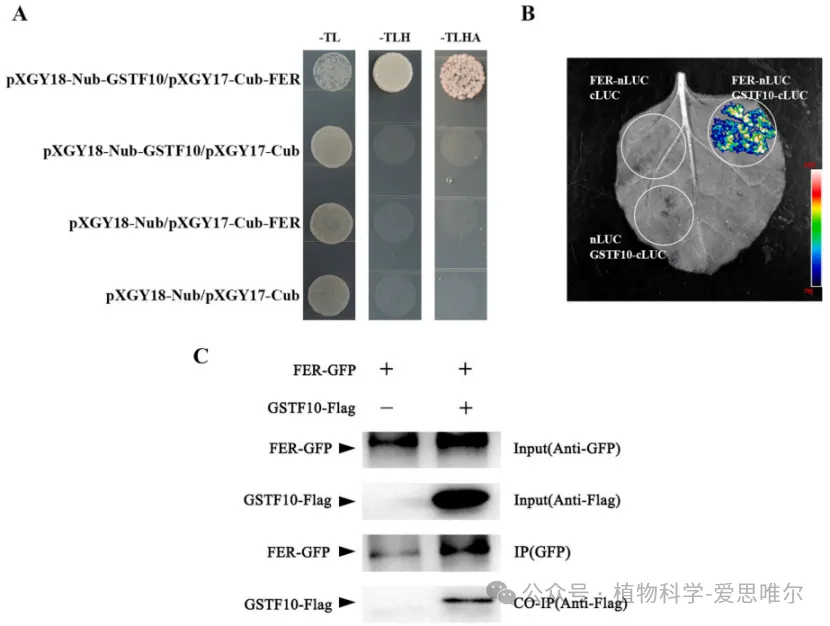
为了鉴定FER信号通路的潜在组分,研究者通过免疫沉淀-质谱学(IP-MS)筛选发现GSTF10为FER的候选互作蛋白。随后通过酵母双杂交、萤光素酶互补实验和免疫共沉淀三种方法,共同证实FER与GSTF10之间存在直接的物理相互作用,提示GSTF10可能通过与FER互作参与拟南芥生殖发育。
2.GSTF10在柱头乳头细胞中高表达,抑制花粉管伸长和结实
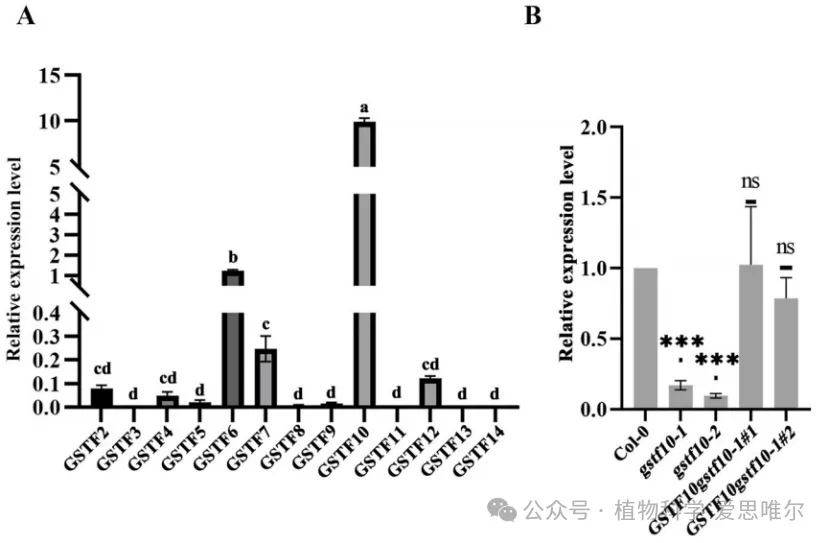
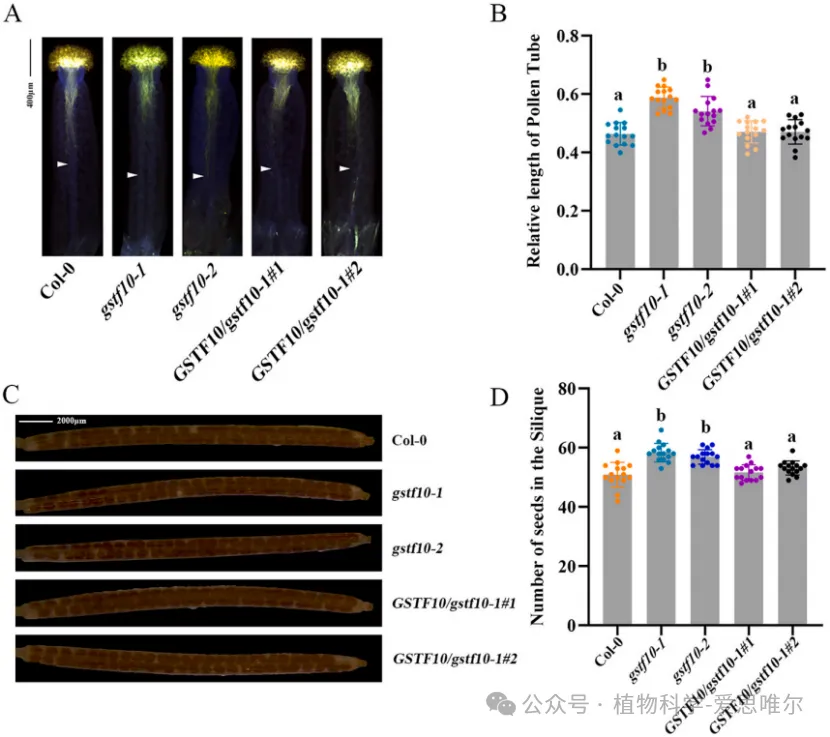
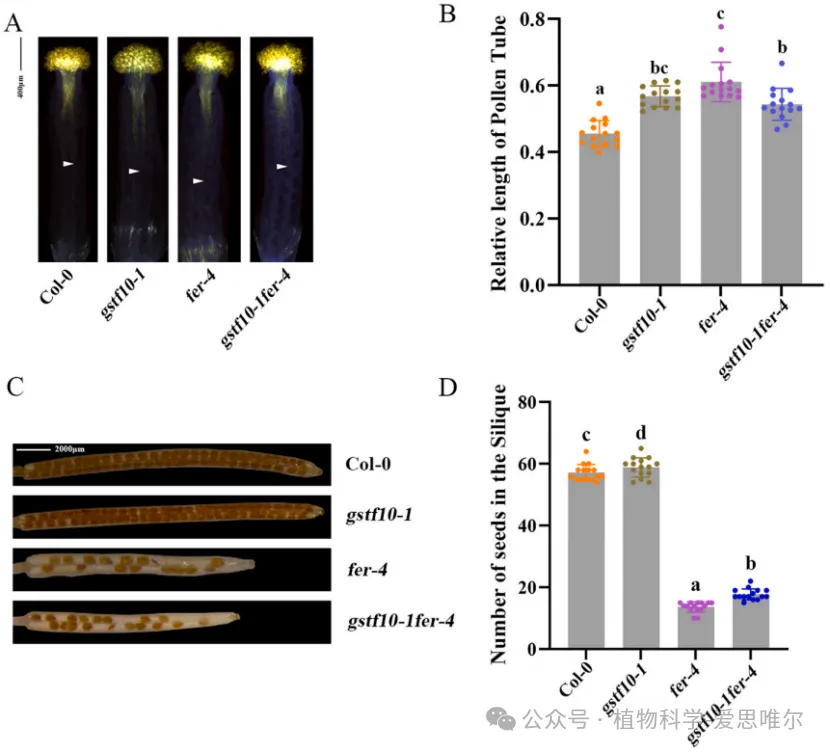
为探究Phi类谷胱甘肽S-转移酶(GSTF)家族与生殖发育的关系,研究者首先通过qRTPCR检测拟南芥13个GSTF基因在柱头乳突细胞中的表达,发现GSTF10在乳突细胞中表达水平显著高于其他GSTF成员,提示其与亲和性花粉柱头互作密切相关。