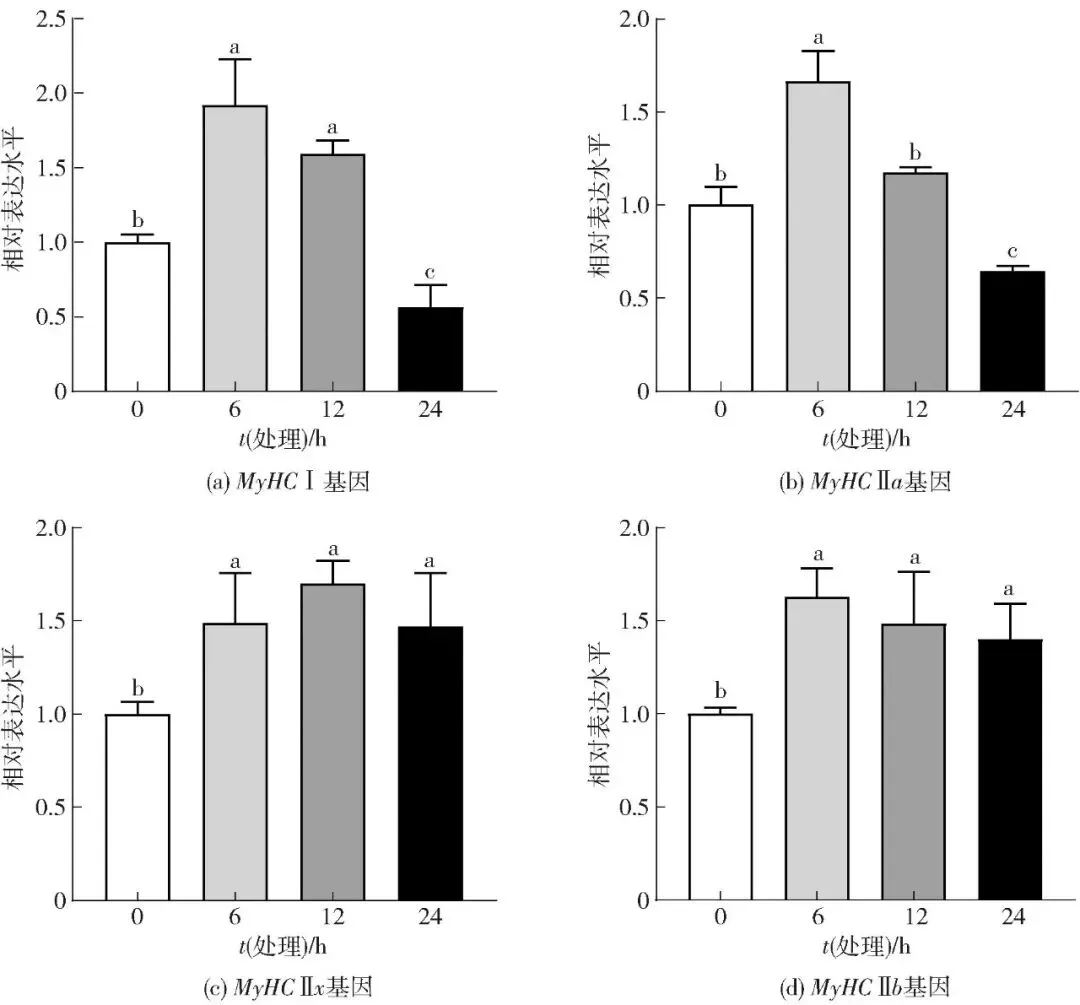
氧化应激可对骨骼肌线粒体功能造成损害,并引起能量代谢异常[21]。鉴于慢肌纤维(Ⅰ型和Ⅱa型)具有较高的线粒体含量,推测该应激状态可能降低其表达水平。为验证这一推测,本研究利用实时荧光定量PCR技术检测了400 μmol/L H2O2处理不同时间(0、6、12、24 h)对牛肌管细胞中肌纤维类型相关基因表达的影响,结果见图2。经24 h氧化应激处理后,慢肌纤维标志基因MyHC Ⅰ和MyHC Ⅱa的表达显著下调(P<0.05),而快肌纤维标志基因MyHC Ⅱx和MyHC Ⅱb的表达则显著上调(P<0.05)。氧化应激处理牛肌管细胞24 h促进了肌纤维类型由慢肌向快肌的转化。氧化应激处理6、12 h后,Ⅰ型和Ⅱa型肌纤维基因表达没有明显下降,其原因可能在于初期活性氧(reactive oxygen species,ROS)水平仍处于细胞内氧化还原体系的调控范围,尚未对线粒体结构及功能造成显著损害。因此,确定400 μmol/L H2O2处理24 h为氧化应激诱导快肌化的体外模型条件。
不同字母表示组间数据差异显著(P<0.05)。
图2 不同H2O2处理时间对牛肌管细胞中肌纤维类型相关基因表达水平的影响Fig.2 Effects of different H2O2 treatment time on related gene expression levels of myofiber type in bovine myotubes
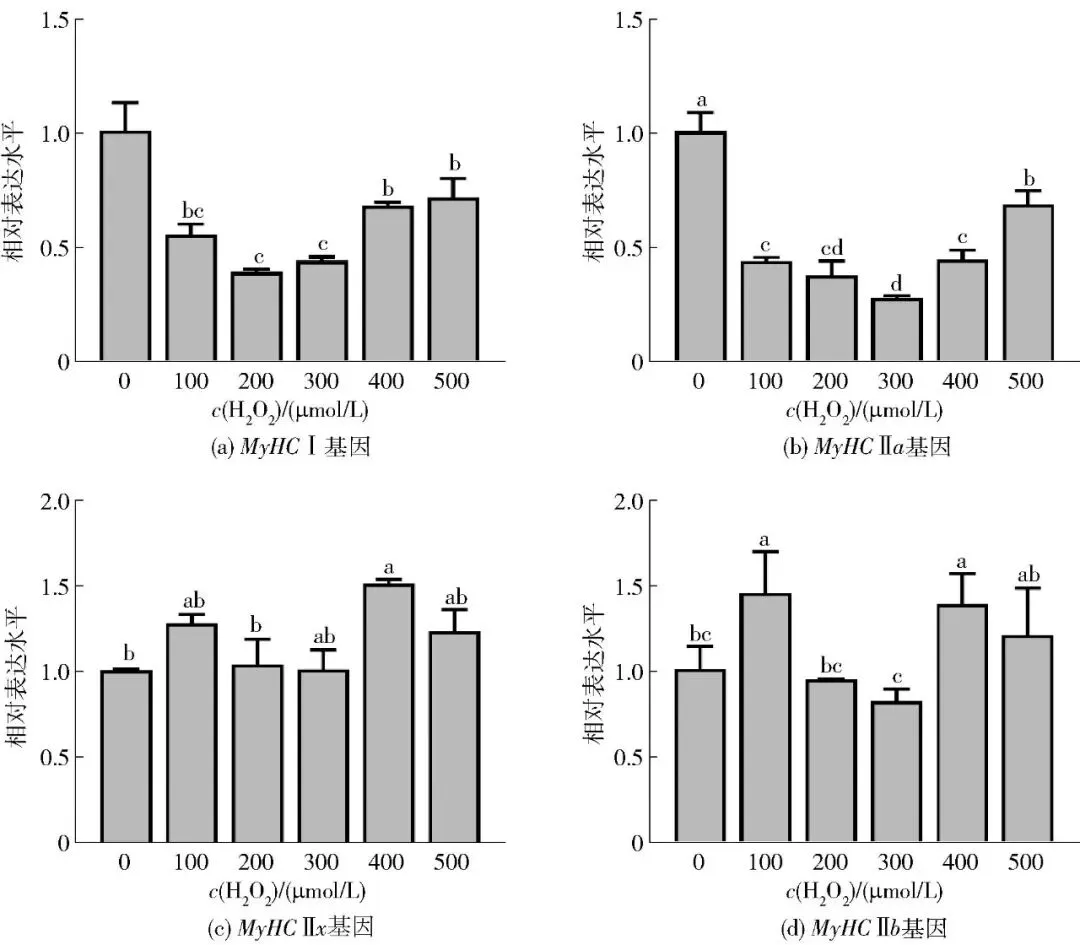
2.1.3 H2O2处理浓度对肌纤维类型的影响
H2O2处理浓度对肌纤维类型的影响见图3。在已确定的400 μmol/L H2O2处理24 h的条件下,进一步用实时吸光定量PCR法验证该浓度对肌纤维类型基因的表达效应。结果显示,与对照组相比,MyHC Ⅰ和MyHC Ⅱa mRNA表达水平显著降低,而MyHC Ⅱx和MyHC Ⅱb表达水平显著升高(P<0.05),证实该条件可稳定诱导慢肌向快肌转化,用作后续氧化应激模型。
不同字母表示组间数据差异显著(P<0.05)。
