草莓花序构型直接决定果实数量、大小均一性和栽培产量,其中基部分枝型花序果实成熟更均匀,是商业化生产的优异性状,但草莓花序节间伸长的遗传调控机制长期未被解析。
2026年5月6日,《The Plant Cell》在线发表了华中农业大学康春颖教授团队题为“The Gα protein FveGPA1 regulates inflorescence architecture via the FveBRI1-mediated brassinosteroid pathway in strawberry”的研究论文。该研究以二倍体林地草莓为模式材料,通过诱变筛选获得4个基部分枝花序突变体,结合图位克隆、遗传互作、激素处理与多组学分析,解析了异三聚体G蛋白α亚基FveGPA1通过油菜素内酯(BR)受体FveBRI1介导的信号通路,协同调控花序节间差异伸长的分子机制,为草莓花序构型的定向改良、实现果实均匀生产和机械化采收提供了重要的理论基础与基因靶点。
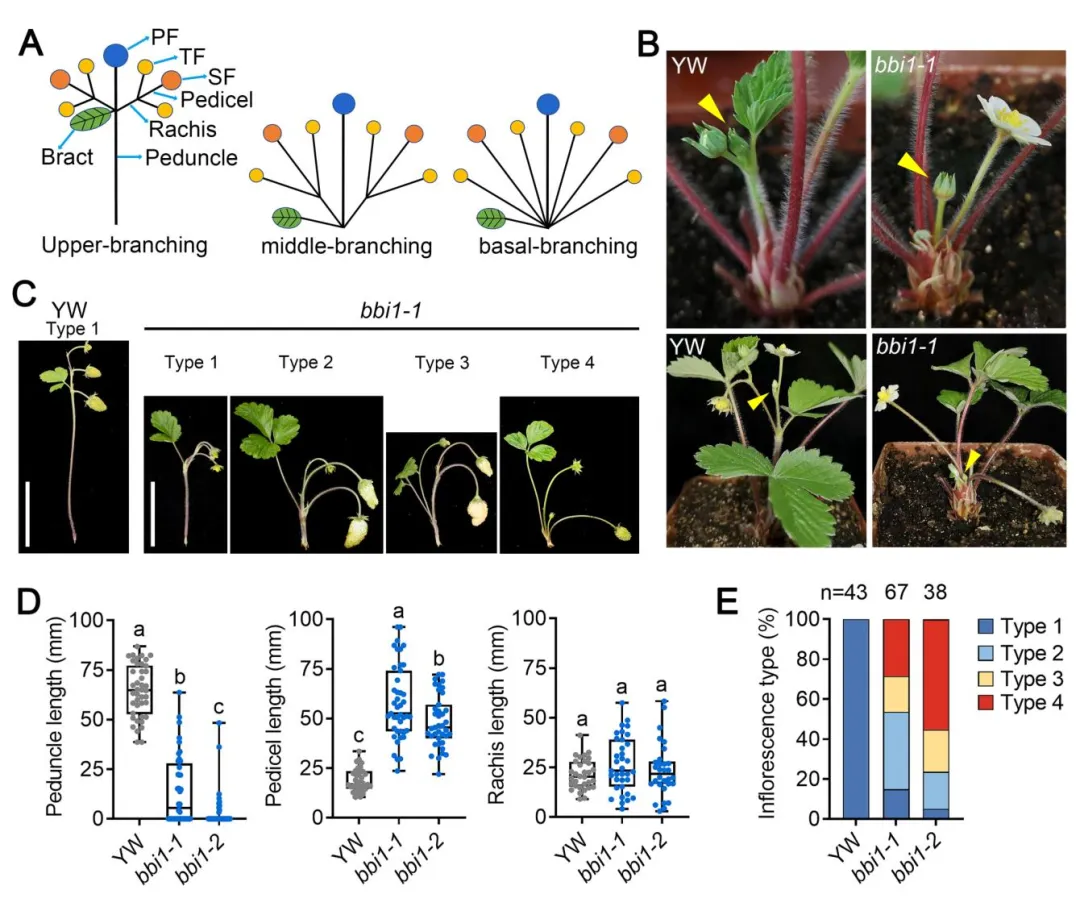
▎1. 基部分枝花序突变体bbi1的表型鉴定
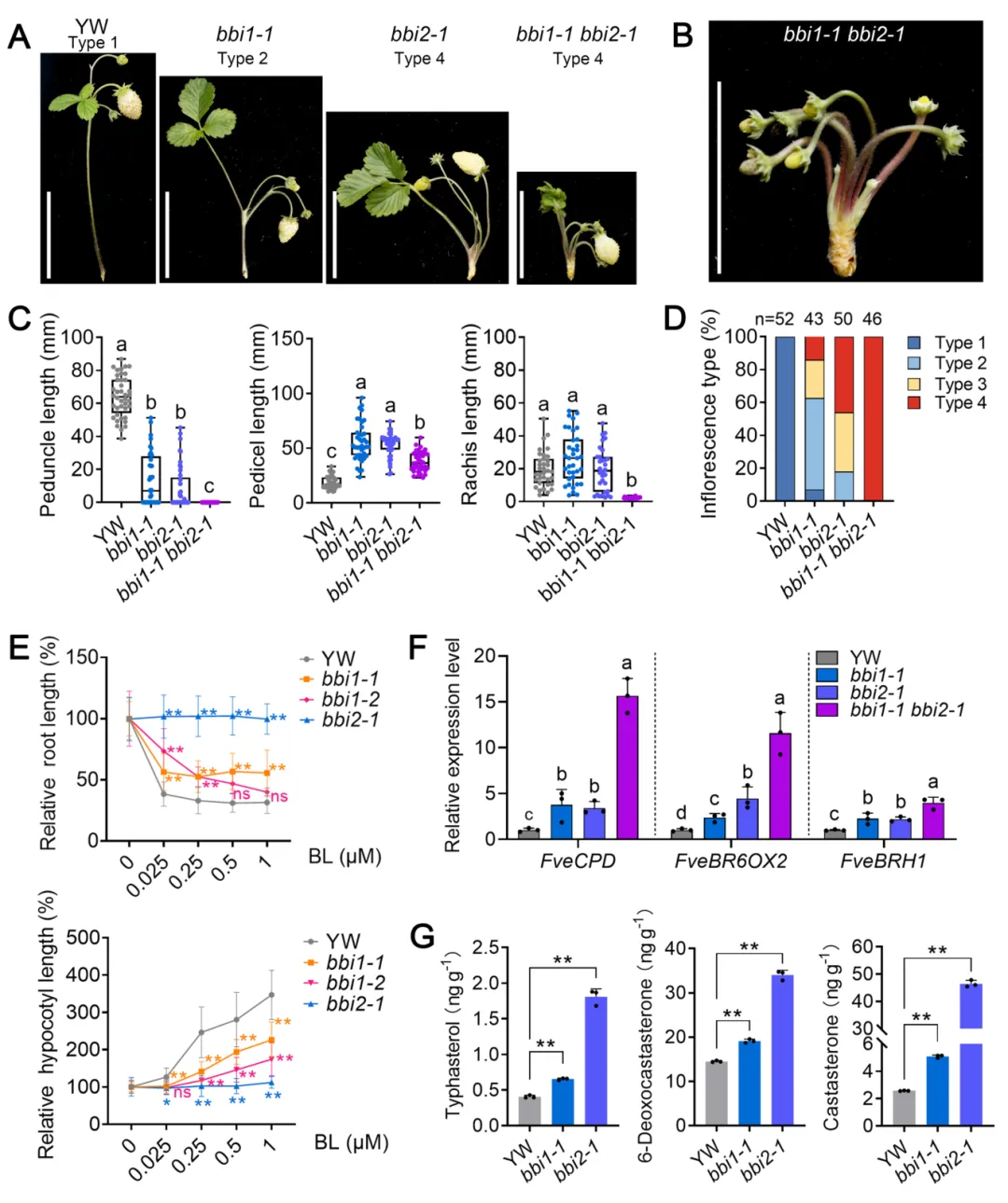
野生型林地草莓‘Yellow Wonder’为典型上部分枝花序,具有长花梗(支撑整个花序的主茎);而EMS诱变获得的两个等位突变体bbi1-1和bbi1-2的花梗显著缩短、花柄(支撑单朵花的茎)伸长,呈现部分基部分枝表型。研究将草莓花序分为4类:类型1(花梗长于花柄,野生型)、类型2(花梗短于花柄)、类型3(无花梗但一级果与二级果间有轴结构)、类型4(无花梗也无一级与二级果间轴)。