Plant Commun | 南京农业大学常明和河南农业大学王燃团队揭示PUB13调控植物免疫与叶片寿命的分子机制
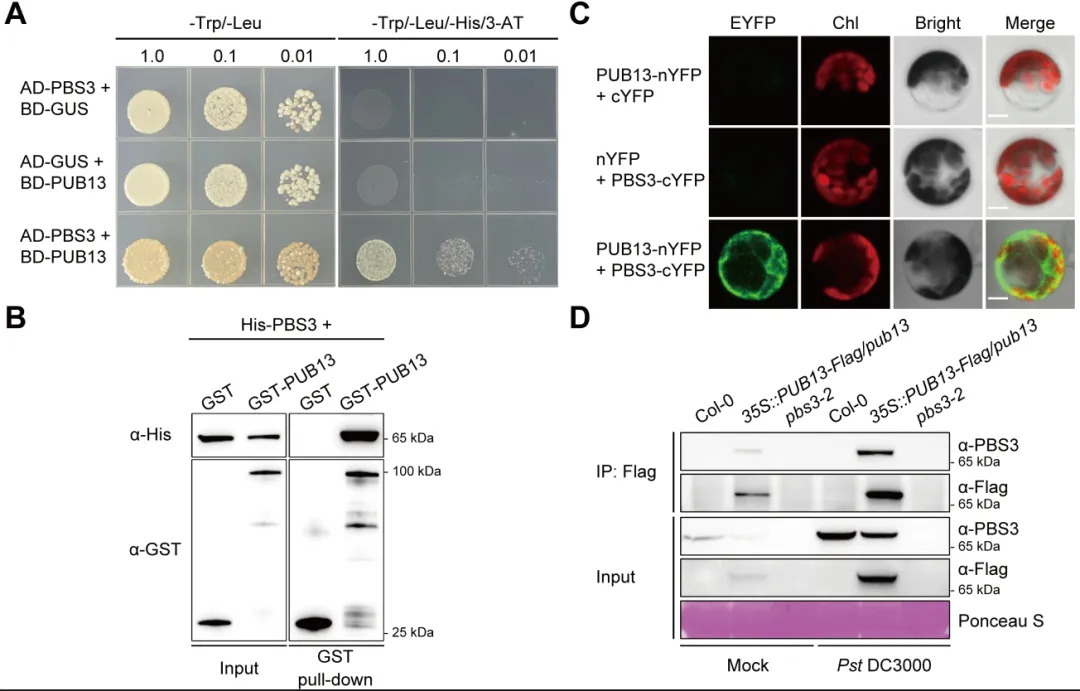
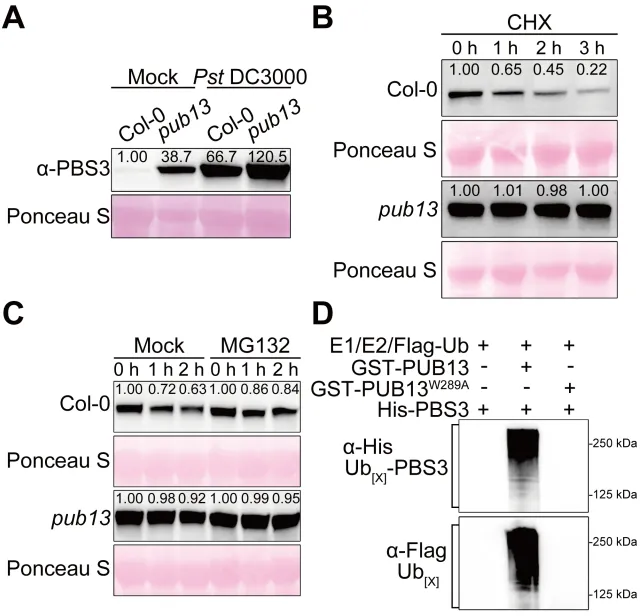
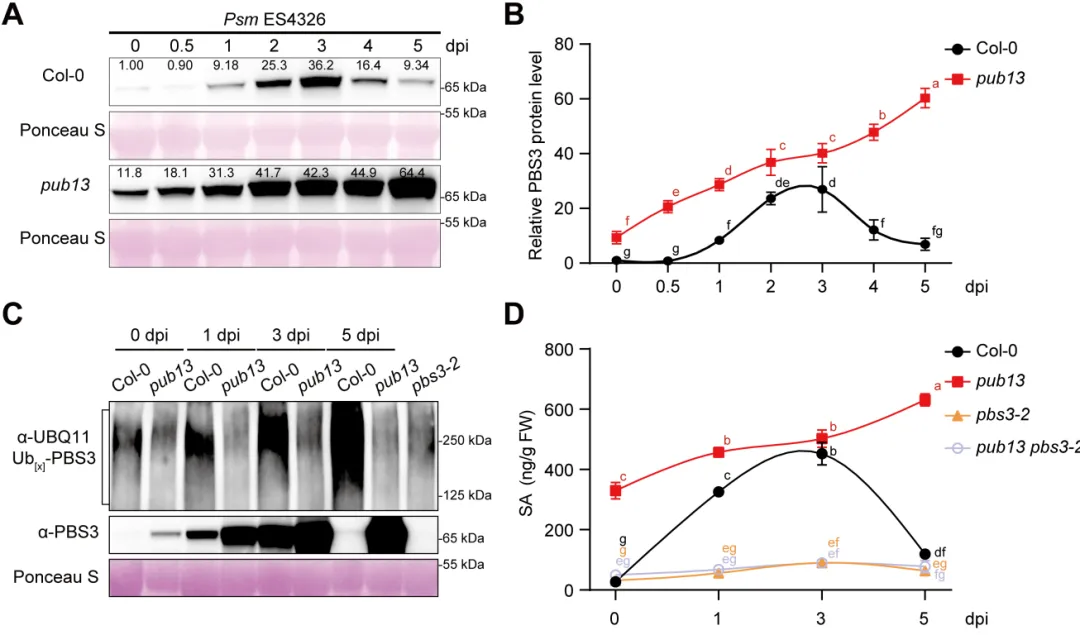
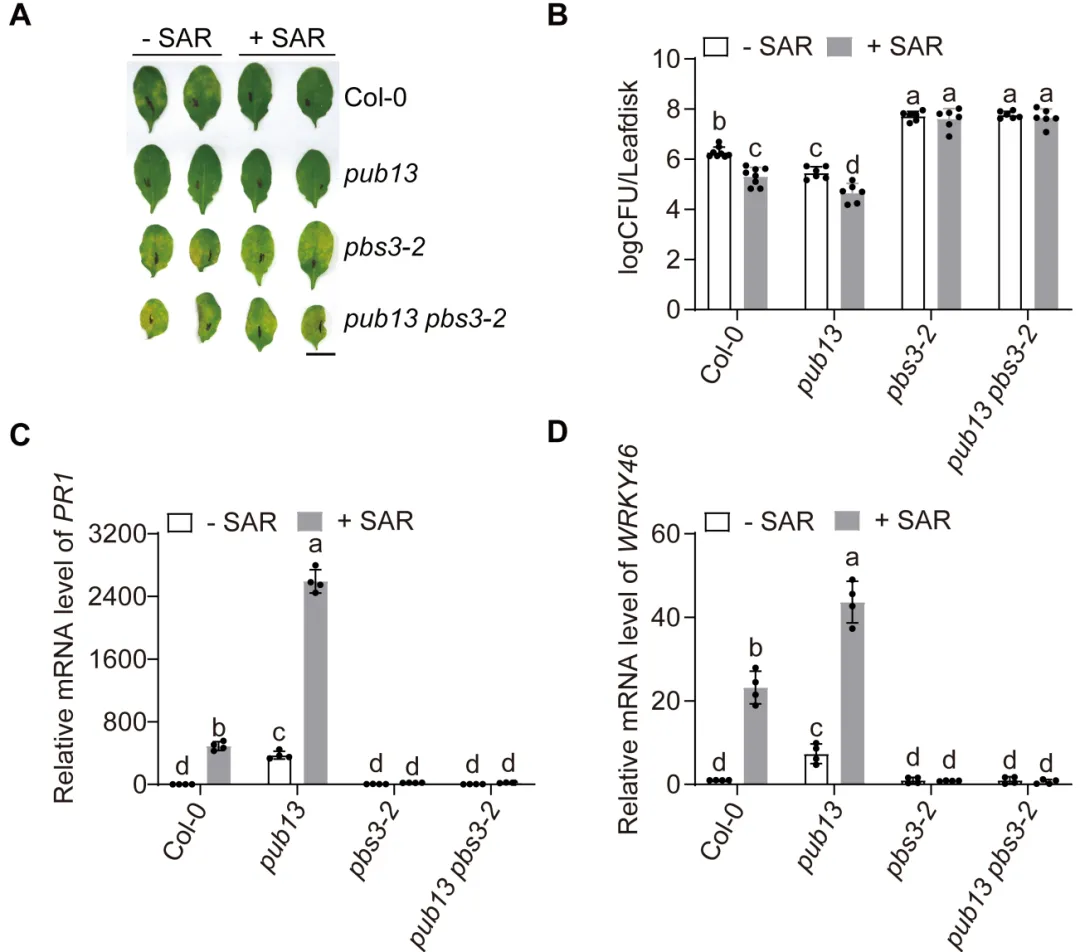
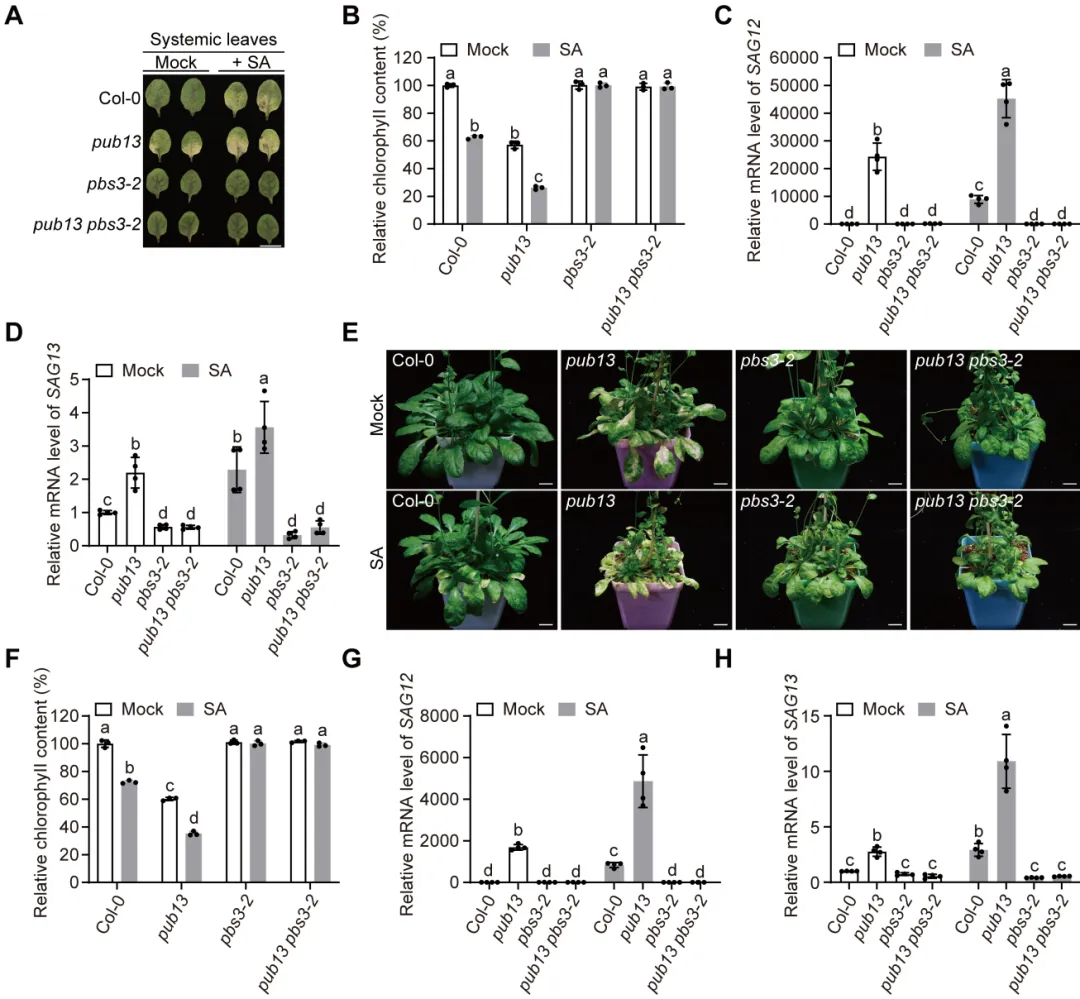
植物遭遇病原菌后需要迅速启动免疫,同时也必须及时关闭高强度防御,避免生长受阻和叶片早衰。水杨酸是连接这两类过程的核心激素:病原侵染会诱导水杨酸积累,激活局部防御和系统获得性抗性,但过量或持续的水杨酸也会促进衰老相关基因表达,缩短叶片寿命。对作物而言,防御持续过久会减少光合叶面积、缩短灌浆或营养生长时间,因此抗病与寿命之间的权衡直接影响田间产量和适应性。PBS3是异分支酸途径中合成水杨酸的关键酶,其转录调控已有较多研究,但PBS3蛋白如何被动态清除、水杨酸高峰如何在免疫后期回落,以及植物怎样在抗病收益与发育代价之间实现平衡,仍缺少明确的分子解释。2026年6月17日,南京农业大学生命科学学院常明教授团队和河南农业大学生命科学学院王燃教授团队在知名期刊《Plant Communications》发表了题为“PUB13-Mediated Degradation of PBS3 Regulates Salicylic Acid Biosynthesis to Coordinate Plant Immunity and Leaf Longevity”的研究论文。