研究背景
玉米作为重要的粮食、饲料和工业原料,是全球种植面积最广的农作物之一。在全球气候持续变暖、极端天气频发、粮食需求不断上升和机械化收获需求增加的背景下,对“早熟、高产”玉米品种的需求也越来越迫切。然而,早花等位基因往往具有多效性,在促进开花的同时,常对产量产生不利影响。这一现象长期困扰着育种家,是作物育种中亟待突破的科学难题。本研究以玉米开花抑制因子ZmRap2.7为切入点,解析了其调控开花期和产量性状的多效性机制,并通过编辑ZmRap2.7的调控区实现了两个性状的解耦联。
文章介绍
近日,Plant Biotechnology Journal 杂志在线发表了由中国农业大学植物抗逆高效全国重点实验室、国家玉米改良中心田丰教授团队撰写的题为“Designed alleles of ZmRap2.7 decouple the trade-off between early flowering and yield penalty”的研究论文。该研究系统解析了玉米开花抑制因子ZmRap2.7介导的开花期和产量性状多效性的分子机制,并通过CRISPR/Cas9技术对ZmRap2.7启动子及其上游增强子Vgt1进行编辑,成功创制了ZmRap2.7在茎尖分生组织(SAM)中特异性下调的编辑系,成功打破早熟与减产之间的负向耦联,为解决作物改良中由基因多效性导致的性状权衡问题提供了新思路和有效策略。