统计显示bbi1-1中仅14.92%为野生型的类型1,bbi1-2中55.26%为最严重的类型4,同时突变体还伴随叶片变圆、叶柄缩短等多组织发育缺陷。
图1 林地草莓bbi1突变体的花序结构表型鉴定
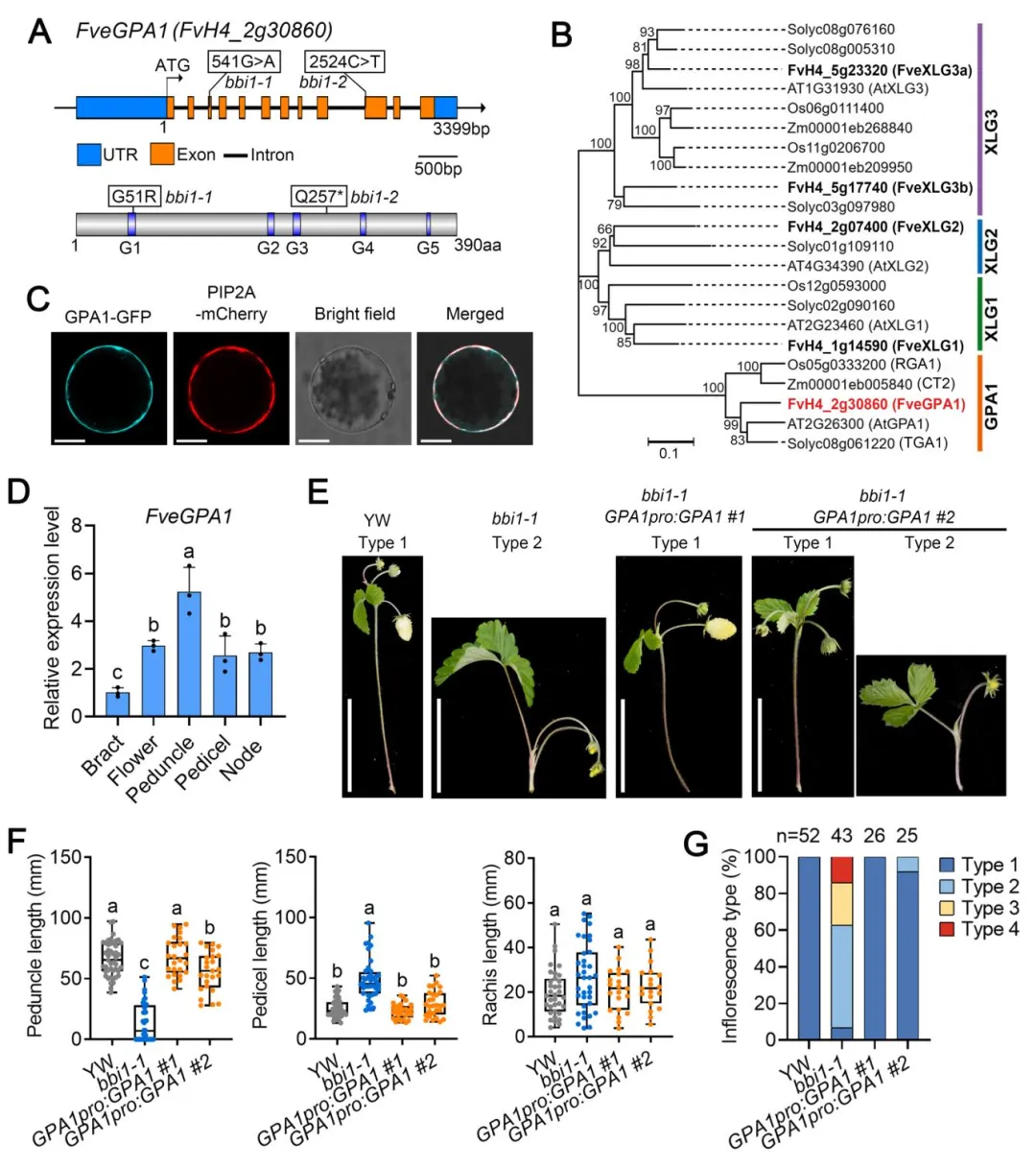
▎2. 图位克隆证实FveGPA1是调控花序结构的关键基因
通过回交群体遗传分析,证明bbi1突变表型由单隐性核基因控制。结合混池重测序定位到候选基因为编码异三聚体G蛋白α亚基的FveGPA1:bbi1-1中该基因发生G-to-A点突变,导致第51位甘氨酸突变为精氨酸;bbi1-2中发生C-to-T点突变,产生提前终止密码子。