研究通过遗传学、生物化学和生理分析发现,E3泛素连接酶PUB13直接识别水杨酸合成酶PBS3,促进其多聚泛素化及26S蛋白酶体降解。该过程在免疫启动后逐渐限制PBS3积累,使水杨酸及时从峰值回落,从而既维持有效的系统获得性抗性,又避免持续免疫造成叶片过早衰老。研究表明,PUB13-PBS3 模块是一种精细调控 SA 生物合成的翻译后机制,它既能确保有效的免疫反应,又能防止 SA 的过度积累及其对植物寿命的不利影响。研究构建了3个PBS3过表达株系和2个互补株系,发现PBS3转录量、蛋白积累和水杨酸含量呈梯度升高。以Psm ES4326-AvrRpt2诱导系统获得性抗性后,pbs3突变体抗病性下降,PBS3过表达植株抗性和PR1表达增强,而且免疫输出随PBS3丰度逐级增加。6周龄时,过表达株系叶片黄化加快、叶绿素下降,SAG12和SAG13表达升高,外源水杨酸使表型进一步加重;pbs3突变体则延迟衰老。单因素方差分析及Tukey检验显示差异显著,说明PBS3是决定免疫强度与叶片寿命的定量调节因子。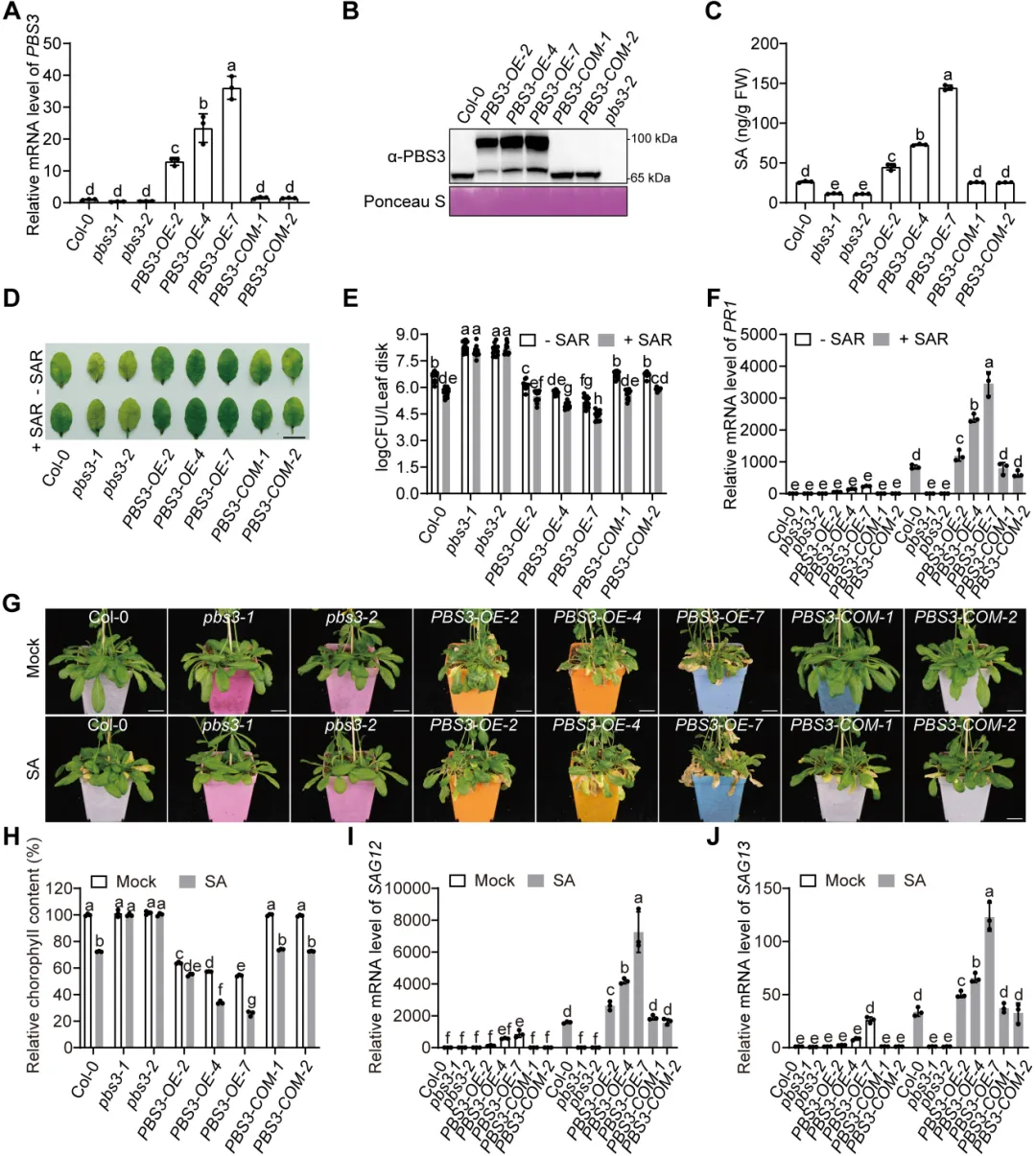 图1:PBS3丰度剂量依赖性调控免疫和叶片衰老以PBS3为诱饵的酵母双杂交筛选鉴定到E3泛素连接酶PUB13,而近缘蛋白PUB12和PUB14不能结合PBS3,PUB13也不结合GH3.7、GH3.15、GH3.17以及ICS1、EPS1,显示出较高底物特异性。结构域分析显示PBS3结合PUB13的ARM重复区域。GST pull-down证实两者可在体外直接结合,双分子荧光互补在拟南芥原生质体中产生明显EYFP信号,共免疫沉淀则在植株中检出复合物;Pst DC3000侵染后相互作用进一步增强,表明免疫激活时积累的PBS3更容易被PUB13识别。