文章链接:https://onlinelibrary.wiley.com/doi/10.1111/pbi.70679
文章信息:

主要内容
ZmRap2.7 最初被鉴定为玉米开花抑制因子,该基因上游约 70 千碱基处的微型反向重复转座元件插入可调控其表达。后续研究发现,ZmRap2.7 还能够调控气生根数量、种子活力等性状,这与该基因在多种组织中广泛表达的特征相符。为深入探究 ZmRap2.7 的多效性功能,本研究创制了 ZmRap2.7-KO1 与 ZmRap2.7-KO2 两个基因敲除株系。与前人研究结果一致,无论在北京长日照环境还是三亚短日照环境下,敲除株系的开花时间均显著早于野生型植株。同时,基因敲除会改变玉米株型,表现为株高与穗位高度降低、叶夹角增大、叶片数量减少;相较于野生型,敲除株系果穗穗长与穗粗减小、果穗整体偏小,籽粒体积与粒重也随之下降,最终造成单株籽粒产量显著降低。综上,ZmRap2.7 可抑制植株开花进程,同时正向调控产量相关性状;该基因在叶片、茎尖分生组织、根系、发育中的花序及籽粒等组织中广泛表达,为其调控植株生长发育与产量形成的多效作用奠定了分子基础(图1)。