图3 不同浓度H2O2对牛肌管细胞中肌纤维类型相关基因表达水平的影响Fig.3 Effects of different concentrations of H2O2 on related gene expression levels of myofiber type in bovine myotubes
2.2 RES、渥曼青霉素、SC79对牛肌管细胞活力的影响
2.2.1 RES对牛肌管细胞活力的影响
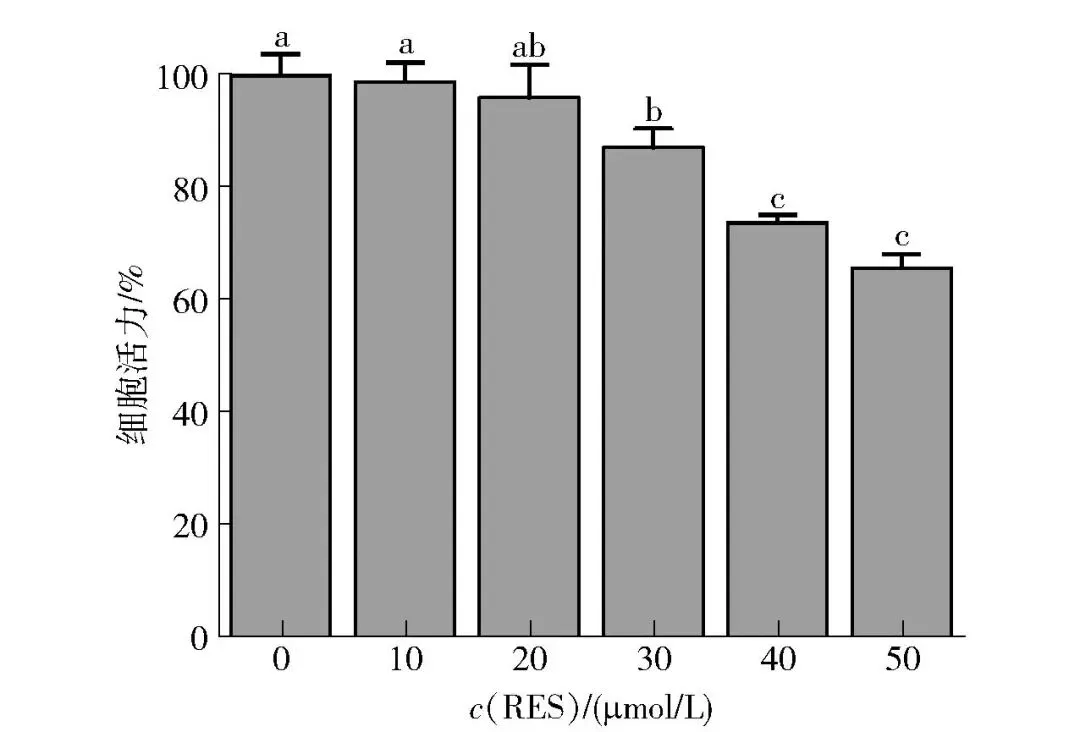
为了观察RES对牛肌管细胞活力的影响,选用不同浓度的RES(0、10、20、30、40、50 μmol/L)处理牛肌管细胞,MTT法检测细胞活力,结果如图4所示。RES浓度为0~30 μmol/L时,牛肌管细胞的活力均在87%以上,表明RES无明显细胞毒性。与对照组相比,10 μmol/L RES处理的牛肌管细胞活力变化最小。有研究表明,RES浓度小于30 μmol/L时,可显著提升慢肌纤维相关基因(MyHC Ⅰ和MyHC Ⅱa等)的表达,而不显著干扰细胞存活率,这也验证了RES低浓度下的良好生物相容性[18]。
不同字母表示组间数据差异显著(P<0.05)。
图4 白藜芦醇对牛肌管细胞活力的影响Fig.4 Effects of resveratrol on cell viability in bovine myotubes
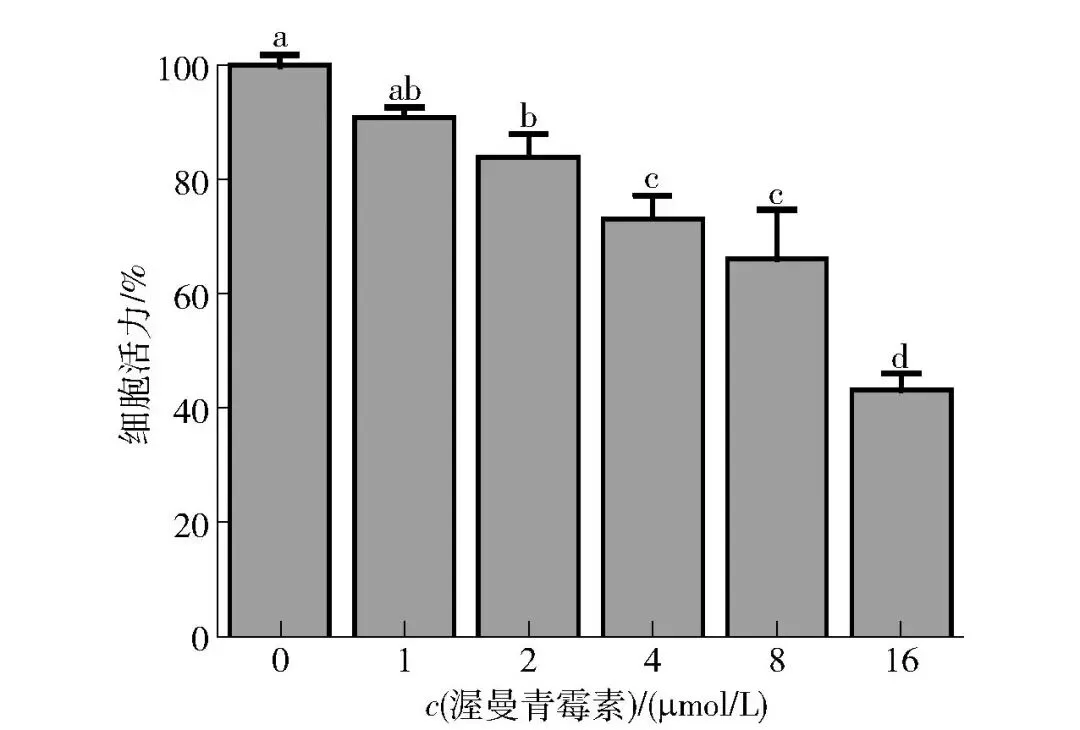
2.2.2 渥曼青霉素对牛肌管细胞活力的影响
渥曼青霉素是一种高效且高选择性的PI3K信号通路抑制剂[22],其作用机制在于能够直接抑制PI3K的催化活性,有效阻断该激酶介入下游的信号活动,进而抑制Akt/FoxO通路的信号传导。本研究用MTT法检测了不同浓度渥曼青霉素(0、1、2、4、8、16 μmol/L)对牛肌管细胞活力的影响,结果如图5所示。0~16 μmol/L时,随着渥曼青霉素浓度的增加,牛肌管细胞活力逐渐降低。与对照组相比,1 μmol/L的渥曼青霉素孵育牛肌管细胞24 h后细胞活力未显著降低(P>0.05)。因此,后续的实验可以用1 μmol/L的渥曼青霉素处理牛肌管细胞。
