《食品科学》:四川农业大学惠腾副教授等:热加工食品中四氢-β-咔啉的形成及其检测方法
- 2026-05-31 23:58:15


在食品加工领域,热加工技术凭借其能改善食品口感和延长保质期等优势,被广泛应用于面包、薯片和烤肉等众多食品的生产中。然而,食品在热加工过程中可形成多种有害化学物质,对食品安全构成潜在威胁。其中,四氢-β-咔啉(THβC)容易在食品加工过程中形成,具有不良活性,特别是具有致癌、致畸、致突变等毒性,会对人体健康造成不良影响,近年来逐渐成为食品安全研究的热点之一。
目前,研究表明,食品加工过程中,美拉德反应和微生物代谢是THβC形成的主要途径,时间、温度、pH值等反应条件影响其形成。但是,对于食品加工过程中THβC研究较少,形成机理不明,系统影响规律不清,检测方法还有待进一步研究。因此,四川农业大学食品学院的曹佳敏、李雪辰、惠腾*等人概括THβC形成的内源性与外源性途径及影响因素和食品中THβC的检测方法,以期为控制食品加工中THβC形成提供参考。

1


TTHHβC形成与影响因素

1.1 THβC概念
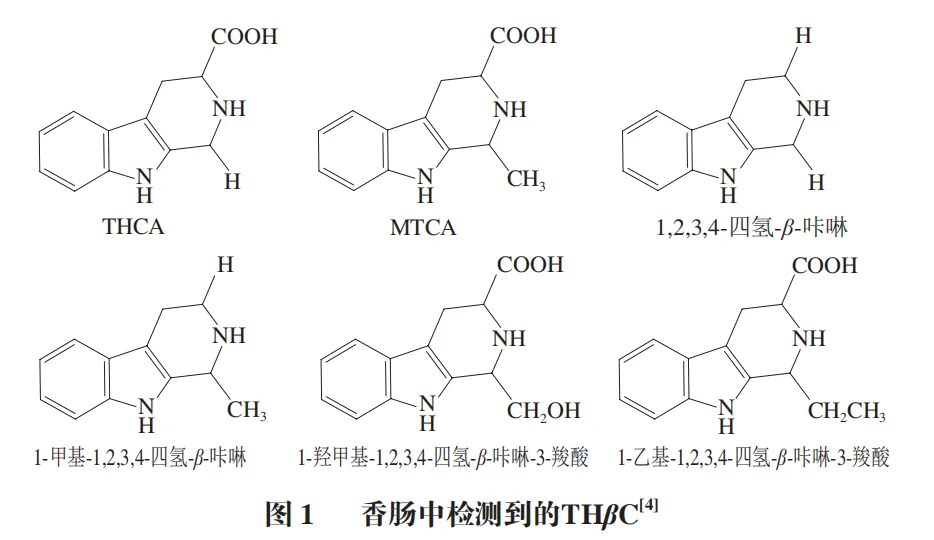
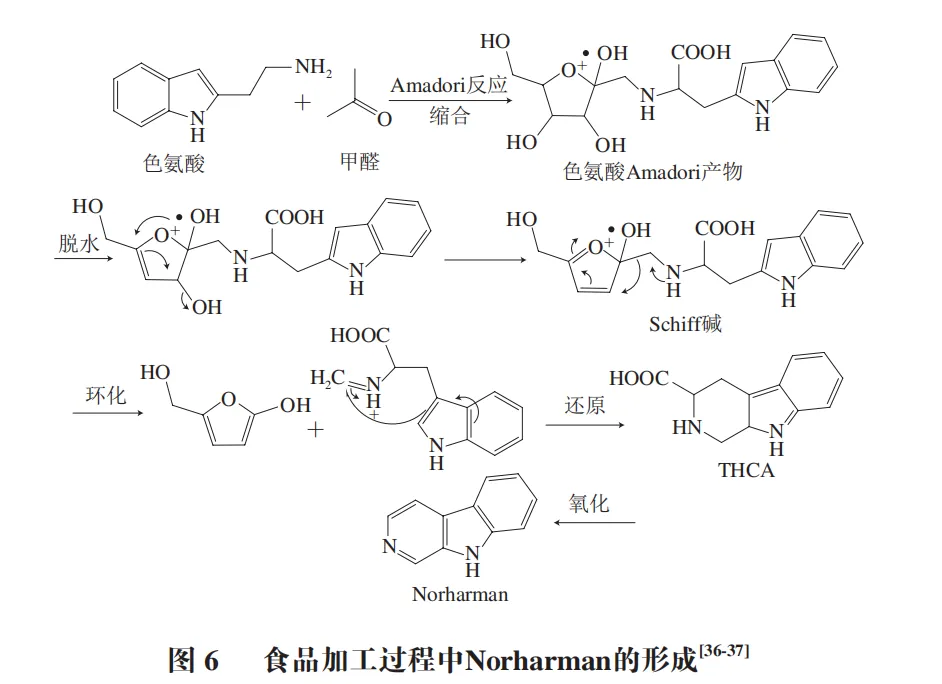
THβC是由吲哚胺和醛或α-酮酸通过Pictet-Spengler反应生成的具有三环吡啶[3,4-b]-吲哚环结构的一类生物碱。THβC广泛存在于动植物组织中,更是食品加工过程中形成的重要产物。目前在猪牛羊等畜禽肉中检测到多种THβC。食物中的THβC以1,2,3,4-四氢-β-咔啉-3-羧酸(THCA)和1-甲基-1,2,3,4-四氢-β-咔啉-3-羧酸(MTCA)为主。Herraiz等在香肠制品中检测到多种THβC(图1)。
1.2 外源形成途径
食品加工过程中容易形成THβC,主要涉及美拉德反应和微生物代谢两个重要途径。最终THβC随着食物的摄入进入人体。
1.2.1 美拉德反应
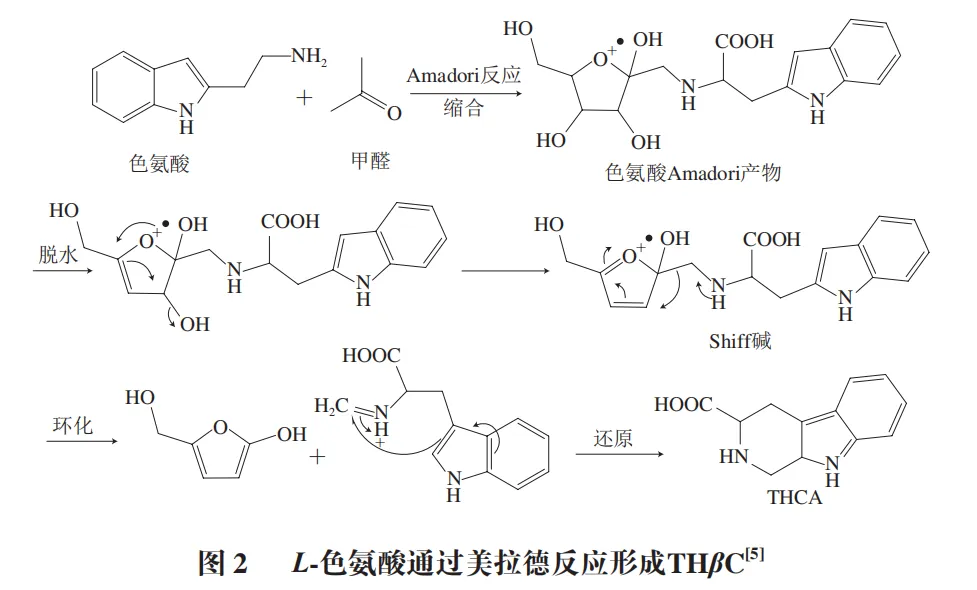
在120~200 ℃高温下,THβC反应机制见图2,主要分为3 步:缩合、环化和还原。色氨酸吲哚环上的氮原子(α-氨基)与醛类的羰基(—CHO)发生缩合,生成Amadori重排产物,脱水后生成Schiff碱(亚胺中间体)。Schiff碱在酸性或高温条件下发生分子内环化,吲哚环与亚胺键结合形成β-咔啉的六元环骨架(含氮杂环结构)。环化后的中间体在还原剂,如食品中的还原糖、VC或催化条件下,双键被还原形成饱和的THβC(图2)。
1.2.2 微生物代谢
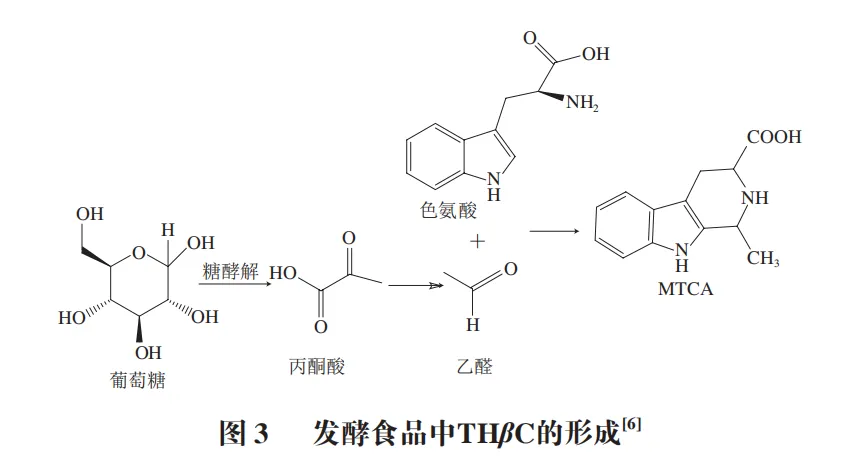
发酵食品中的THβC主要来自微生物代谢,不需要高温反应条件。在厌氧条件下,酵母菌通过糖酵解过程将葡萄糖转化为丙酮酸,丙酮酸进一步脱羧形成乙醛,乙醛可与色氨酸代谢产物结合生成THβC(图3)。例如在酱油发酵过程中,米曲霉分泌的蛋白酶分解大豆蛋白释放色氨酸,同时酵母菌通过糖酵解代谢产生乙醛,二者在发酵后期发生缩合反应,导致THβC含量增加。