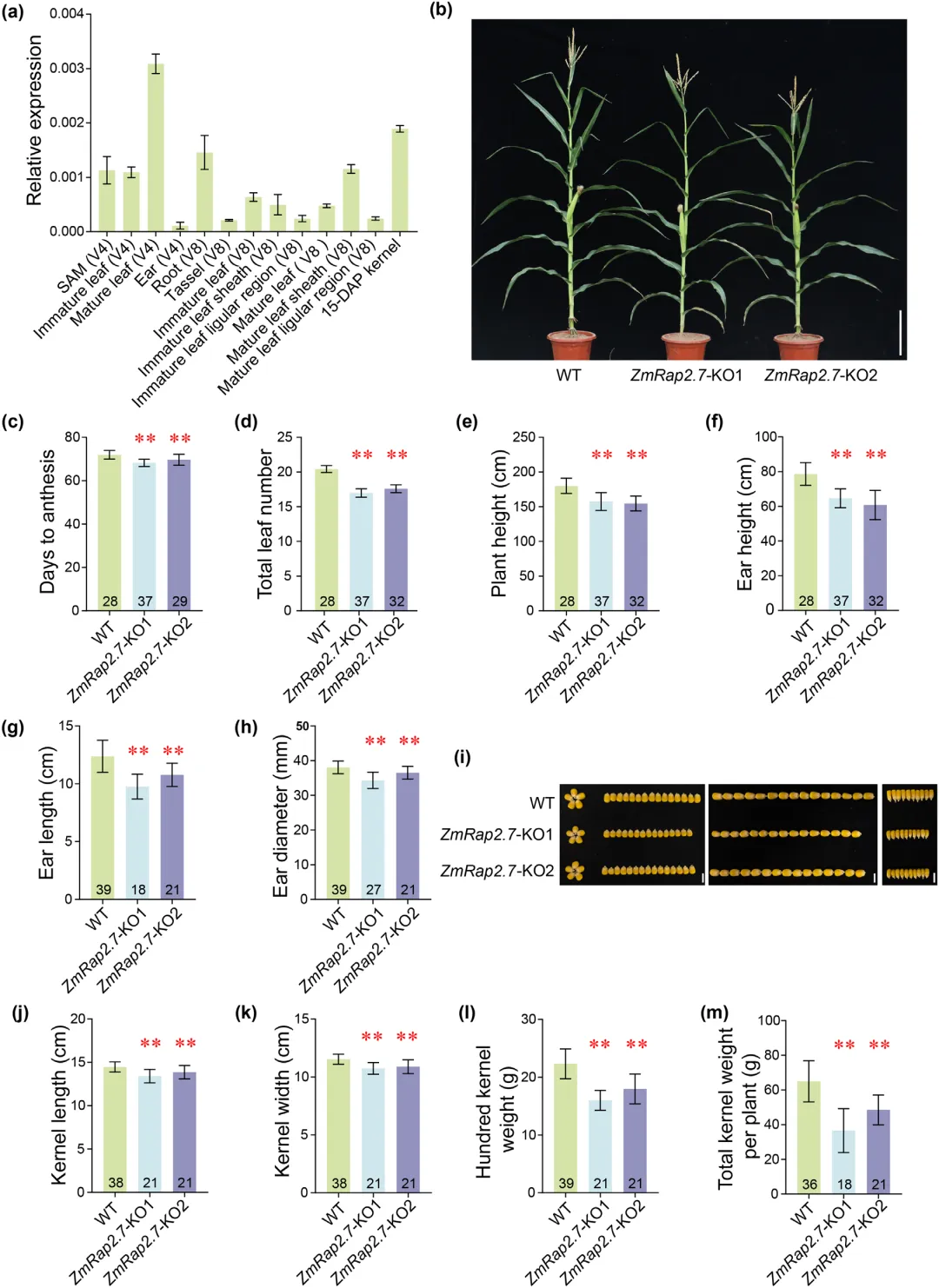
图1 ZmRap2.7在玉米中的多效性表型鉴定
为解析 ZmRap2.7 调控开花的分子机制,该研究在成花转换期采集 ZmRap2.7 敲除株系与野生型成熟叶片开展转录组测序,共筛选得到 311 个差异表达基因,其中包含 ZCN8、ZmMADS67 等多个已报道的开花相关基因。研究发现,玉米成花素基因 ZCN8 在敲除株系中表达显著上调,与已有研究结论相符。为明确 ZmRap2.7 与 ZCN8 的遗传关联,将两种基因敲除材料杂交获得双突变体,表型观测显示双突变体开花时间与 ZCN8 单突变体基本一致,说明 ZmRap2.7 敲除引发的早花表型主要依赖 ZCN8 发挥功能,据此推测 ZCN8 是 ZmRap2.7 的直接作用靶点。依据已知结合基序,在 ZCN8 起始密码子上游约 3.3 千碱基区域鉴定出两处潜在结合位点,酵母单杂交、凝胶迁移阻滞实验与染色质免疫共沉淀定量 PCR 试验均证实二者可直接结合,表明 ZmRap2.7 能够通过直接抑制叶片中 ZCN8 介导的成花素信号,进而负向调控植株开花进程。双突变体开花时间仍略早于 ZCN8 单突变体,提示存在其他下游基因参与开花调控。鉴于 ZmRap2.7 在成花关键部位茎尖分生组织中表达,推测该基因还可通过茎尖分生组织通路调控开花。