不同字母表示组间数据差异显著(P<0.05)。
图5 渥曼青霉素对牛肌管细胞活力的影响Fig.5 Effects of wortmannin on cell viability of bovine myotubes
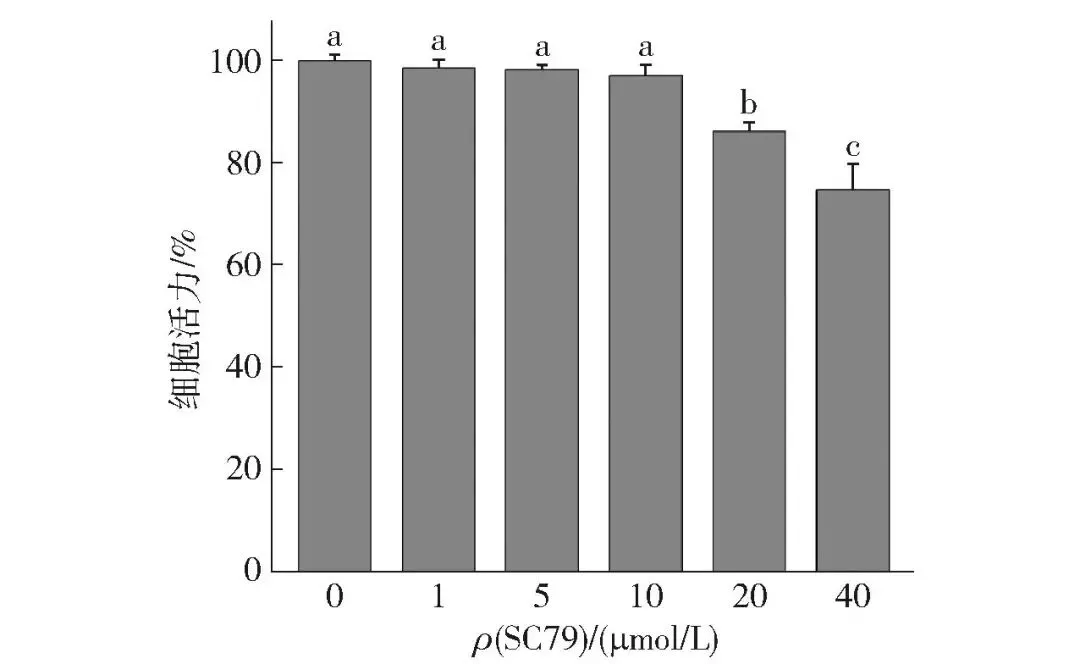
2.2.3 SC79对牛肌管细胞活力的影响
SC79是一种特异性Akt激活剂,可以与Akt的plecktrin同源性结构域结合,增强其磷酸化水平和活性[23]。Jo等[24]研究发现,SC79在Thr308和Ser473位点促进了小鼠大脑组织中Akt的磷酸化。本研究用MTT法检测了不同浓度的SC79(0、1、5、10、20、40 μmol/L)对牛肌管细胞活力的影响,结果如图6所示。SC79浓度为0~40 μmol/L时,随着SC79浓度增加,牛肌管细胞活力逐渐降低。与对照组相比,1、5、10 μmol/L SC79孵育牛肌管细胞24 h后,细胞活力未显著降低(P>0.05)。So等[25]研究发现,10.96 μmol/L的SC79促进了血清饥饿细胞和血清培养基中细胞胰岛素样生长因子诱导的Akt磷酸化。10 μmol/L为SC79作用于牛肌管细胞的适宜安全浓度,该浓度下可在保证细胞正常活力的基础上发挥Akt激活作用。
不同字母表示组间数据差异显著(P<0.05)。
图6 SC79对牛肌管细胞活力的影响Fig.6 Effects of SC79 on cell viability of bovine myotubes
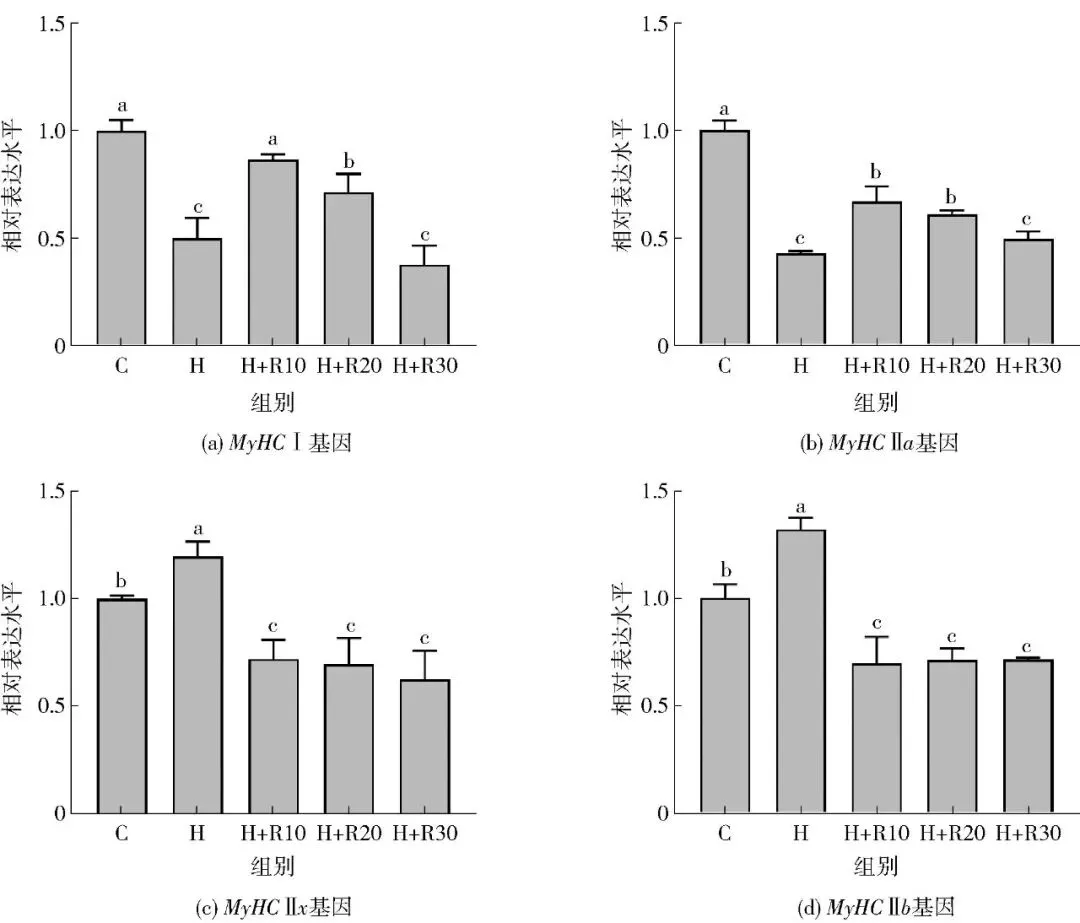
2.3 不同浓度RES对氧化应激条件下牛肌管细胞肌纤维类型的影响
选用不同浓度的RES(10、20、30 μmol/L)在氧化应激前预处理牛肌管细胞24 h,采用实时荧光定量PCR法检测牛肌管中肌纤维类型相关基因的表达,探究RES对氧化应激条件下肌纤维变化的浓度效应,结果如图7所示。