1.2.3 内源形成途径
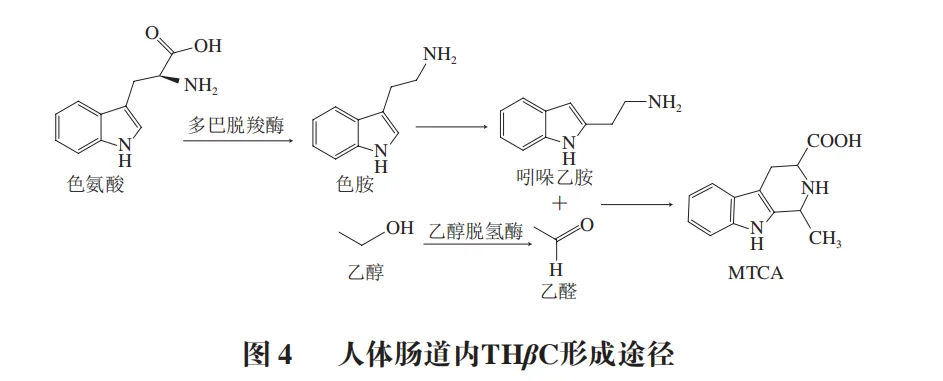
人体中THβC的生物合成涉及色氨酸代谢通路。当摄入高蛋白食物后,肠道菌群通过多巴脱羧酶的脱羧作用将色氨酸转化为色胺。若同时摄入乙醇,乙醇在肝脏经乙醇脱氢酶代谢为乙醛。在胃酸环境(pH 1.5~3.5)下,吲哚乙胺与乙醛通过Pictet-Spengler反应缩合,生成MTCA(图4)。研究表明,乙醇摄入导致MTCA与THCA的比率升高,乙醇被认为可以增强MTCA生成或抑制THCA生成,乙醇造成的影响在摄入后4 h内仍持续。
1.3 影响形成的因素
1.3.1 反应物浓度
THβC形成与醛类和氨基酸或胺等前体物质密切相关。在适宜反应条件下,前体越多,THβC生成越多。在L-色氨酸与酚醛反应生成酚类THβC的模型体系研究中,L-色氨酸、苯甲醛、酚醛浓度的增加,THβC显著增加。高的前体浓度使得反应体系中反应物分子间的碰撞几率增加,从而促进了反应的进行,利于THβC的形成。在葡萄汁制作研究过程中,发酵初期色氨酸迅速被酵母吸收利用导致其浓度快速下降,从而抑制前期THβC的形成,但是随着发酵的持续进行,酵母在生长和代谢过程中产生乙醛,与酒液中剩余的色氨酸反应,导致THβC的含量显著增加。研究人员发现,在Chardonnay葡萄酒发酵过程中,酵母产生的乙醛与发酵后期剩余的色氨酸反应生成了含量较高的MTCA,且随着葡萄酒中色氨酸含量的不同,酒液中MTCA量也不同。
1.3.2 加工温度
加工温度显著影响THβC的生成。Herraiz研究发现,在鱼肉加工过程中,随着加工温度的升高,THCA和MTCA的含量显著增加。这表明在食品加工过程中,高温环境为THβC的生成提供了有利条件。在烘焙咖啡豆时,随着咖啡豆温度的急速提高,THβC含量显著增多。研究发现,在含有色氨酸和乙醛的模型溶液中,温度升高会加快反应速率,促进MTCA的生成。当模型溶液贮存温度为42 ℃和80 ℃时,MTCA的生成量最高;而在-2 ℃和12 ℃时,生成量极少。在酒类贮藏过程中,较高的贮存温度会加速酒中色氨酸和乙醛反应形成THβC,而较低的贮存温度会显著降低反应速率,减少THβC形成。
1.3.3 加工时间
时间是影响THβC形成的关键因素。在食品加工过程中,时间与温度、pH值相互作用,共同影响THβC的形成和累积。在果汁加工过程中,随加工时间延长,L-色氨酸和醛类前体物质反应时间延长,生成的THβC更多。葡萄汁在加热浓缩过程中,随着加工时间的延长,1-戊羟基戊基-1,2,3,4-四氢-β-咔啉-3-羧酸的含量显著上升。在鱼肉加工中也出现同样现象,对鱼肉进行不同时间的烹饪实验,发现随着烹饪时间的延长,THCA和MTCA的含量明显增加。在香蕉和梨贮存过程也发现,随着贮存时间的延长,MTCA的含量显著增加。
1.3.4 反应pH值