系统发育分析显示FveGPA1是林地草莓中唯一的经典Gα亚基,定位于细胞质膜,在花梗中表达量最高。遗传互补实验表明,转入FveGPA1全长序列可完全恢复bbi1-1的花序表型;CRISPR敲除FveGPA1也能获得与bbi1类似的基部分枝花序表型,证实FveGPA1是调控草莓花序结构的关键基因。
图2 FveGPA1的基因克隆与功能验证
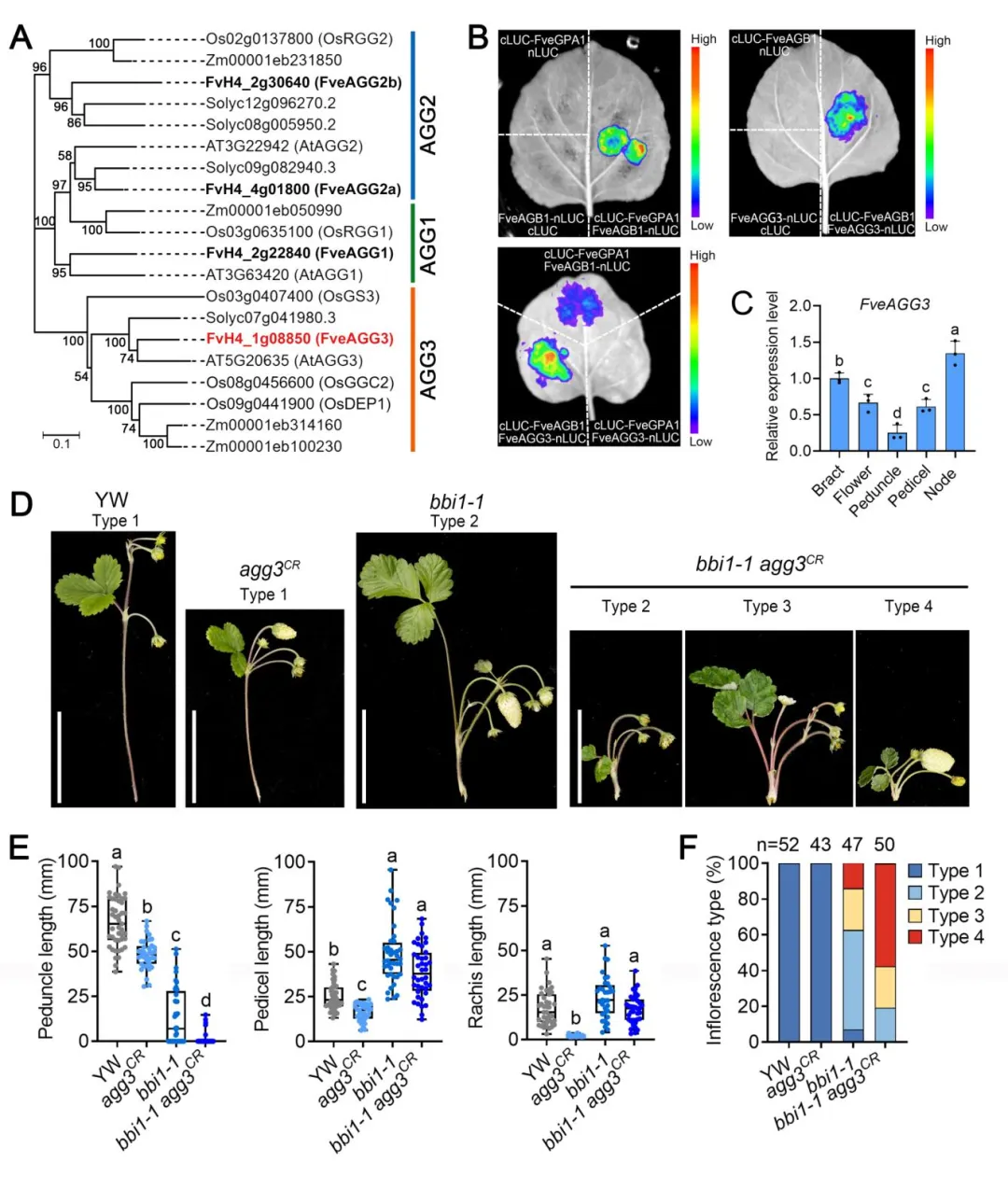
▎3. G蛋白γ亚基FveAGG3与FveGPA1协同调控花序发育
林地草莓基因组中共鉴定到4个Gγ亚基编码基因,其中FveAGG3与拟南芥、水稻中调控器官发育的AGG3同源。Split-LUC实验证实FveGPA1与Gβ亚基FveAGB1互作,FveAGB1进一步与FveAGG3互作,三者可形成完整的异三聚体G蛋白复合体。