图2:PUB13特异且直接结合水杨酸合成酶PBS3pub13突变体在正常及病原侵染条件下均积累更多PBS3,而pub12没有相同变化。在瞬时表达体系中,两种基因型的PBS3转录水平相近,但pub13原生质体中的PBS3蛋白明显升高,证明调控发生在转录后。环己酰亚胺追踪显示PBS3在野生型中快速下降,在pub13中基本稳定;蛋白酶体抑制剂MG132可阻断野生型中的降解。体外反应加入E1、E2、ATP和泛素后,PUB13能够催化PBS3多聚泛素化,说明PUB13通过泛素-26S蛋白酶体系统控制PBS3稳定性。 图3:PUB13介导PBS3多聚泛素化与蛋白酶体降解PUB13表达会被病原侵染和叶片衰老诱导。与野生型相比,pub13突变体水杨酸水平更高、系统获得性抗性和PR1表达更强,同时叶片衰老加速;3个PUB13过表达株系则随PUB13表达量升高而逐步降低水杨酸和抗病输出,并表现出延迟衰老和更长叶片寿命。相关指标采用单因素方差分析和Tukey检验比较,呈现一致的表达剂量效应,说明PUB13负向调节水杨酸积累和系统免疫,却正向维持叶片寿命。时间序列分析显示,Psm ES4326侵染后,野生型PBS3蛋白迅速升高,在约3天达到峰值,随后下降;pub13在侵染前即具有较高PBS3,并在0至5天持续积累。野生型PBS3多聚泛素化随侵染进程增强,pub13中则明显减弱。水杨酸变化与PBS3动态高度一致:野生型先快速升高再回落,pub13保持高水平;pbs3-2及pub13 pbs3-2双突变体几乎不能产生病原诱导的水杨酸。双因素方差分析表明差异显著,说明PUB13在免疫后期加速PBS3清除,是水杨酸结束持续积累的关键机制。图4 PUB13动态限制PBS3并促进病原诱导水杨酸回落遗传分析进一步检验PUB13是否通过PBS3调节系统获得性抗性。