pH值对THβC的形成具有显著影响,总体而言酸性条件利于其生成。L-色氨酸与苯甲醛、香草醛等酚醛在酸性水溶液中反应生成酚类THβC,除苯甲醛外的其他酚醛与L-色氨酸在低pH条件下环化反应更容易,THβC形成更多。在水果汁加工和贮存过程中,pH值的改变会影响THβC的含量。在酸性较强的果汁中,THβC的形成更为有利,从而导致其含量相对较高。在模型体系中,pH值对色氨酸与乙醛反应生成MTCA的反应速率影响显著。在25 ℃条件下,pH值低于3时,反应速率极快;pH值在4及以上时,反应速率很慢;当pH值高于6时,在20 d内几乎检测不到MTCA的生成。以达到10 mg/L MTCA所需时间为例,模型pH值为5时约需820 d,pH值为4时约需227 d,pH值为3时约需24 d,pH值为2时约需6.4 d,pH值为1.5时约需5.7 d。这充分说明低pH值能大幅提升反应速率,即pH值越低,MTCA形成越快。
1.3.5 交互作用
食品加工过程中THβC形成是复杂反应介质条件控制的结果。以1-戊羟基戊基-1,2,3,4-四氢-β-咔啉-3-羧酸为例,它由L-色氨酸和D-葡萄糖通过Pictet-Spengler反应形成,在低pH值和高温条件下,其形成量显著增加;然而在高pH值(7和9)时,即使高温也几乎不发生反应;在低温和低pH值条件下,其形成反应速率非常缓慢。相比其他一些THβC,1-戊羟基戊基-1,2,3,4-四氢-β-咔啉-3-羧酸的形成需要更极端的条件。加工时间对THβC形成的影响受到pH值的制约。在L-色氨酸与酚醛反应生成酚类THβC的过程中,在低pH值环境下,加工时间的延长会促进反应进行,生成更多产物。但在高pH条件下,即使加工时间很长,反应也难以有效进行,THβC的形成量很少。
1.3.6 加工方式
不同加工方式会显著影响食品加工过程中THβC的含量。发酵和热加工是食品加工过程中THβC最主要的来源方式。研究人员在巧克力及可可豆的研究中发现,成品中的THβC主要来自发酵。同样,在大蒜的研究中也发现,发酵是发酵大蒜产生THβC的主要方式,且时间对其影响较大。同样有研究表明,对于煮、炸和烟熏等高温加工方式,对形成THβC含量从大到小的热加工方式依次排序为烟熏、炸、煮。烟熏时产生的烟雾中含有醛类等物质,增加了前体物质的浓度,高温也加速了反应进程,导致THβC的生成量上升,烟熏产品表面的THβC含量多于产品内部。烹饪过程中的高温促使前体物质发生反应,随着烹饪时间的延长,反应进行得更充分,从而生成更多的THβC。炸制比煮制过程中加工温度更高,因此会产生更多的THβC。
1.3.7 抑制剂
在食品加工过程中添加抑制剂会减少THβC的生成。目前对于这方面的研究较少。在葡萄酒加工中,二氧化硫用作酿酒中的防腐剂和抗氧化剂,可减少有害物的形成。亚硫酸氢盐阴离子与醛反应生成羟基磺酸盐,可减少醛与发酵液中残留的色氨酸发生反应,从而可可最大限度地减少贮存过程中THβC的形成。同时,色氨酸在亚硫酸盐存在下发生氧化反映被破坏,也会导致发酵液中THβC含量的减少。
2