CRISPR敲除FveAGG3获得的agg3CR突变体同样表现为花梗缩短、花序基部分枝表型;构建bbi1-1 agg3CR双突变体后,其基部分枝表型显著增强,类型4花序占比从bbi1-1的13.95%提升至57.69%,说明FveAGG3与FveGPA1在花序结构调控中存在协同效应。
图3 FveAGG3与FveGPA1的遗传互作分析
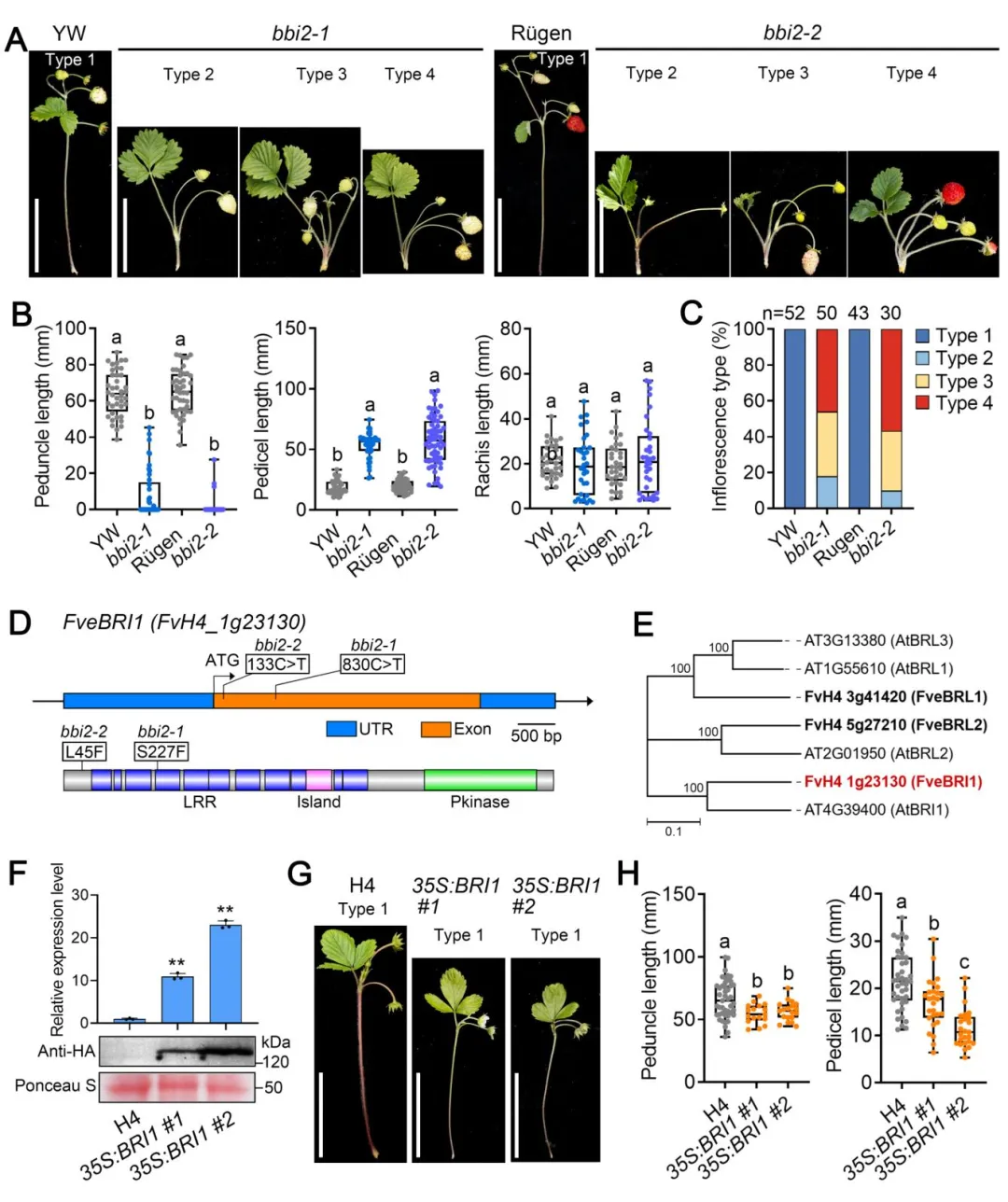
▎4. BR受体编码基因FveBRI1是bbi2突变体的致病基因
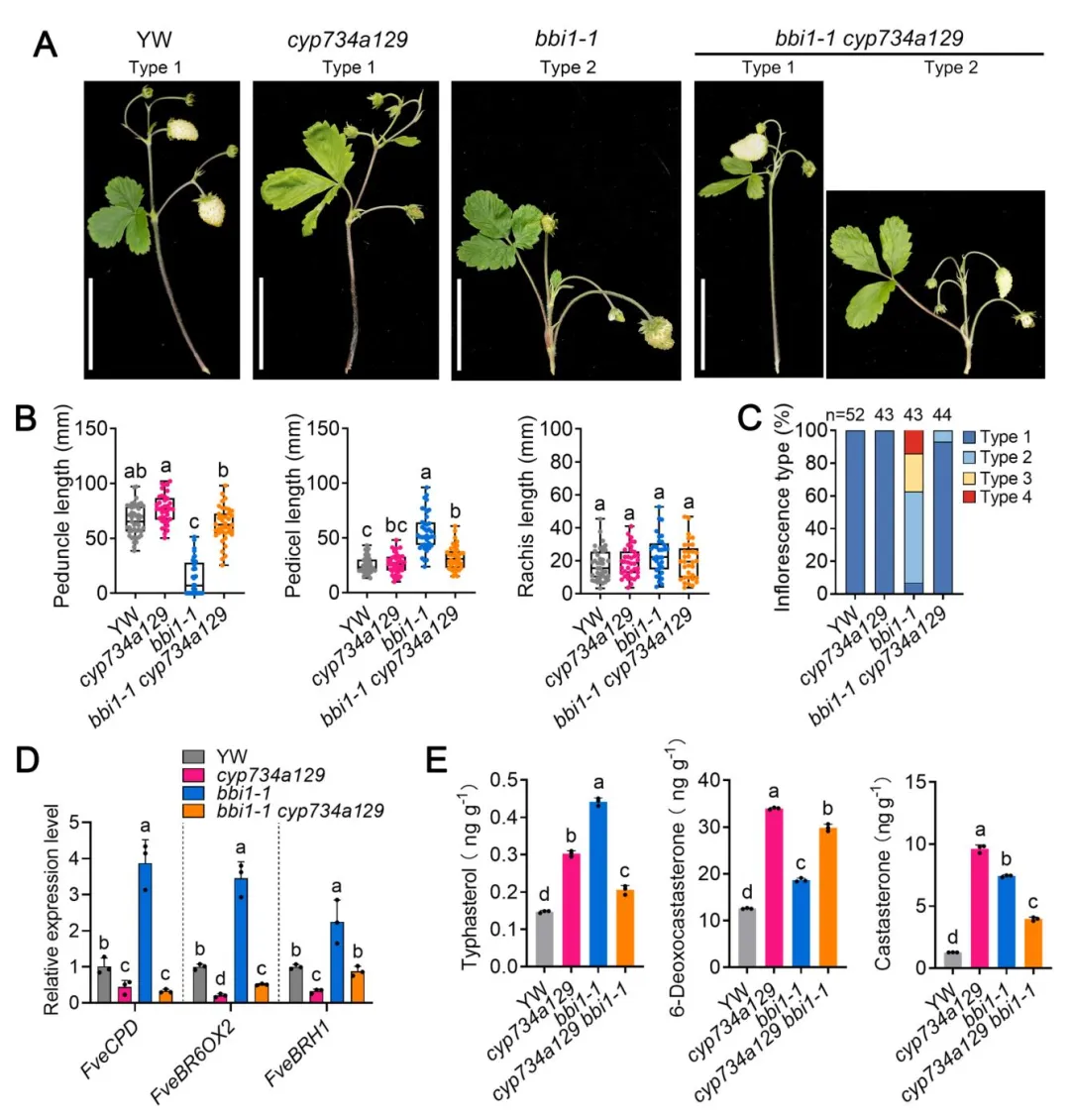