10 μmol/L和20 μmol/L的RES可以显著提高氧化应激诱导的牛肌管细胞MyHC Ⅰ和MyHC Ⅱa的mRNA表达水平(P<0.05);在10~30 μmol/L时显著降低了氧化应激诱导的牛肌管细胞MyHC Ⅱx和MyHC Ⅱb的mRNA表达水平(P<0.05)。其中,10 μmol/L的RES对氧化应激诱导的牛肌管细胞肌纤维类型组成变化改善效果最好,后续实验选用RES的浓度为10 μmol/L。
C,对照组;H,400 μmol/L H2O2处理组;H+R10,400 μmol/L H2O2+10 μmol/L RES处理组;H+R20,400 μmol/L H2O2+20 μmol/L RES处理组;H+R30,400 μmol/L H2O2+30 μmol/L RES处理组。不同字母表示组间数据差异显著(P<0.05)。
图7 不同浓度RES对氧化应激诱导的牛肌管细胞中肌纤维类型相关基因表达水平的影响Fig.7 Effects of different concentrations of resveratrol on related gene expression levels of myofiber type in oxidative stress-induced bovine myotubes
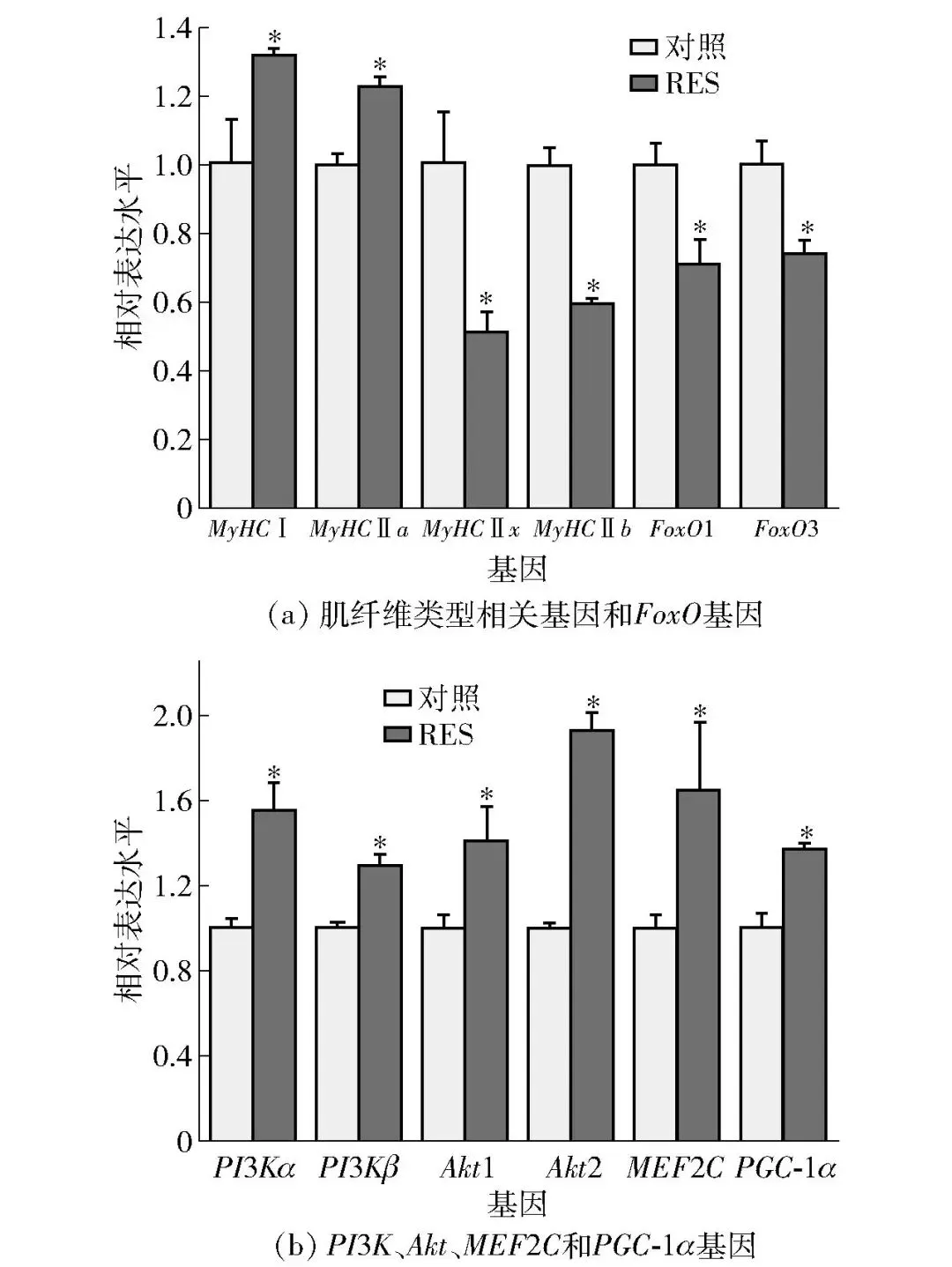
2.4 RES对牛肌管细胞肌纤维类型及Akt/FoxO信号通路调控的影响
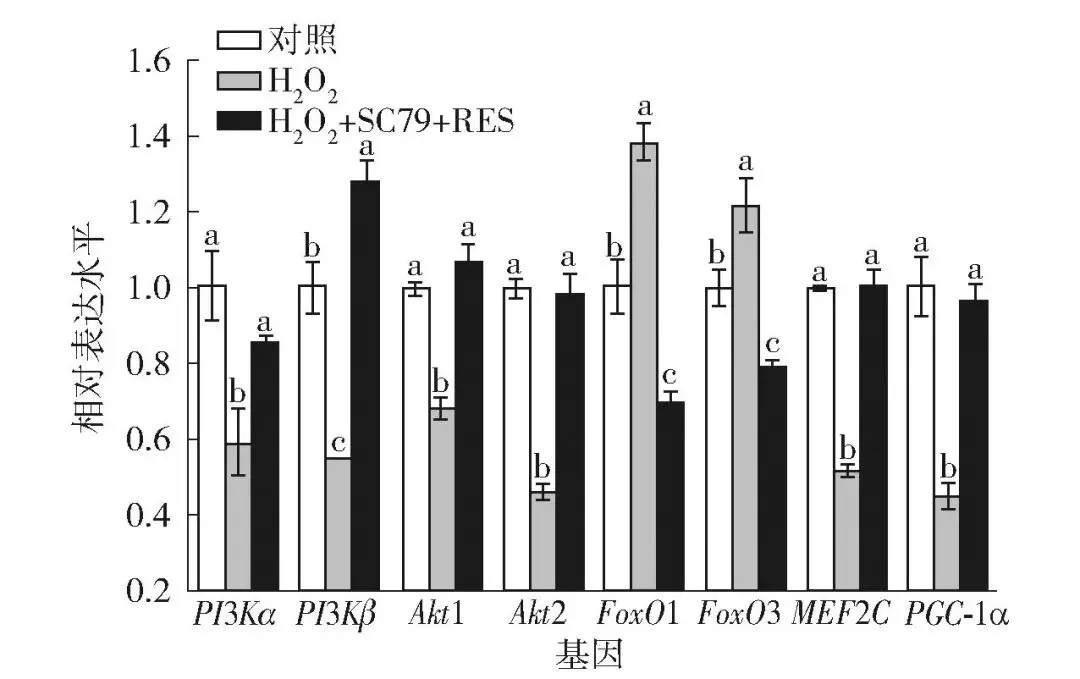
以牛肌管细胞为研究对象,以体外实验评估RES经Akt/FoxO信号通路对促进肌纤维类型转化的作用(图8)。本实验室前期研究发现,10 μmol/L RES可激活AMPK/PGC-1α信号通路,进而诱导牛肌管细胞发生由快肌向慢肌的转化[14]。因此,实验采用了10 μmol/L RES干预牛肌管细胞,并利用实时荧光定量PCR技术检测了肌纤维类型标志基因、Akt/FoxO信号通路关键基因的mRNA表达水平。