对该组织样本进行转录组分析,鉴定出 456 个差异表达基因,其中涵盖多个经典开花调控基因,实时荧光定量 PCR 验证了基因表达差异的真实性。凝胶迁移与酵母单杂交实验证明,ZmRap2.7 可直接结合 ZmCOL3 与 ZmSPL29 的启动子并调控其表达。差异基因功能富集显著富集于生殖生长与花发育通路,证实 ZmRap2.7 是茎尖分生组织成花转换过程中的核心调控因子(图2)。
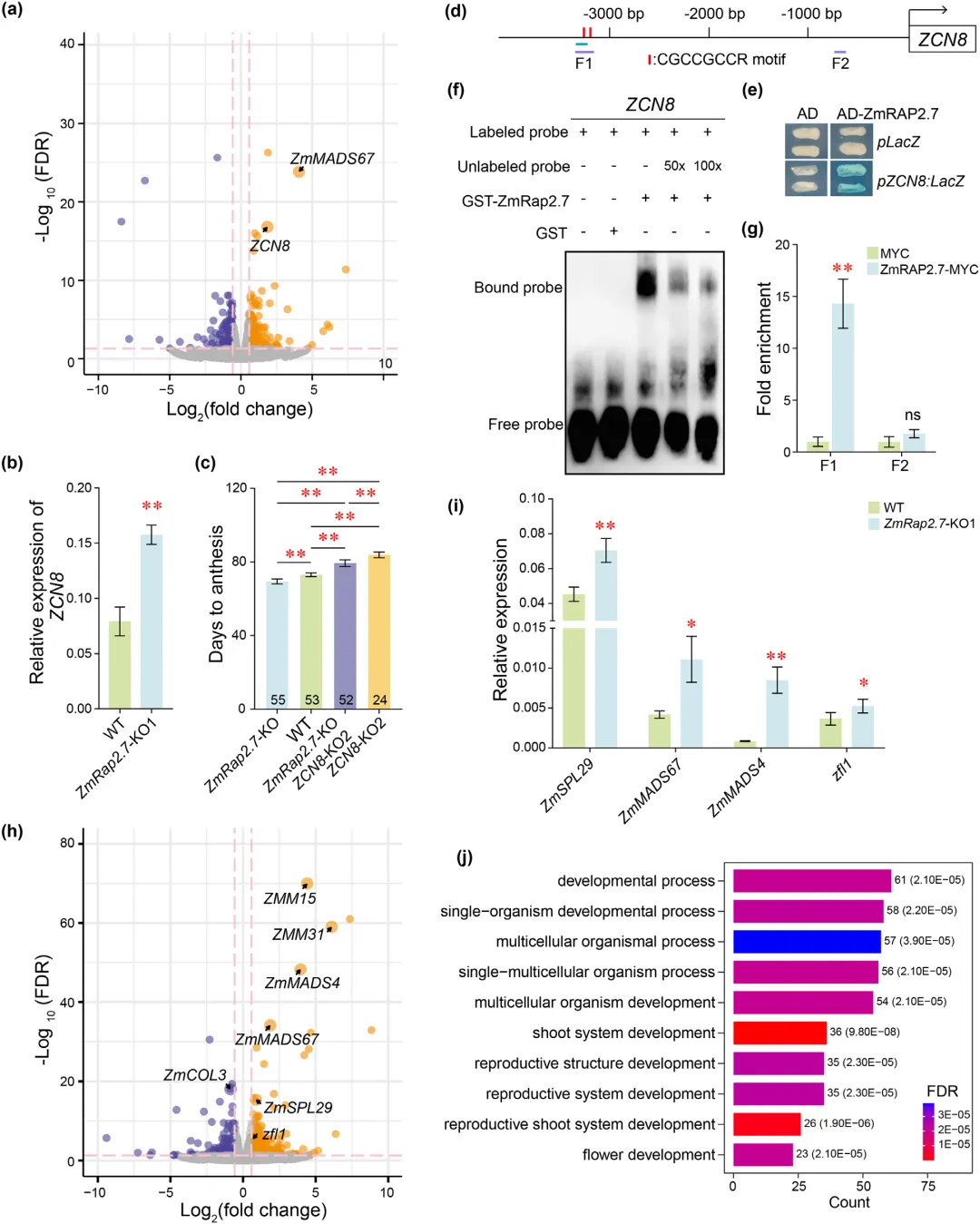
图2 ZmRap2.7在叶片和SAM中延迟开花的调控机制
生殖生长阶段,玉米雌穗花序分生组织依次分化形成小穗对分生组织、小穗分生组织与花分生组织,花序分生组织体积可决定花分生组织形成能力,进而影响穗长与穗粗。电镜观测发现,ZmRap2.7 敲除株系花序分生组织显著短小,原位杂交结果显示该基因主要在各类穗部分生组织中表达,具备维持分生组织活性的作用。对幼穗开展转录组测序共鉴定出 3784 个差异表达基因,功能富集集中于信号传导、激素调控、生长及生殖结构发育等过程,生长素、乙烯相关通路基因差异显著。敲除株系中多条生长素通路基因表达下调,说明 ZmRap2.7 可通过生长素信号调控穗部发育;多个调控花发育的 MADS-box 家族基因表达也发生明显改变,其中花序特征基因 ZmMADS4 表达显著上调。