另外两个等位突变体bbi2-1(YW背景)和bbi2-2(Rügen背景)表现出与bbi1高度类似的基部分枝花序表型,其中bbi2-2表型更严重,类型4花序占比更高。通过图位克隆定位到候选基因为BR受体编码基因FveBRI1:bbi2-1中发生第227位丝氨酸到苯丙氨酸的点突变,bbi2-2中发生第45位亮氨酸到苯丙氨酸的点突变。过表达FveBRI1的转基因株系花柄显著缩短,叶片变窄、叶柄伸长,与bbi2突变体表型相反,证实FveBRI1同样是调控草莓花序结构的关键因子。
图4 bbi2突变体的表型鉴定与FveBRI1基因克隆
▎5. FveGPA1通过BR信号通路调控花序结构
构建bbi1-1 bbi2-1双突变体,其表型呈现显著协同增强效应:所有花序均为类型4,完全丧失花梗和果轴,花直接从茎上长出,表明FveGPA1与FveBRI1存在遗传互作。进一步实验证实:①bbi1突变体对BR处理的敏感性显著降低,类似BR不敏感突变体bbi2;②bbi1和bbi2突变体中BR合成基因(FveCPD、FveBR6OX2)和响应基因(FveBRH1)均显著上调,内源BR代谢物(香蒲甾醇、6-脱氧油菜素甾酮、油菜素甾酮)含量显著升高,存在典型的BR信号受损后的反馈上调;③将bbi1-1与BR分解代谢酶基因突变体cyp734a129杂交后,双突变体的花序表型几乎完全恢复,93.18%的花序回到野生型的类型1,BR信号相关基因的表达也恢复至野生型水平,充分证明FveGPA1通过调控BR信号通路参与花序结构调控。
图5 FveGPA1与BR通路的遗传互作验证
图6 cyp734a129对bbi1突变体表型的恢复效应