FoxO1主要激活蛋白降解基因(Atrogin-1/MuRF1),FoxO3主要调控线粒体动态与PGC-1α介导的氧化代谢,10 μmol/L RES处理24 h后,慢肌纤维标志基因(MyHC Ⅰ、MyHC Ⅱa)及氧化代谢主控基因(MEF2C、PGC-1α)的表达水平均显著升高(P<0.05),而快肌纤维标志基因(MyHC Ⅱx、MyHC Ⅱb)的表达显著降低(P<0.05)。与此同时,Akt上游激活因子PI3Kα/β及其2种亚型Akt1/2的转录水平显著上调,下游负调控转录因子FoxO1/3被抑制(P<0.05)。表明RES可能通过激活Akt/FoxO信号通路,从而促进牛肌管细胞肌纤维类型转化。
*表示组间数据差异显著(P<0.05)。
图8 RES对牛肌管细胞肌纤维类型标志基因及 Akt/FoxO信号通路基因表达水平的影响Fig.8 Effects of resveratrol on expression levels of myofiber type marker genes and Akt/FoxO signaling pathway genes in bovine myotubes
2.5 SC79联合白藜芦醇对氧化应激牛肌管细胞Akt/FoxO通路的影响
为验证激活Akt/FoxO通路对白藜芦醇逆转氧化应激诱导肌纤维快肌化的调控效应,本实验采用10 μmol/L SC79与10 μmol/L白藜芦醇联合干预氧化应激诱导的牛肌管细胞,检测Akt/FoxO通路关键基因表达的变化,结果如图9所示。与对照相比,400 μmol/L H2O2处理24 h显著抑制PI3Kα、PI3Kβ、Akt1、Akt2、MEF2C及PGC-1α mRNA的表达,同时升高FoxO1与FoxO3的表达水平(P<0.05,n=3)。10 μmol/L SC79+10 μmol/L RES协同处理后,PI3Kα、Akt1、Akt2、MEF2C及PGC-1α基因表达恢复至对照水平(P>0.05),PI3Kβ基因表达显著上调(P<0.05),FoxO1与FoxO3基因表达仍显著低于对照(P<0.05)。未设置SC79或RES单独处理组,结果仅反映联合效应。
不同字母表示不同处理间差异显著(P<0.05)。