田间表型试验证实 ZmMADS4 负调控雌穗大小,其过表达株系穗型缩小,敲除株系穗部增大。序列分析与酵母单杂交、凝胶迁移、染色质免疫共沉淀试验验证,ZmRap2.7 可直接结合 ZmMADS4 启动子区域抑制其表达。双突变体表型分析进一步证明,ZmRap2.7 通过靶向抑制 ZmMADS4,进而调控玉米雌穗发育与穗部形态(图3)。
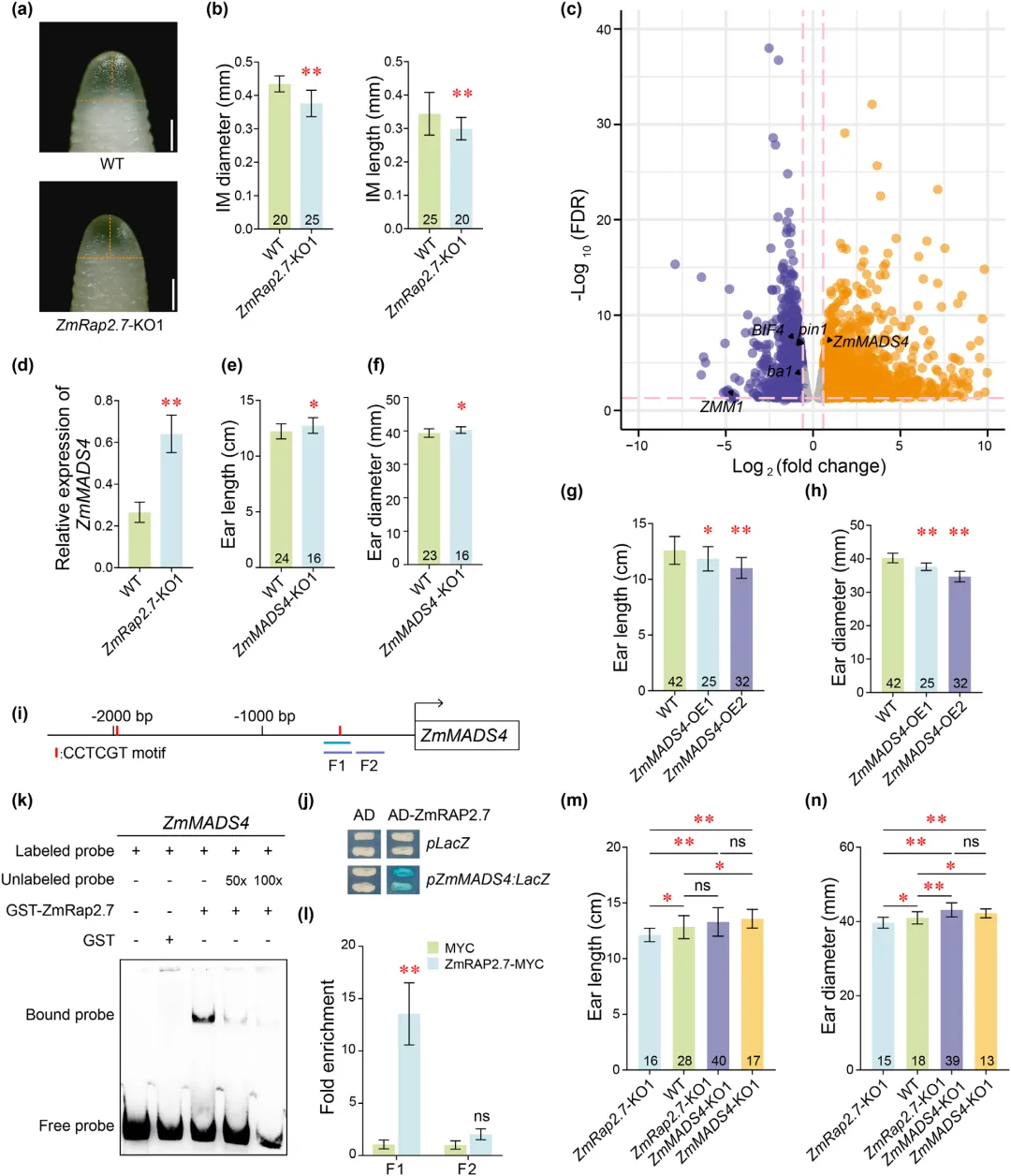
图3 ZmRap2.7通过抑制ZmMADS4调控玉米穗发育的机制
胚乳是玉米籽粒主要组成部分,决定籽粒大小与重量,淀粉和蛋白质为胚乳主要储藏物质。测定结果显示,ZmRap2.7 敲除株系与野生型籽粒的淀粉、蛋白质及可溶性糖含量无显著差异。籽粒大小还受细胞扩张影响,授粉后 15 天籽粒胚乳细胞数量趋于稳定,检测发现敲除株系胚乳细胞宽度明显减小,细胞总数无明显变化,说明细胞体积差异是籽粒大小和粒重改变的主要原因。
对该时期籽粒开展转录组测序,共筛选出 2457 个差异表达基因,其中多数五肽重复序列家族基因表达下调,这类基因已被证实参与籽粒发育。