pub13表现出增强的系统抗性,pbs3-2则明显受损;pub13 pbs3-2双突变体完全失去pub13的增强表型,并与pbs3-2相似。系统抗性诱导后,pub13中的PBS3积累以及PR1、WRKY46表达均高于野生型,而这些响应在pbs3-2和双突变体中大幅减弱。细菌生长量和基因表达经单因素方差分析及Tukey检验显示显著差异,证明PBS3是PUB13调控系统免疫的主要下游组分。图5 PUB13通过限制PBS3积累精细调节系统获得性抗性研究团队在诱导系统抗性后以100微摩尔水杨酸处理系统叶片,观察免疫相关衰老。野生型出现中度黄化,pub13叶片衰老明显加重,而pbs3-2和pub13 pbs3-2基本保持绿色。叶绿素测定与外观一致,pub13中的SAG12和SAG13表达显著升高,双突变体则保持低水平。更重要的是,去除PBS3可完全抑制pub13的过度衰老,且PBS3蛋白积累量与衰老严重程度正相关,说明PUB13通过周转PBS3来防止免疫激活后水杨酸信号过强地推动叶片衰老。在正常年龄依赖性衰老和外源水杨酸处理条件下,6周龄pub13植株均表现出明显早衰,pbs3-2接近野生型,而pub13 pbs3-2双突变体更接近pbs3-2。pub13叶绿素损失最严重,SAG12和SAG13在正常及水杨酸处理条件下均强烈诱导;pbs3-2和双突变体中的表达维持较低。单因素方差分析和Tukey检验支持这些差异,由此建立明确的遗传层级:PBS3位于PUB13下游,是PUB13连接水杨酸合成、免疫输出与叶片寿命的关键功能靶点。图6 PBS3在PUB13下游介导水杨酸依赖的叶片衰老研究揭示了植物免疫不仅需要快速“打开”,也需要精准“收尾”。静息状态下,PUB13持续限制PBS3和基础水杨酸水平;病原侵染后,PBS3快速积累并推动水杨酸爆发,激活防御的同时也提高衰老风险;免疫进入后期,PUB13增强PBS3多聚泛素化和蛋白酶体降解,促使水杨酸回落并恢复生长稳态。