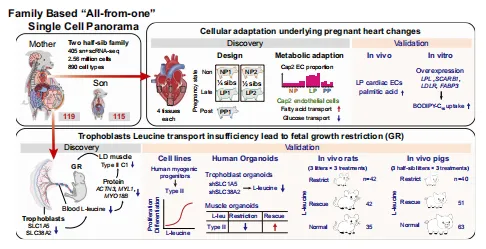
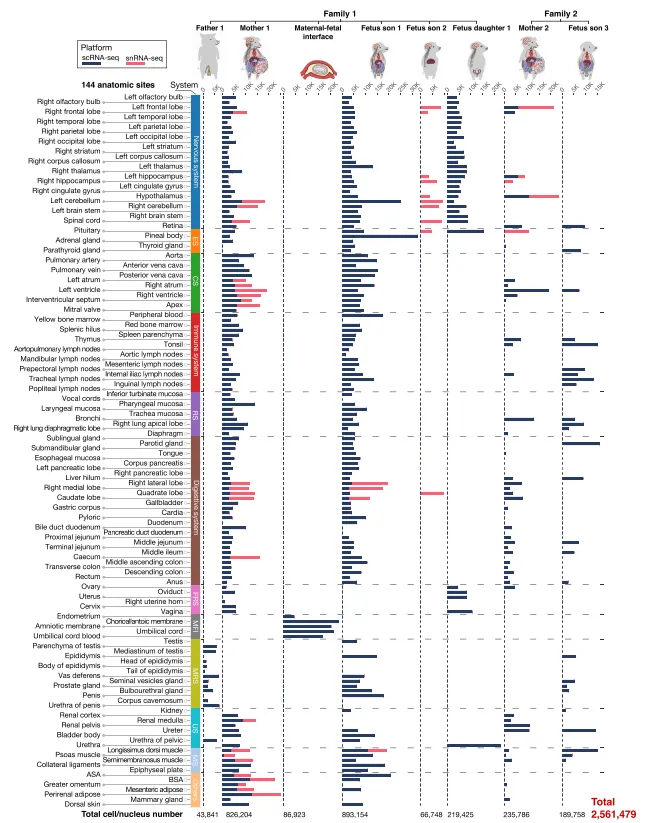
一共从一头怀孕母猪和它的胎儿身上采集了115个组织,拿到了256万个细胞,分出了76种主要细胞类型和412种内皮亚型,这就是(图1)。
图1.来自两个猪科物种的“一体化”单细胞全景图概览
💡Tips:采用“同一家庭、同一时间”的大规模组织解离与单细胞测序,最大优势是消除了遗传、年龄和环境差异,使细胞类型和转录因子的比较极为纯净。
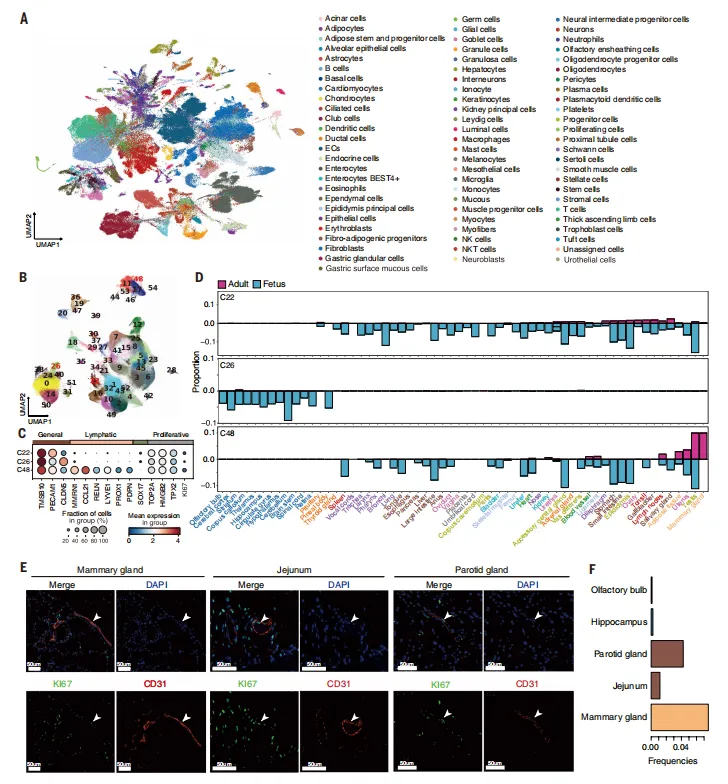
通过全局UMAP和内皮细胞整合分析,展示了76种主要细胞类型,并揭示了增殖性内皮细胞在脑与外周组织间的差异分布及脑特异性增殖性内皮的存在(图2)。
图2.“一体化”单细胞图谱的构建及胎儿与成年猪全身器官内EC分布特征的表征