为解析GSTF10体内功能,研究选用并鉴定了两个独立T-DNA插入缺失突变体:gstf10-1和gstf10-2。观察gstf10-1和gstf10-2突变体,发现它们柱头上花粉管明显更长。另外,gstf10每角果种子数显著多于野生型。这就说明GSTF10负调控花粉管在柱头内的伸长,并负调控胚珠/种子数目。
3. GSTF10通过柱头乳突ROS动态平衡调节花粉水合作用
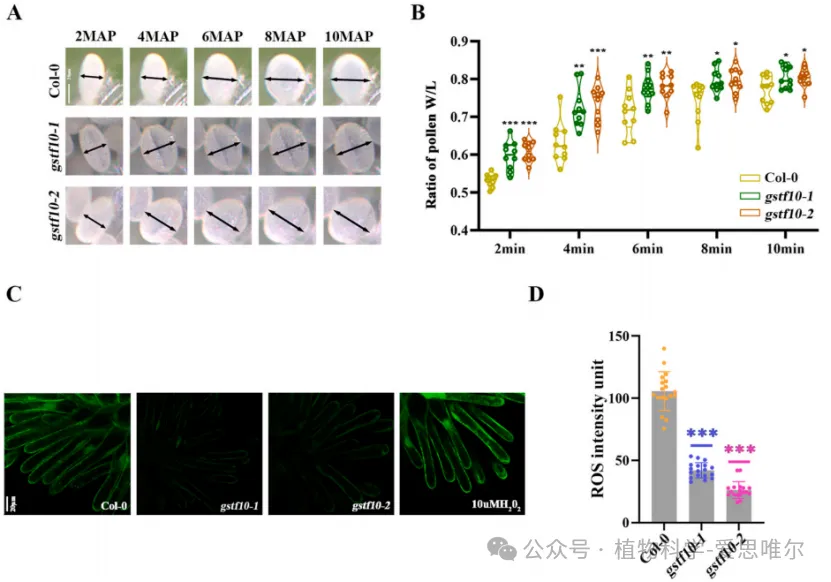
为探究gstf10突变体中花粉管生长加速的机制,作者检测了花粉水合速率和柱头ROS水平。结果显示,gstf10突变体柱头上的花粉水合速度显著快于野生型,同时其柱头乳突细胞中的ROS水平显著低于野生型。此外,对拟南芥RNA-seq数据库(ARS)的分析表明,GSTF10在生长中的花粉管中也高度表达。这些结果表明,GSTF10通过维持柱头ROS水平来延迟花粉水合,从而负调控花粉管生长,并可能直接作用于花粉管限制其伸长。
4. GSTF10位于FER下游调节花粉管生长和结实