胚乳特异性基因 ZmGRAS11 在敲除株系中表达显著降低,该基因缺失会缩小籽粒尺寸且不影响储藏物质积累,二者籽粒缺陷表型高度相似,且基因表达模式相近,推测 ZmGRAS11 作用于 ZmRap2.7 下游。分子互作试验证实 ZmRap2.7 可直接结合 ZmGRAS11 启动子区域,表明 ZmRap2.7 通过正向调控 ZmGRAS11 表达,进而增大玉米籽粒体积、提升粒重(图4)。
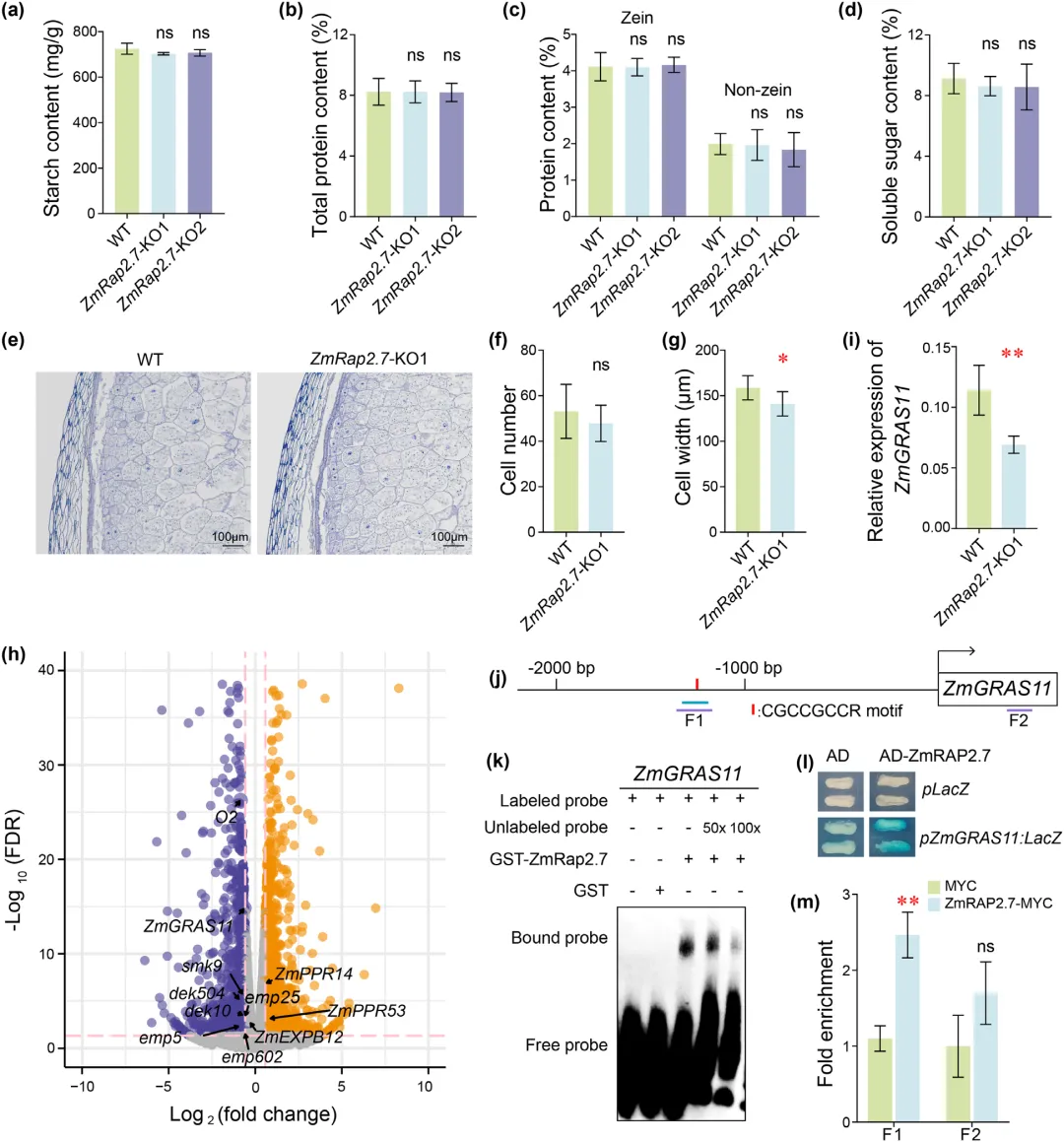
图4 ZmRap2.7通过激活ZmGRAS11调控玉米籽粒发育的机制
ZmRap2.7 的多效性源于其多组织广泛表达特性。对比叶片、茎尖分生组织、幼穗与籽粒的差异基因,筛选出 266 个共有差异基因,半数以上表达变化趋势一致,雌雄穗早期发育通路重叠致使开花期与产量性状易形成权衡制约。参考启动子编辑定向调控基因表达的成功案例,推测编辑 ZmRap2.7 调控区可缓解早花带来的产量损失。该基因上游的 Vgt1 区域为增强子,可通过染色质环调控基因表达,天然微型转座子插入会减弱增强子活性、下调基因表达并促使植株早花。
利用 CRISPR-Cas9 技术编辑玉米自交系 LH244 的 Vgt1 增强子与基因启动子,获得多类纯合编辑株系。田间鉴定表明,部分编辑株系开花时间显著提前,早花效应优势突出,且单株产量未出现明显下降,具备早熟育种应用潜力。