💡该方法通过降采样UMAP可视化和大规模内皮细胞整合,优点是能在单一个体背景下清晰揭示组织特异性的增殖内皮分布,但降采样可能掩盖稀有细胞亚群,且需要上百个样本的协同处理,复现门槛极高。
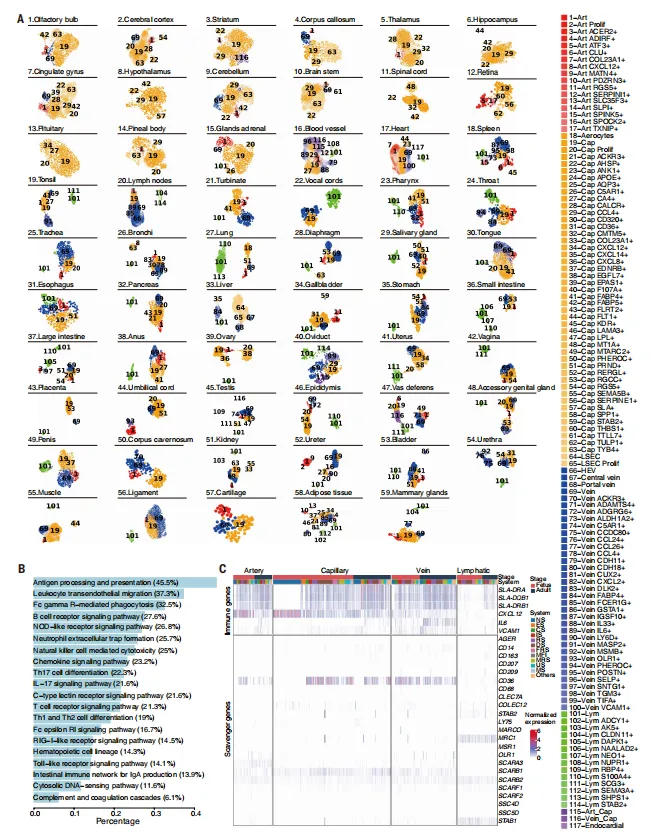
内皮细胞在脑与外周器官间呈现增殖、免疫和代谢差异。KEGG免疫通路基因在412个EC亚型中广泛表达,清道夫受体基因在血管类型、器官和发育阶段间表达异质(图3)。
图3.59个器官中内皮细胞(ECs)的器官内异质性及免疫调节功能多样性
💡方法上依赖高质量的单细胞注释和大量已知标记基因,可复现性较好,但清道夫受体功能验证仍需后续实验。
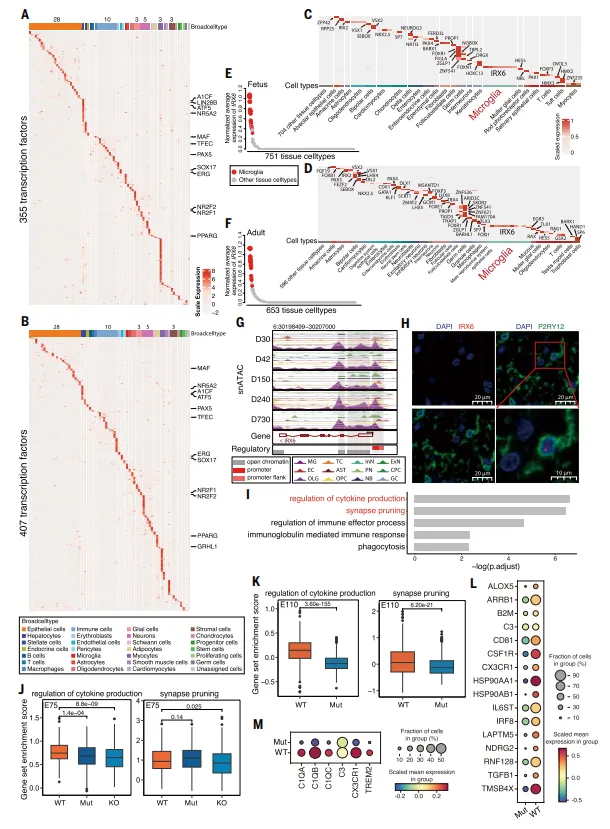
在胎儿和成年中分别鉴定出355和407个高特异性TF,以及32和50个组织细胞类型特异性TF。其中IRX6为小胶质细胞特异性TF,在11个脑区保守表达,并经基因编辑猪验证(图4)。