TTHHβC生物活性

2.1 有益活性
2.1.1 信号传递
单胺氧化酶(MAO)是一种参与神经递质代谢的关键酶,可降解多巴胺、血清素和去甲肾上腺素等单胺类神经递质。相关研究表明,多巴胺经MAO-B降解产生羟自由基,羟自由基作用于神经元后发生氧化应激导致神经元死亡,加速帕金森病发作。Yildiz等研究发现,血清素水平降低与抑郁症发生有关,且血清素水平升高可起到产后抗抑郁作用,尤其是在产后早期。去甲肾上腺素则在调节注意力表现和控制冲动方面起着重要作用,去甲肾上腺素可以调节对相关刺激的注意力焦点并减少分心。THβC及其衍生物可以作为MAO的底物或抑制剂,减少神经递质的降解,从而使这些神经递质在突触间隙中的浓度升高,增强神经信号传递。
2.1.2 抗氧化性
THβC具有显著的抗氧化活性。Ichikawa等从大蒜中分离出4 种THβC,在体外实验中研究中发现,在250 μmol/L的浓度下,(1R,3S)-MTCA、(1S,3S)-MTCA、1-甲基-1,2,3,4-四氢-β-咔啉-1,3-二羧酸(1R,3S)((1R,3S)-MTCdiC)和(1S,3S)-MTCdiC对过氧化氢有较强清除率,其中(1S,3S)-MTCdiC的活性比常见抗氧化剂抗坏血酸更强。(1S,3S)-MTCdiC可通过在1位点脱羧、在1到2位之间脱水提供电子清除过氧化氢,起到抗氧化作用(图5)。
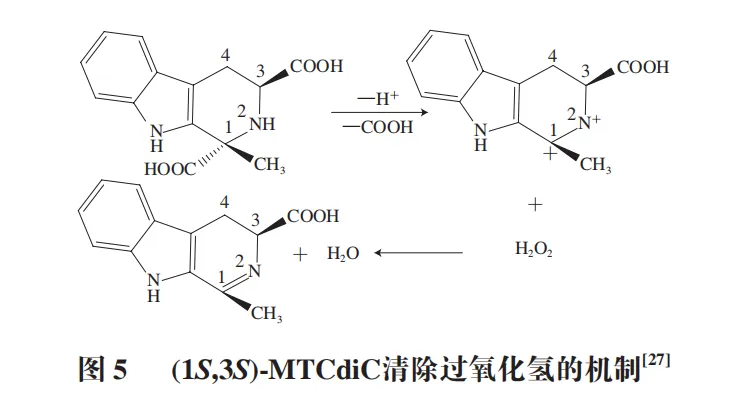
与此同时,Sato等发现短期发酵大蒜提取物中(1R,3S)-MTCA和(1S,3S)-MTCA含量增加,且发酵大蒜提取物对过氧化氢的清除活性比未发酵的大蒜提取物至少强10 倍,这表明THβC水平的增加与活性氧清除活性的增强相关。Yang Li等从车前玉簪花中提取到(1R,3S)-MTCA和(1S,3S)-MTCA,通过研究发现这两种THβC都具有较强的抗氧化活性。Arutselvan等通过色氨酸与乙醛的Pictet-Spengler缩合反应合成了MTCA异构体,并通过硫酸铁法评价了该化合物在亚油酸自氧化系统中的抗氧化活性,结果表明该化合物具有中等强度的抗氧化活性。此外,在亚油酸体系中,MTCA和MTCdiC还能抑制2,2′-偶氮二(2-甲基丙基咪)二盐酸盐诱导的脂质过氧化。
2.1.3 抗菌抗炎
THβC的刚性吲哚环和柔性哌啶基序使其具有一系列显著的药理学特性。Liang Yan等研究发现一系列苯甲酰的THβC衍生物对奶牛乳腺炎的病原菌具有较弱的抑菌作用。Song Yuanze等合成了一系列1,2,3,4-THβC衍生物,并评估了它们对革兰氏阳性菌和革兰氏阴性菌的抗菌能力。结果表明,这些化合物均表现出中度至良好的抗菌活性,其中4-苯并氨基苯基-THβC对革兰氏阳性菌,尤其是耐甲氧西林金黄色葡萄球菌的抑制作用优于庆大霉素。部分THβC表现出一定的抗炎作用,可用于抗炎药物的研发。1-乙基-1,2,3,4-四氢-β-咔啉-3-羧酸和1-苯乙酮-1,2,3,4-四氢-β-咔啉-3-羧酸通过抑制RAW264.7巨噬细胞中的炎症介质一氧化氮的生成发挥其抗炎活性,这两种THβC衍生物能抑制NO的过量生成,且具有剂量依赖性。
2.2 不良活性
2.2.1 诱导杂环胺形成
杂环胺被国际癌症研究机构归类为2A类致癌物。实验表明杂环胺能使小鼠和大鼠等动物多个器官产生肿瘤,在非人类灵长类动物中也有引发肿瘤的案例。对于人体而言,虽然直接致癌证据有限,但大量流行病学研究表明,长期大量食用含杂环胺的食物,会增加患结肠癌、乳腺癌和前列腺癌等癌症的风险。杂环胺是易致突变的化合物之一,其代谢中的氮离子中间体是强大的亲电试剂,易于与DNA形成C-8鸟嘌呤加合物,导致细胞发生突变。在食品加工中,食物中的色氨酸容易与葡萄糖发生Pictet-Spengler反应生成THβC,进一步氧化为Harman和Norharman等β-咔啉类杂环胺,反应步骤见图6。由此可知,THβC是杂环胺的前体物质,减少食品加工中ThβC的含量,可减少食品中杂环胺的形成风险。
2.2.2 诱导N-亚硝基化合物形成
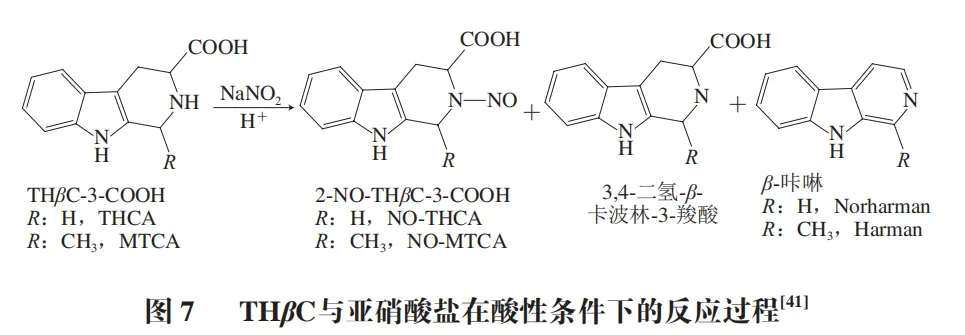
THβC可与食物、口腔和胃中的亚硝酸盐发生反应,生成强致癌化合物N-亚硝基化合物。同时一些THβC也是致突变化合物。Wakabayashi等从日本酱油中提取出两种THβC,鉴定后确定其为诱变剂亚硝基化合物的前体物质。THβCs与亚硝酸盐可在酸性条件下反应,人体胃部pH值在1~2之间,为这类化合物的亚硝化提供了适宜环境(图7)。在模拟胃液条件下,N-亚硝基THβCs会发生降解并产生亚硝化衍生物,亚硝化衍生物可与DNA中的碱基反应,导致DNA损伤和突变。例如,亚硝化反应可能使鸟嘌呤转化为黄嘌呤,引起DNA的G∶C到A∶T转换,这种突变在某些致癌过程中起重要作用,增加患癌风险。
3