有意思的是,PUB13由此把早期病原识别与后期激素代谢控制连接起来,形成兼顾防御强度和持续时间的稳态开关。该工作补充了PBS3蛋白周转这一长期缺失的调控环节,也为在不牺牲叶片寿命的前提下优化作物抗病性提供了潜在分子靶点。原文链接:https://doi.org/10.1016/j.xplc.2026.101973
图1:PBS3丰度剂量依赖性调控免疫和叶片衰老以PBS3为诱饵的酵母双杂交筛选鉴定到E3泛素连接酶PUB13,而近缘蛋白PUB12和PUB14不能结合PBS3,PUB13也不结合GH3.7、GH3.15、GH3.17以及ICS1、EPS1,显示出较高底物特异性。结构域分析显示PBS3结合PUB13的ARM重复区域。GST pull-down证实两者可在体外直接结合,双分子荧光互补在拟南芥原生质体中产生明显EYFP信号,共免疫沉淀则在植株中检出复合物;Pst DC3000侵染后相互作用进一步增强,表明免疫激活时积累的PBS3更容易被PUB13识别。图2:PUB13特异且直接结合水杨酸合成酶PBS3pub13突变体在正常及病原侵染条件下均积累更多PBS3,而pub12没有相同变化。在瞬时表达体系中,两种基因型的PBS3转录水平相近,但pub13原生质体中的PBS3蛋白明显升高,证明调控发生在转录后。环己酰亚胺追踪显示PBS3在野生型中快速下降,在pub13中基本稳定;蛋白酶体抑制剂MG132可阻断野生型中的降解。体外反应加入E1、E2、ATP和泛素后,PUB13能够催化PBS3多聚泛素化,说明PUB13通过泛素-26S蛋白酶体系统控制PBS3稳定性。 图3:PUB13介导PBS3多聚泛素化与蛋白酶体降解PUB13表达会被病原侵染和叶片衰老诱导。