💡优点:验证手段扎实,局限性在于IRX6在人类中是否完全保守尚需跨物种验证。
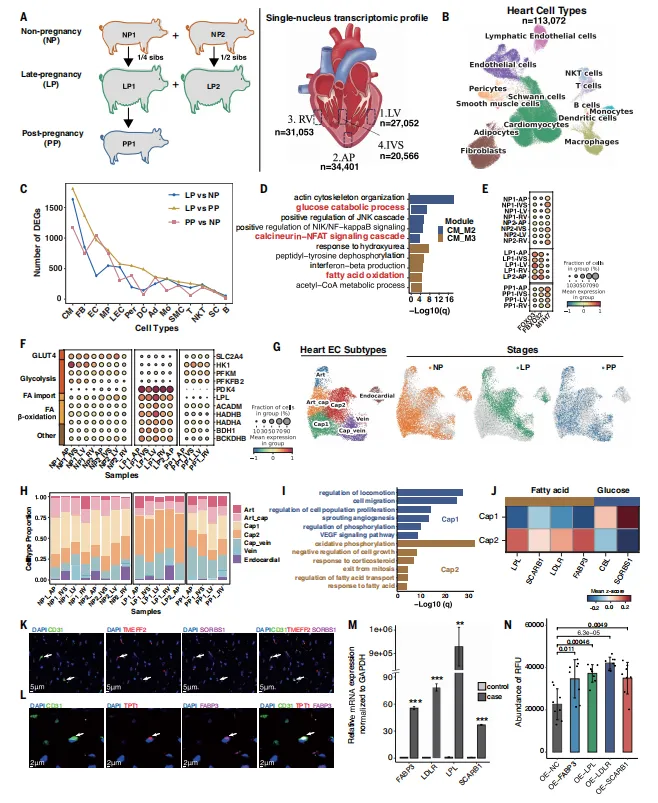
晚孕期心肌细胞上调FOXO3抗肥大通路和脂肪酸氧化基因,下调葡萄糖代谢基因。心脏中出现妊娠富集的毛细血管内皮亚型,高表达脂肪酸转运基因,其丰度在产后恢复。体外过表达这些基因可增强脂肪酸摄取(图5)。
图5.猪心脏在NP、LP和PP阶段的动态细胞及功能适应性变化
💡采用半同胞设计(非孕期、晚孕期、产后)联合snRNA-seq、hdWGCNA、代谢组及PIEC细胞功能实验,系统揭示了心脏适应性重塑。
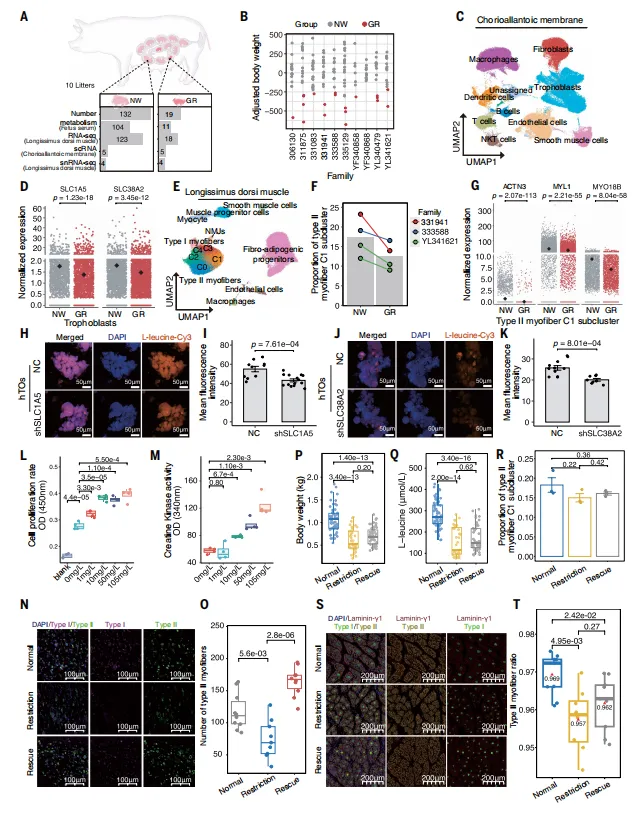
FGR胎儿绒毛膜尿囊膜滋养层中氨基酸转运体SLC1A5和SLC38A2表达降低,导致胎儿血清L-亮氨酸减少。肌肉单核图谱显示FGR胎儿缺失一种特异的II型肌纤维亚型(IIC1)。在滋养层类器官、肌肉类器官及孕期大鼠/猪体内实验中,补充L-亮氨酸可部分恢复肌纤维形成和胎儿体重(图6)。