食品中TTHHβC检测技术

3.1 前处理
通常使用离子交换或反相机制等技术将THβC从复杂的食物和生物基质中分离出来,主要包括液-液萃取和固相萃取(SPE)等技术。作为水性缩合产物,THβC在样品制备过程中可由痕量游离醛和吲哚胺反应形成。为防止样品制备过程中THβC的形成,常用氨基脲作为醛类捕获试剂,或用氟虫胺或氯甲酸甲酯捕获吲哚胺等前体物质。THβC形成与pH值有关,常在中性或微碱性pH值条件下均质可以减少或避免样品制备中THβC的形成。与此同时,尽可能减少样品前处理,将样品直接引入色谱仪,有助于降低前处理过程中THβC的形成。
3.1.1 SPE
SPE的发展促进了从食品和生物样品等复杂基质中分离THβC。食品中萃取THβC主要使用C18柱和SCX柱。
1) C18柱
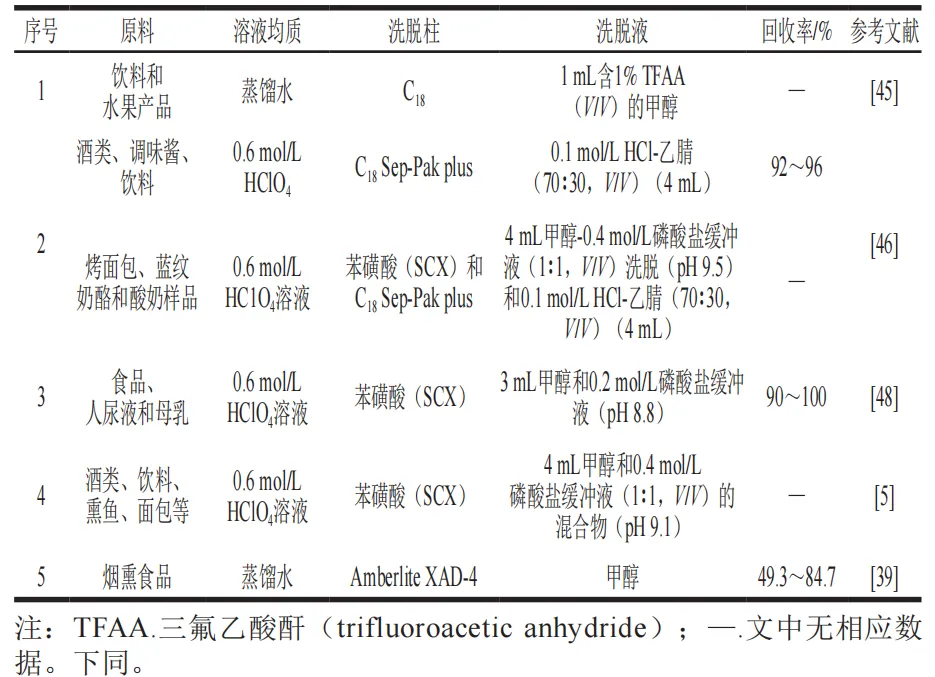
C18柱的分离原理是基于溶质分子与固定相之间的疏水相互作用、极性基团的偶极作用和氢键作用等次级力,从而实现对样品中非极性或中极性化合物的分离、纯化和富集。早在1996年,就已经将SPE(用乙腈洗脱的C18小柱)用于分离大鼠脑中的THβC,该C18柱分离得到的样品纯度低于92%。Gutsche等也用C18小柱分离乙醇饮料和水果产品中的THβC,在此程序中,将用蒸馏水稀释的等分试样调节至pH值为1,并加载到C18小柱上。用酸性水洗涤小柱,用含有1%(V/V)三氟乙酸(TFA)的甲醇洗脱化合物。蒸发后,将重新溶解在乙腈水溶液中的提取物利用高效液相色谱-串联质谱(HPLC-MS/MS)检测。
C18柱对复杂基质的干扰物(如蛋白质和脂质等)去除效果较差,需结合SCX柱联用以提高回收率。Herraiz等采用4 个C18 Sep-Pak plus柱或6 个苯磺酸SCX柱结合4 个C18 Sep-Pak plus柱对不同样品进行分离。对于酒类、调味酱和饮料等非固体样品,采用0.6 mol/L HClO4溶液进行匀浆,匀浆后先用甲醇(2 mL)和0.1 mol/L HCl(2 mL)冲洗柱子。洗脱液为0.1 mol/L HCl(2 mL)、0.1 mol/L HCl-乙腈(90∶10,V/V)(2 mL)和0.1 mol/L HCl-乙腈(70∶30,V/V)(4 mL)的混合溶液。对于烤面包,奶酪等固体样品可采用6 个苯磺酸SCX柱结合4 个C18 Sep-Pak plus柱对THβC进行分离,也可采用4 个C18 Sep-Pak plus柱直接进行分离。
2) SCX柱