图9 RES协同SC79逆转H2O2对Akt/FoxO通路关键基因表达的抑制效应Fig.9 Reversal of H2O2-induced suppression on key gene expression in Akt/FoxO pathway by resveratrol combined with SC79
2.6 渥曼青霉素阻断Akt/FoxO通路对白藜芦醇调控肌纤维类型转化的影响
为进一步验证Akt/FoxO通路是白藜芦醇调控牛肌管细胞肌纤维类型转化的核心介导通路,本实验采用1 μmol/L渥曼青霉素预处理牛肌管细胞,阻断Akt/FoxO信号通路,分别检测通路阻断后白藜芦醇对肌纤维类型标志蛋白、通路核心蛋白表达的调控效应变化。
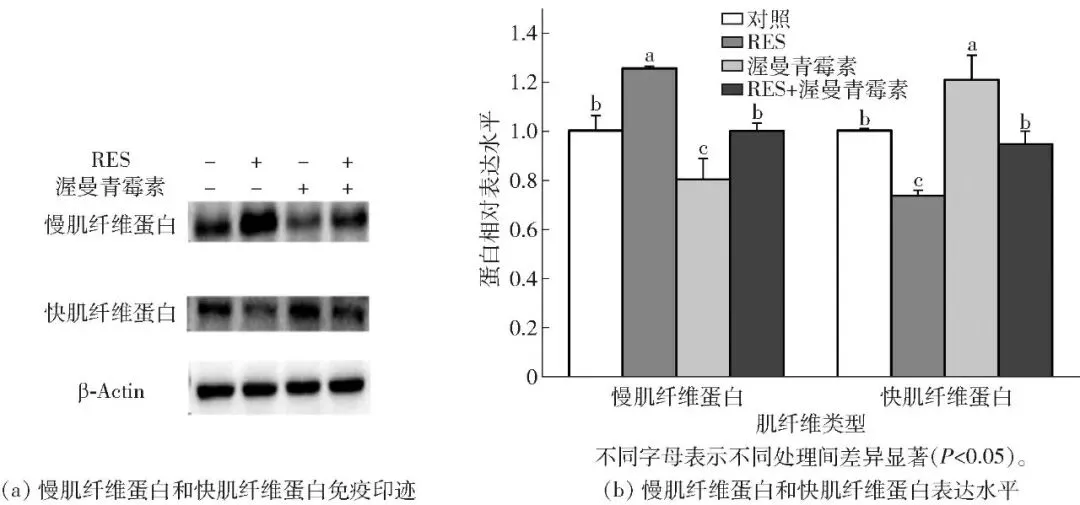
2.6.1 白藜芦醇调控肌纤维类型标志蛋白表达的影响
蛋白免疫印迹结果如图10。结果表明,RES上调慢肌纤维蛋白表达的作用被显著削弱(P<0.05),同时白藜芦醇下调快肌纤维蛋白表达的作用被显著削弱(P<0.05);而渥曼青霉素预处理阻断Akt/FoxO通路后,RES促进慢肌纤维蛋白表达、抑制快肌纤维蛋白表达的调控效应均被显著削弱(P<0.05)。
图10 渥曼青霉素阻断Akt/FoxO通路对白藜芦醇调控牛肌管细胞Akt/FoxO通路核心蛋白表达的影响Fig.10 Effects of Akt/FoxO pathway blockade by wortmannin on resveratrol regulated core protein expression of Akt/FoxO pathway in bovine myotubes
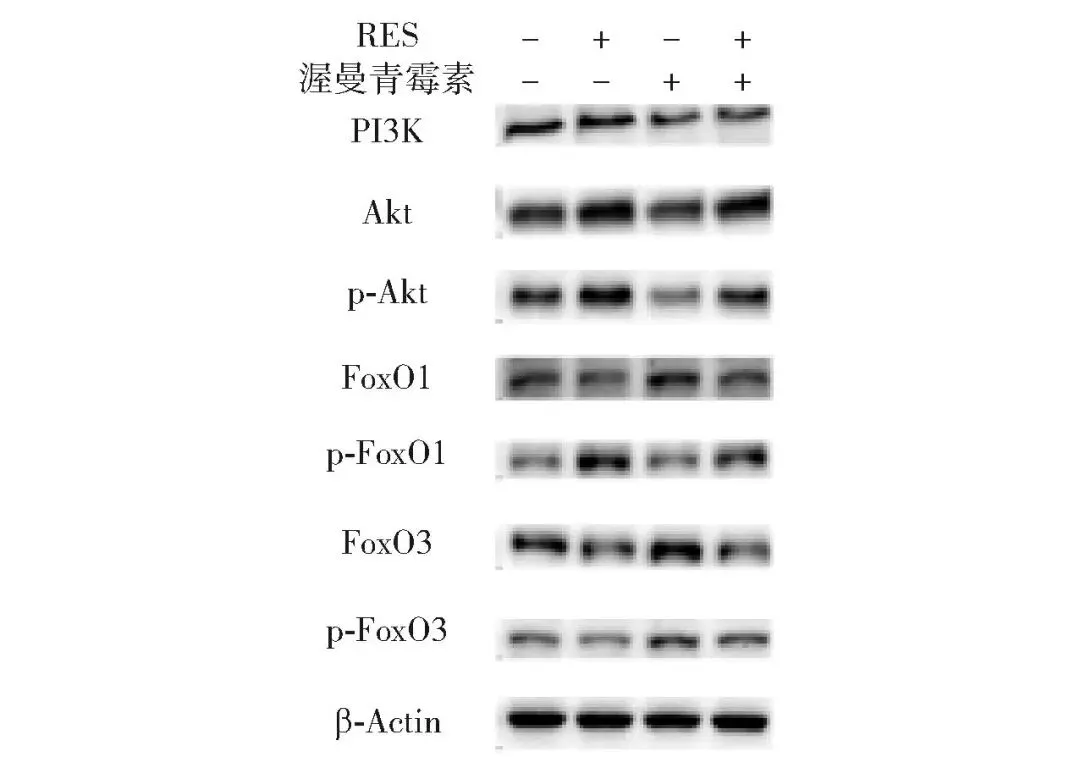
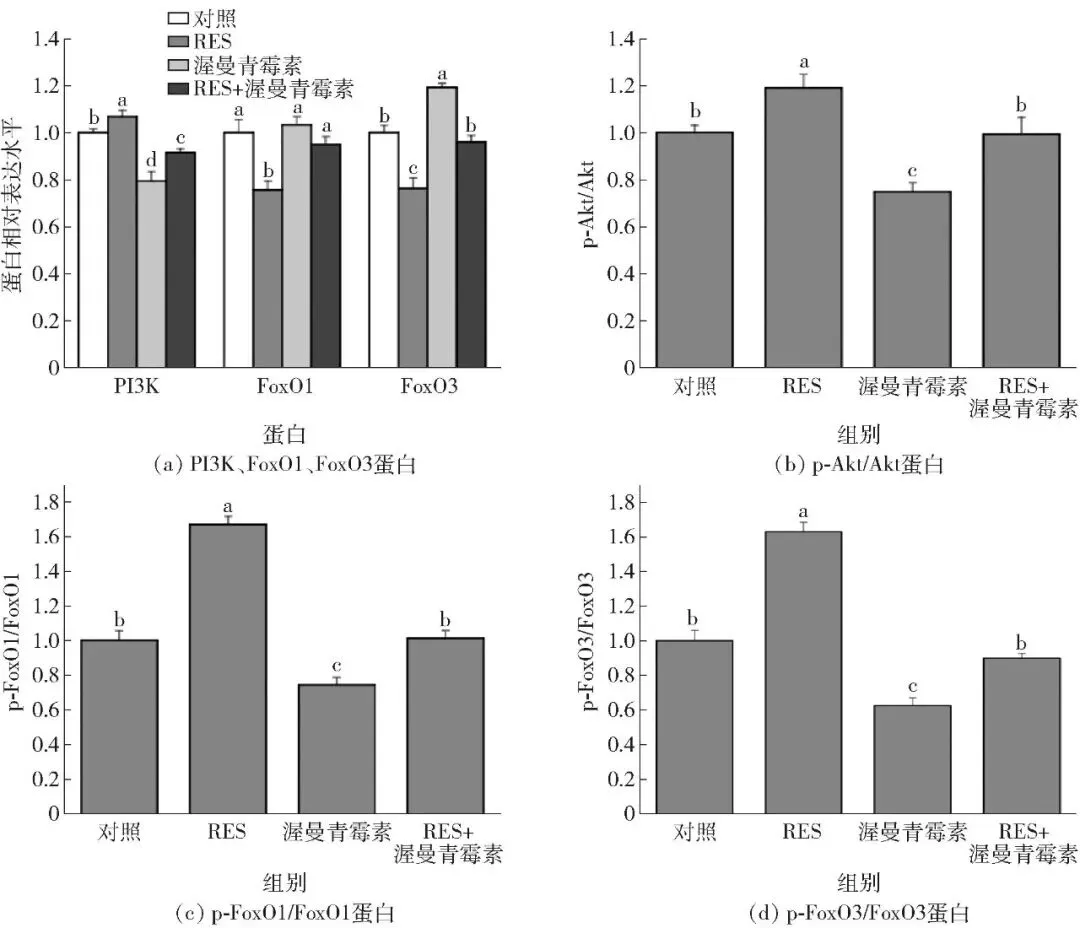
2.6.2 对白藜芦醇调控Akt/FoxO信号通路核心蛋白表达的影响