💡因果链条完整(转运体下调→血清亮氨酸下降→肌纤维缺失→体重降低)。
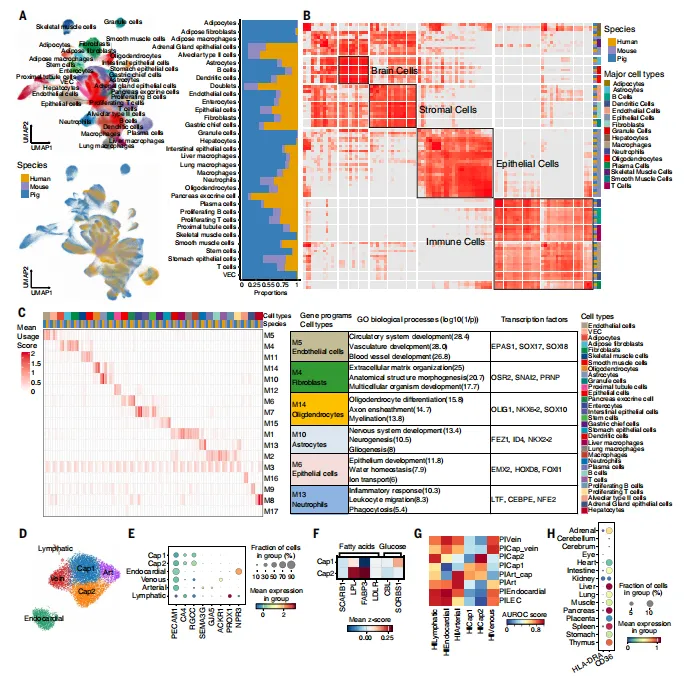
整合人、小鼠、猪15种共有组织的单细胞数据,发现免疫、上皮、基质和脑细胞类型高度保守。人类心脏中也存在Cap2亚型,且表达特征与猪相似;人类胎盘中SLC1A5/SLC38A2高表达;人类胎儿肌肉中存在与猪IIC1亚型转录相似的古纤维亚群(图7)。
图7.比较分析表明猪、人类和小鼠之间的细胞类型及基因表达程序存在跨物种保守性
💡注意:利用MetaNeighbor和cNMF对三个物种的公共数据进行整合分析,思路是验证猪模型的人类转化价值。优点是数据重用和跨物种基准分析,方法标准化程度高,较易复现。









 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