为明确GSTF10与FER之间的遗传关系,构建了gstf10-1 fer-4双突变体。在花粉管长度方面,双突变体表型与gstf10-1相似,均显著短于fer-4。在结实方面,fer-4单突变体种子数严重减少,而双突变体介于fer-4与野生型之间,呈现部分恢复。这表明GSTF10在遗传上位于FER下游:FER可能通过促进GSTF10的功能来抑制花粉管伸长,同时通过抑制GSTF10的功能来促进结实。
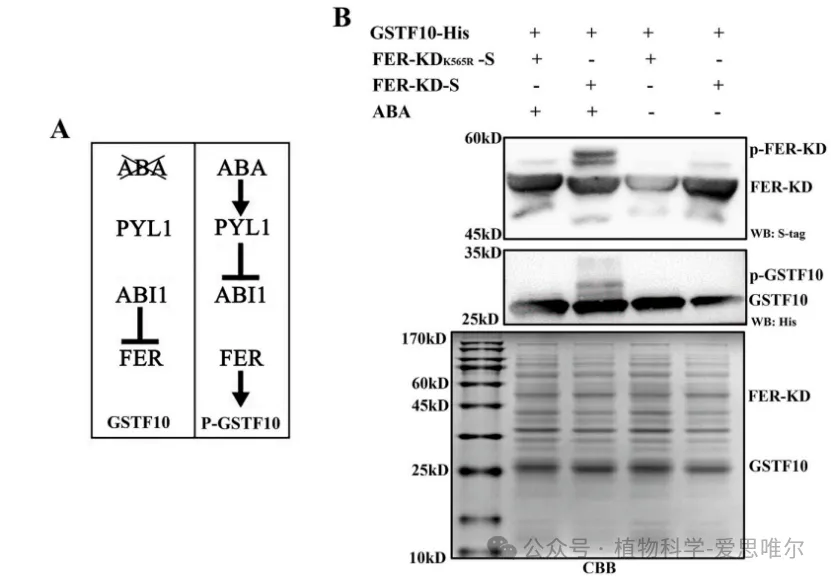
利用ABA诱导的大肠杆菌共表达系统,同时表达PYL1、ABI1、FER激酶结构域(FER-KD)及GSTF10。加入ABA后,FER-KD与GSTF10均出现磷酸化条带;而以激酶失活突变体FER-KDK565R替代FER-KD后,GSTF10未被磷酸化。说明FER激酶可在体外直接磷酸化GSTF10,且该过程依赖FER的激酶活性。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