目前,在分离THβC时苯磺酸SCX柱应用较多。苯磺酸强阳离子交换柱(SCX柱)的分离原理是通过柱内填充的强阳离子交换树脂与目标阳离子发生静电吸附-解吸作用,实现对样品中阳离子的分离、纯化和富集。Herraiz等使用苯磺酸SCX色谱柱进行SPE分离游离THCA,将含有氨基脲的酸性样品加载到SCX柱上,用0.1 mol/L HCl、甲醇、水洗涤,并用0.4 mol/L磷酸盐缓冲液(pH 9.1)冲洗。THβC用0.4 mol/L磷酸盐缓冲溶液和甲醇(1∶1,V/V)(pH 10.1)的混合溶液回收,该混合溶液可以破坏THβC与吸附剂之间的离子和疏水相互作用。THβC回收率≥95 %,相对标准偏差<6%。
对于人尿液和母乳中的THβC也可采用SCX柱进行分离。先采用3 mL甲醇洗涤两次,3 mL 0.1 mol/L盐酸洗涤两次活化色谱柱,然后用5 mL 0.1 mol/L盐酸溶液、2 mL甲醇和6 mL水进行洗涤,再用3 mL 0.4 mol/L磷酸盐缓冲液(pH 9.1)冲洗每根色谱柱,最后用3 mL甲醇和0.2 mol/L磷酸盐缓冲液(pH 8.8)的混合物洗脱THβC。
与C18-SPE相比,苯磺酸SCX柱更能去除干扰化合物,更多地用于肉类这种杂质多的样品。通过使用苯磺酸SCX柱,然后进行反相(RP)-HPLC-荧光检测器(FLD)检测,能获得THβC的干净色谱图。
3) 其他色谱柱
除了以上常见的用C18-SPE和苯磺酸SCX柱分离以外,也有研究者用Amberlite XAD-4柱分离THβC。样品前处理步骤较为复杂,包括冷冻干燥、匀浆、离心和多次调节pH值等步骤。上样后,采用蒸馏水作为洗涤液,色氨酸处理的无羰基甲醇作为洗脱液,洗脱THβC。表1对多种原料采用的SPE方法进行比较,以供后续研究参考。

3.1.2 液-液萃取
目前使用液-液萃取分离THβC的研究较少。Arbouche等采用乙醚-二氯甲烷-己烷-异戊醇(50∶30∶20∶0.5,V/V)的混合溶液作为萃取剂对头发进行液-液萃取。Mußhoff等将碱化尿液在室温下水解30 min,用顺丁烯二酸(0.1 g)酸化,加入5 mL己烷-乙酸乙酯(9∶1,V/V)混和溶液作为萃取液。Tsuchiya等对乙醇饮料中的THCA进行了分析,用荧光素处理后,在碱性培养基中用乙酸乙酯提取反应混合物,再将混合物加入0.2 mol/L HCl溶液中,然后在碱性培养基中用乙醚提取,提取物蒸发再溶解后用于后续的RP-HPLC检测,在啤酒、葡萄酒和尿液中的回收率高于90%。Lee等用80%甲醇溶液作为萃取剂从猪肉样品中提取出一种THβC。
液-液萃取还可和SPE组合使用对THβC进行必要的分离、浓缩和净化。例如采用SPE(C18和SCX小柱)和液-液萃取,进而鉴定食品中的THβC-3-COOH。在HPLC和气相色谱-质谱(GC-MS)联用技术检测之前,使用二氯甲烷萃取(pH 9~10)和SCX从乙醇饮料中分离出一种新的β-咔啉酯——1-甲基-1,2,3,4-四氢-β-咔啉-3-羧酸乙酯,回收率高于90%。在表2和表3中对4 种液-液萃取方法和多种前处理方法的原料、试剂和回收率进行比较,以供后续实验参考。