表达检测发现,编辑株系仅茎尖分生组织中 ZmRap2.7 表达显著下调,叶片、幼穗及籽粒中表达基本稳定,下游开花相关基因表达随之发生组织特异性改变。双荧光素酶试验证实编辑位点序列变异降低了调控元件活性。综上,顺式调控编辑可实现 ZmRap2.7 组织特异性表达抑制,在促进开花的同时维持穗部与籽粒正常发育,能够有效打破开花期与产量间的性状权衡,为作物多效基因的性状改良提供可行技术路径(图5)。
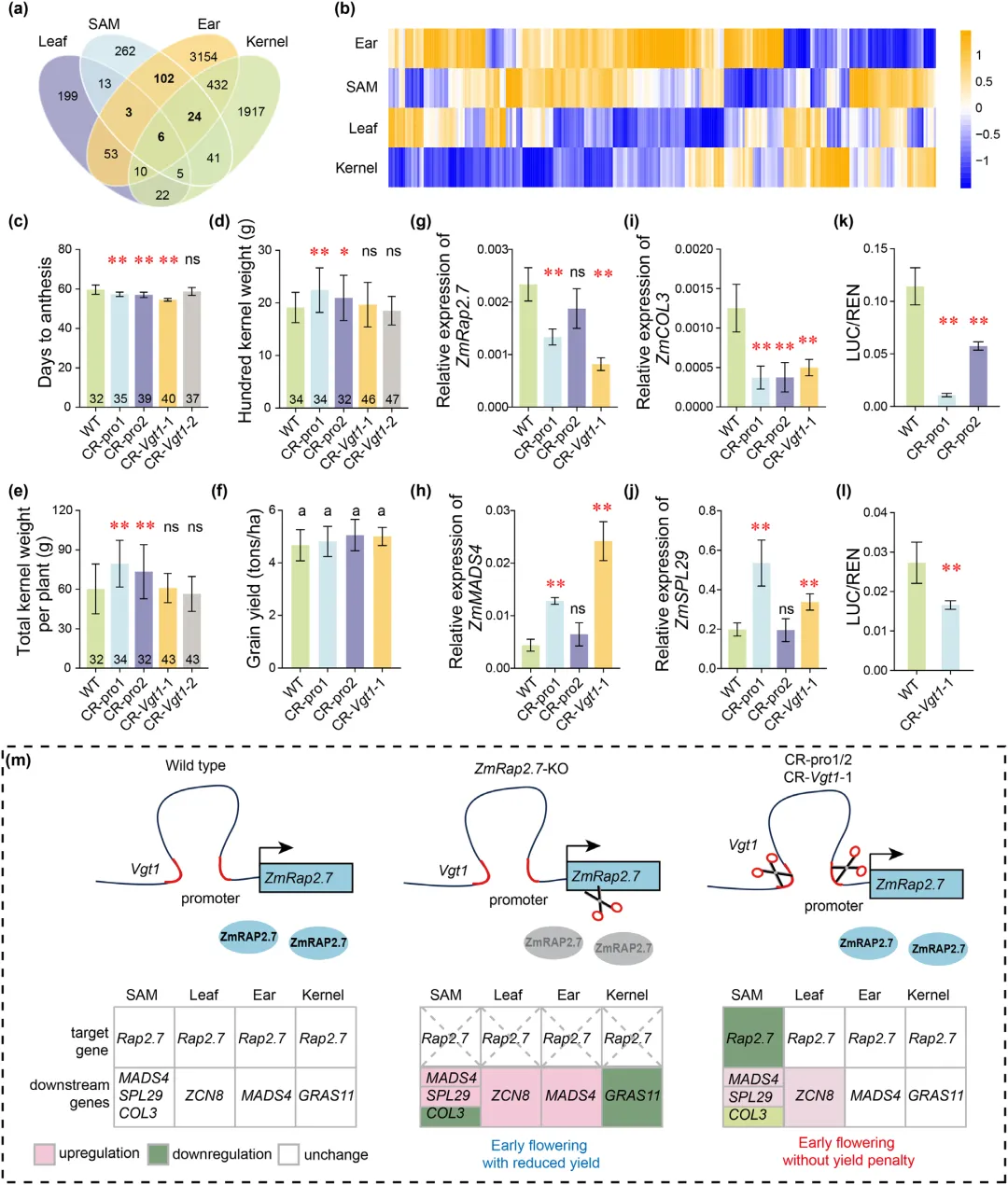
图5 ZmRap2.7编辑株系解耦早花与产量损失的表型与机制
研究结论
该研究系统解析了单一基因同时调控多个农艺性状的多效性分子机制,首次通过顺式调控区编辑实现了玉米“早花不减产”的性状改良,为玉米早熟高产品种培育提供了珍贵的新种质资源。这种通过组织特异性顺式调控编辑解耦多效性性状的策略,也为水稻、小麦等其他作物的复杂性状精准改良提供了可推广的范式。未来可进一步评估这些编辑等位基因在不同遗传背景和生态区的产量稳定性,同时深入解析调控区编辑实现组织特异性表达的精细分子机制,拓展该策略在更多功能基因上的应用,为作物适应性育种提供更多创新路径。
END
本期分享员
刘阳
2025级硕士研究生