与野生型相比,pub13突变体水杨酸水平更高、系统获得性抗性和PR1表达更强,同时叶片衰老加速;3个PUB13过表达株系则随PUB13表达量升高而逐步降低水杨酸和抗病输出,并表现出延迟衰老和更长叶片寿命。相关指标采用单因素方差分析和Tukey检验比较,呈现一致的表达剂量效应,说明PUB13负向调节水杨酸积累和系统免疫,却正向维持叶片寿命。时间序列分析显示,Psm ES4326侵染后,野生型PBS3蛋白迅速升高,在约3天达到峰值,随后下降;pub13在侵染前即具有较高PBS3,并在0至5天持续积累。野生型PBS3多聚泛素化随侵染进程增强,pub13中则明显减弱。水杨酸变化与PBS3动态高度一致:野生型先快速升高再回落,pub13保持高水平;pbs3-2及pub13 pbs3-2双突变体几乎不能产生病原诱导的水杨酸。双因素方差分析表明差异显著,说明PUB13在免疫后期加速PBS3清除,是水杨酸结束持续积累的关键机制。图4 PUB13动态限制PBS3并促进病原诱导水杨酸回落遗传分析进一步检验PUB13是否通过PBS3调节系统获得性抗性。pub13表现出增强的系统抗性,pbs3-2则明显受损;pub13 pbs3-2双突变体完全失去pub13的增强表型,并与pbs3-2相似。系统抗性诱导后,pub13中的PBS3积累以及PR1、WRKY46表达均高于野生型,而这些响应在pbs3-2和双突变体中大幅减弱。细菌生长量和基因表达经单因素方差分析及Tukey检验显示显著差异,证明PBS3是PUB13调控系统免疫的主要下游组分。图5 PUB13通过限制PBS3积累精细调节系统获得性抗性研究团队在诱导系统抗性后以100微摩尔水杨酸处理系统叶片,观察免疫相关衰老。