3.2 检测技术
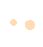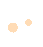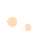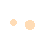
3.2.1 GC-MS
GC部分的分离原理是基于各组分在固定相和流动相(载气)之间的分配系数差异,通过色谱柱时保留时间不同而分离。然后进入质谱仪,通常是电子电离源电离后产生碎片离子,通过质量分析器(比如四极杆)按质荷比分离,检测器检测,得到质谱图。通过保留时间和质谱图定性,峰面积定量。
THβC常温下是非挥发性或弱挥发性物质,采用GC-MS检测时需要进行衍生化处理。THβC衍生化一般用酰化试剂,例如氯甲酸甲酯(MCF)、TFAA、五氟丙酸酐或七氟丁酸酐(HFBA)等。THβC经过氯甲酸甲酯衍生化形成甲酯衍生物便于分析3-羧基化合物。
最常用的衍生化试剂是MCF。Herratz等用MCF对调味酱、酒类、果汁、烤面包、蓝纹奶酪和酸奶等产品进行衍生化处理,按照一定体积样品中THβC含量的不同,采用不同的前处理程序。最后通过GC-MS检测到6 种THβC。Herratz用MCF对酒类样品中的THβC进行衍生化,结合GC-MS和RP-HPLC检测出一种新型THβC。
由于MCF的毒性,也有研究采用其他的衍生化试剂处理样品。较为常用的试剂是HFBA。Bringmann等用HFBA衍生化样品,后通过GC-MS和选择性离子监测模式在大鼠血液和尿液等样品中检测出一种新型氯化THβC——1-三氯甲基-1,2,3,4-四氢-β-咔啉,全血、尿液和脑中的检测限分别为4、2 ng/g和5 ng/g,定量限分别为12、10 ng/mL和15 ng/g。Matsubara等用HFBA将人尿液衍生化处理后,用GC-MS检测出两种新型THβC,分别为1-甲基-l,2,3,4-四氢-β-咔啉和6-甲氧基-1-甲基-1,2,3,4-四氢-β-咔啉。两种化合物的检测限分别为0.2 pmol/mL和2 pmoL/mL,变异系数分别为4.3%和4.9%。然而,食品中THβC检测很少使用GC-MS,其原因在于THβC为非挥发性或弱挥发性物质,在进行GC-MS前需要进行衍生化处理,与其他的检测方法相比,操作更复杂。
3.2.2 HPLC-FLD
HPLC与FLD或MS检测结合分析THβC,简单、选择性好且灵敏度好。HPLC-FLD的检测原理为:当经HPLC分离后的THβC流经FLD时,THβC被特定波长的激发光(λem,一般为270 nm,对应吲哚环共轭结构)照射,分子从基态跃迁到激发态,返回基态时发射更长波长的荧光(λem,一般为343 nm),检测器仅接收特定λem的信号,能显著降低背景干扰(如溶剂、杂质的非特异性吸收)。THβC分子结构中含吲哚环,具有共轭双键,通常具备天然荧光特性,可直接用FLD检测,无需进行衍生化处理。Herraiz用HPLC-FLD从酒类、饮料、调味酱、面包和熏鱼等样品中检测出3 种THβC,分别为THCA、(1S,2S)-MTCA、(1R,2S)-MTCA,但具体的检测限、定量限并未提及。与此同时,Herraiz也通过RPHPLC-FLD检测商业食品中的THβC。通过将THβC衍生化,得到THβC的N-甲氧基羰基衍生物,可保持荧光特性,并且在相同的色谱条件下比母体化合物保留的时间更长。
3.2.3 HPLC-MS/MS
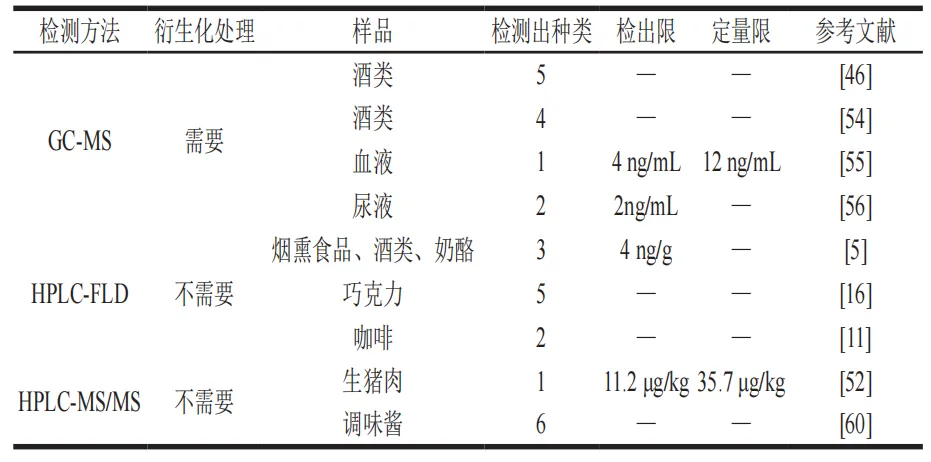
HPLC分离不同组分,MS则对每个组分进行离子化和质量分析,通过保留时间和质荷比定性,峰面积或峰高定量。Gutsche等开发出一种不需要进行样品前处理即可直接进行检测的HPLC-电喷雾电离(ESI)-MS/MS方法。其将调味酱进行稀释过滤后直接进行检测,采用Symmetry-C18色谱柱,通过比较保留时间、子母离子信息([M+H]+),鉴定出6 种THβC,检测限为10 ng/mL。Lee等为检测肉类样品中L-色氨酸衍生物的残留,开发出一种HPLC-MS/MS的检测方法。先将样品用蒸馏水均质,后加入80%甲醇溶液(10 mL),用Cortecs C8柱进行分离,流动相A由10 mmol/L甲酸铵和0.1%甲酸水溶液组成,流动相B由含0.1%甲酸的甲醇溶液组成,进行梯度洗脱,检测器为配备有ESI离子源的TSQAltis三重四极杆质谱仪,最终检测到一种THβC,为MTCA,检出限和定量限分别低于11.2 μg/kg和35.7 μg/kg(表4)。
4