通路核心蛋白的表达结果见图11、图12。添加白藜芦醇显著提高了牛肌管细胞中PI3K、p-Akt/Akt、p-FoxO1/FoxO1和p-FoxO3/FoxO3的蛋白表达水平,而渥曼青霉素则抵消了白藜芦醇对PI3K、p-Akt/Akt、p-FoxO1/FoxO1和p-FoxO3/FoxO3蛋白表达的促进作用(P<0.05)。此外,白藜芦醇显著降低了牛肌管细胞中FoxO1和FoxO3的蛋白表达水平,渥曼青霉素减弱了白藜芦醇对FoxO1和FoxO3蛋白表达的抑制作用(P<0.05)。结果表明,渥曼青霉素阻断Akt/FoxO通路信号传导会促进FoxO1和FoxO3表达,从而影响牛肌管细胞肌纤维类型组成。
图11 渥曼青霉素处理牛肌管细胞后Akt/FoxO信号通路蛋白免疫印迹图Fig.11 Western blotting of Akt/FoxO signaling pathway proteins in bovine myotubes treated with wortmannin
不同字母表示不同处理间差异显著(P<0.05)。
图12 渥曼青霉素对牛肌管细胞中Akt/FoxO信号通路蛋白表达水平的影响Fig.12 Effects of wortmannin on protein expression levels of Akt/FoxO signaling pathway in bovine myotubes
本研究建立了氧化应激诱导的牛肌管细胞快肌化模型,确定400 μmol/L H2O2处理24 h为较佳建模条件,该条件在保证细胞存活率的同时,能有效诱导肌纤维类型由慢肌向快肌转化。浓度梯度实验表明,10 μmol/L RES的调控效果较优,可逆转H2O2诱导的慢肌基因表达抑制和快肌基因表达激活。确定了渥曼青霉素(1 μmol/L)和SC79(10 μmol/L)的干预剂量,为体内研究奠定剂量基础。研究证实,RES通过激活Akt/FoxO信号通路实现肌纤维类型的调控,mRNA水平检测显示,10 μmol/L RES可显著上调Akt/FoxO通路正调控因子(PI3Kα、PI3Kβ、Akt1、Akt2)及氧化代谢相关基因(MEF2C、PGC-1α)的表达,同时抑制负调控因子(FoxO1、FoxO3)的转录;H2O2处理可显著抑制PI3K/Akt通路活性、激活FoxO1/FoxO3,而RES与Akt激活剂SC79联用可完全恢复通路正常功能;渥曼青霉素干预则不仅削弱了RES对慢肌纤维的促进作用和对快肌纤维的抑制作用,还逆转了其对Akt/FoxO通路关键蛋白的调控效应,证明该通路是RES逆转氧化应激诱导快肌化的介导途径。