野生型出现中度黄化,pub13叶片衰老明显加重,而pbs3-2和pub13 pbs3-2基本保持绿色。叶绿素测定与外观一致,pub13中的SAG12和SAG13表达显著升高,双突变体则保持低水平。更重要的是,去除PBS3可完全抑制pub13的过度衰老,且PBS3蛋白积累量与衰老严重程度正相关,说明PUB13通过周转PBS3来防止免疫激活后水杨酸信号过强地推动叶片衰老。在正常年龄依赖性衰老和外源水杨酸处理条件下,6周龄pub13植株均表现出明显早衰,pbs3-2接近野生型,而pub13 pbs3-2双突变体更接近pbs3-2。pub13叶绿素损失最严重,SAG12和SAG13在正常及水杨酸处理条件下均强烈诱导;pbs3-2和双突变体中的表达维持较低。单因素方差分析和Tukey检验支持这些差异,由此建立明确的遗传层级:PBS3位于PUB13下游,是PUB13连接水杨酸合成、免疫输出与叶片寿命的关键功能靶点。图6 PBS3在PUB13下游介导水杨酸依赖的叶片衰老研究揭示了植物免疫不仅需要快速“打开”,也需要精准“收尾”。静息状态下,PUB13持续限制PBS3和基础水杨酸水平;病原侵染后,PBS3快速积累并推动水杨酸爆发,激活防御的同时也提高衰老风险;免疫进入后期,PUB13增强PBS3多聚泛素化和蛋白酶体降解,促使水杨酸回落并恢复生长稳态。有意思的是,PUB13由此把早期病原识别与后期激素代谢控制连接起来,形成兼顾防御强度和持续时间的稳态开关。该工作补充了PBS3蛋白周转这一长期缺失的调控环节,也为在不牺牲叶片寿命的前提下优化作物抗病性提供了潜在分子靶点。原文链接:https://doi.org/10.1016/j.xplc.2026.101973

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