结 语

人体内THβC主要来源于膳食摄入,THβC具有双重性生物活性,一方面,THβC衍生物可以抑制MAO的活性,增强神经信号传递,还具有抗菌抗炎抗氧化等作用,可以用于相关药物的研发;另一方面,THβC氧化产物Harman和Norharman会增强其他杂环胺的致癌性,THβC的N-亚硝基衍生物会作用于DNA,导致DNA受损和断裂,其健康风险不容忽视。因此,建立一个符合健康要求的每日摄入安全限量至关重要。食品高温加工过程中易产生THβC,部分影响THβC形成的因素已明确,如温度、pH值、时间和前体物质的量等,但反应条件具体的作用机制尚不明确,不同反应条件之间的交互作用研究较少,THβC对食品(特别是肉制品)品质的影响尚不清楚,需要进一步研究其形成机制和影响条件,从而为控制食品加工中THβC的形成提供理论依据。

第一作者:

曹佳敏,硕士研究生在读,研究方向为肉品加工与质量安全控制。
教育工作:
2020.09-2024.06,鲁东大学食品科学与工程学院,学士;
2024.09-至今,四川农业大学食品学院,硕士(在读)。
通信作者:

惠腾,博士(后),副教授,硕士生导师,四川省“天府峨眉计划”青年人才。一直从事肉品科学与加工技术研究,聚焦肉品加工与质量安全控制、生鲜肉保鲜减损、屠宰副产物高值化利用与装备研发。
教育工作经历:
2022.05-至今,四川农业大学食品学院,副教授;
2018.12-2022.04,中国农业科学院农产品加工研究所,助理研究员;
2016.11-2018.10,中国农业大学食品科学与营养工程学院,博士后;
2011.09-2016.09,南京农业大学食品科技学院,博士;
2007.09-2011.06,南京农业大学食品科技学院,学士。
主持科研项目:
1. 国家自然科学基金面上项目,内外源因子关联调控Pictet-Spengler反应诱导烤肉中9H-吡啶并[3,4-b]吲哚形成机理,2024-2027。
2. 国家自然科学基金青年基金,活性羰基物质诱导肉制品中2-氨基-1-甲基-6-苯基咪唑并[4,5-b]吡啶(PhIP)形成机理,2019-2022。
3. 中国博士后科学基金面上项目,活性羰基物质诱导肉制品加工过程中9H-吡啶并[3,4-b]吲哚形成机理,2022-2024。
4.“十四五”国家重点研发计划子课题,易腐农产品跨域运输多级仓储网络布局与优化研究,2024-2027。
5.“十三五”国家重点研发计划子课题,传统熏炸烤食品添加剂减量增效与品质提升协同技术研发,2019-2022。
社会兼职:
1. 中国农学会农产品贮藏加工分会第五届委员会委员。
2. 中国畜产品加工研究会第八届理事会理事。


引文格式:
曹佳敏, 李雪辰, 许嘉豪, 等. 热加工食品中四氢-β-咔啉的形成及其检测方法[J]. 食品科学, 2026, 47(1): 327-335. DOI:10.7506/spkx1002-6630-20250604-016.
CAO Jiamin, LI Xuechen, XU Jiahao, et al. Formation of tetrahydro-β-carbolines in thermally processed foods and their detection methods[J]. Food Science, 2026, 47(1): 327-335. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-20250604-016.
点击下方阅读原文即可查看文章相关信息。


实习编辑:李雄;责任编辑:张睿梅。点击下方阅读原文即可查看全文。图片来源于文章原文及摄图网
近期研究热点
《食品科学》:浙江省农业科学院杨颖副研究员等:两种浙产铁皮石斛茎多糖对秀丽隐杆线虫的抗衰老作用
《食品科学》:山东省食品药品检验研究院刘艳明研究员等:室温皂化联合气相色谱-四极杆-飞行时间质谱检测肉制品中的胆固醇氧化物
《食品科学》:沈阳农业大学周倩教授等:外源花青素处理调控采后蓝莓果实抗氧化能力保持果实品质的机理
《食品科学》:中国农业大学杨海霞教授等:乌鸡蛋黄油组分分析及其缓解β-乳球蛋白致敏小鼠过敏反应的作用机制
《食品科学》:塔里木大学张锐利教授、中国农业科学院毕金峰研究员等:多维度解析烫漂预处理对吊干杏果干质构形成的影响机理


长按或微信扫码进行注册
会议招商招展
联系人:杨红;电话:010-83152138;手机:13522179918(微信同号)

随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 管控漏洞频发!农行河南两家机构违规支付房产中介佣金遭监管重罚 多名责任人被追责
- 青·城纪丨她带着一群“农业小白”,在山里把日子过成“向往的生活”
- PBJ | 中国农业大学孙其信院士团队揭示小穗数基因WAPO1通过翻译后修饰TaFT1调控小麦穗型与产量分子机制
- 农业农村部四部门联合推介奶牛健康监测与疾病早期诊断技术!
- 青海省惠民惠农财政补贴政策项目之农业生产救灾资金-饲草料调运补助项目
- 【农业类】低版面费知网期刊大合集!最好发的农业知网刊都在这里了!选对期刊,录用轻松拿下!
- 作品征集丨2026福州渔业工美创作大赛
- 关于“生态农业优质高产'四位一体'种植技术”的深度调研报告
- 农业农村部公示2026年第一批全国名特优新农产品及特质农品;现代产业技术体系首席科学家和岗位科学家名单公示|一周快讯
- 南京农业大学成立农业合成生物学中心