然而,本研究仅基于细胞实验,其结果存在一定局限性。细胞模型难以完全模拟体内复杂的微环境及系统调控网络;此外,RES在体内的代谢稳定性、生物利用度及其对整体动物的潜在影响,仍需进一步探讨。后续研究有必要在动物模型中进一步验证RES对应激条件下肌纤维类型转化的调控机制,以全面评估其潜在应用价值。
逯莹,孙静,朱立贤,等. 白藜芦醇调控氧化应激诱导的牛肌管细胞快肌化的Akt/FoxO信号通路机制[J]. 食品科学技术学报,2026,44(2):161-172. LU Ying, SUN Jing, ZHU Lixian, et al. Akt/FoxO signaling pathway mechanism of resveratrol regulating oxidative stress-induced fast-twitch transformation in bovine myotubes[J]. Journal of Food Science and Technology, 2026,44(2):161-172.
国家自然科学基金面上项目(32572494、32072239);山东省牛产业创新团队项目(SDAIT-09-09);国家肉牛牦牛产业技术体系项目(CRAS-37)。
General Project of National Natural Science Foundation of China (32572494,32072239);Shandong Province Cattle Industry Innovation Team Project (SDAIT-09-09);China National Beef Cattle and Yak Industrial Technology System Project (CRAS-37).
制作:赵宇飞
编辑:李宁
审核:叶红波、张逸群







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