▎6. FveGPA1与FveBRI1通过调控细胞分裂和扩张调节节间伸长
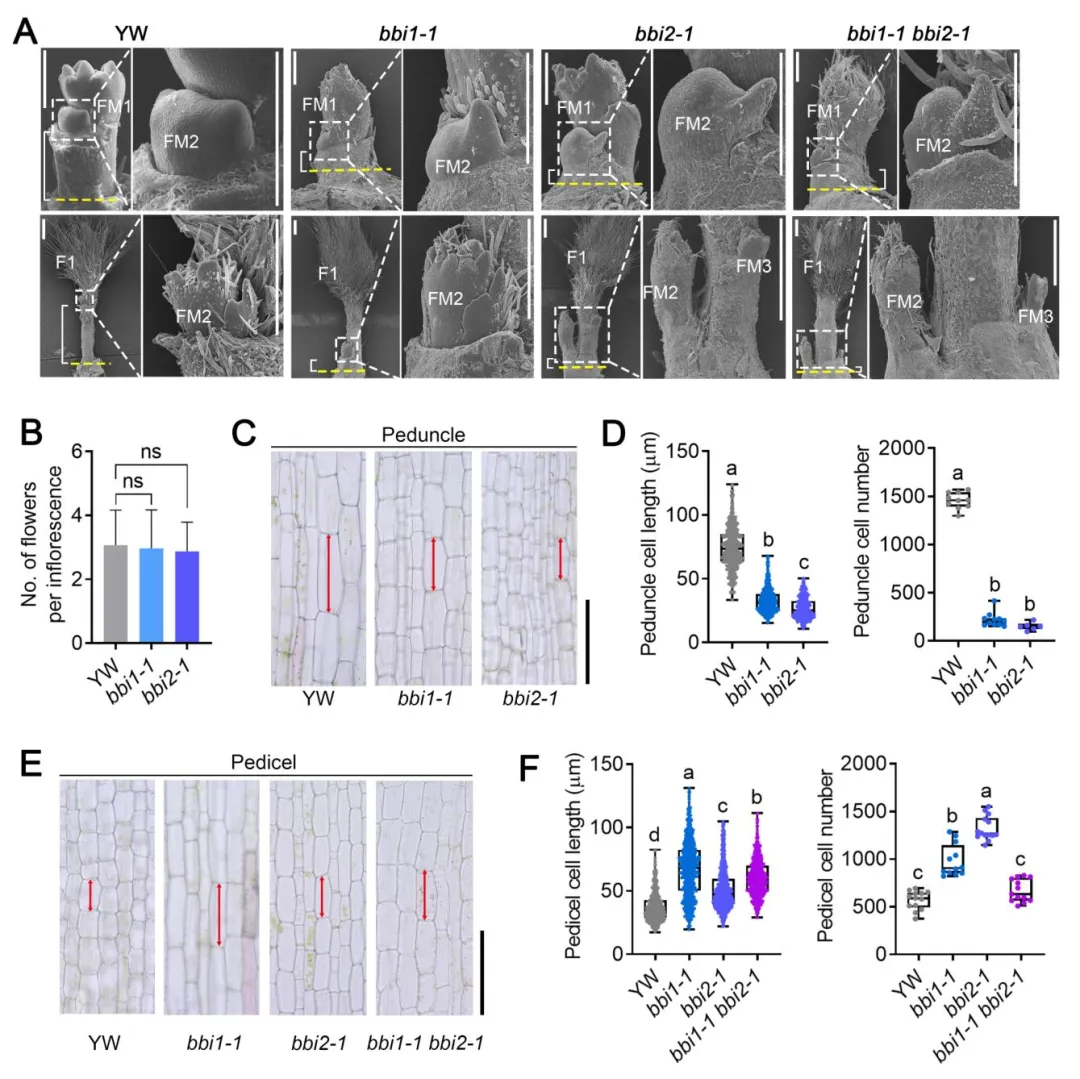
扫描电镜观察显示,突变体与野生型的花序分生组织发育、单花序花数量无显著差异,花序结构的差异来源于节间伸长过程的不同。对成熟花梗和花柄的纵切观察发现:bbi1和bbi2突变体的花梗皮层细胞长度显著缩短、细胞数量减少,导致花梗变短;而花柄的皮层细胞长度显著增加、细胞数量增多,导致花柄伸长,说明FveGPA1和FveBRI1通过组织特异性地调控细胞分裂和扩张,实现花梗和花柄的差异伸长,最终决定花序构型。
图7 FveGPA1与FveBRI1对花梗和花柄细胞发育的调控
▎7. 转录组分析揭示FveGPA1与FveBRI1的共同下游调控网络
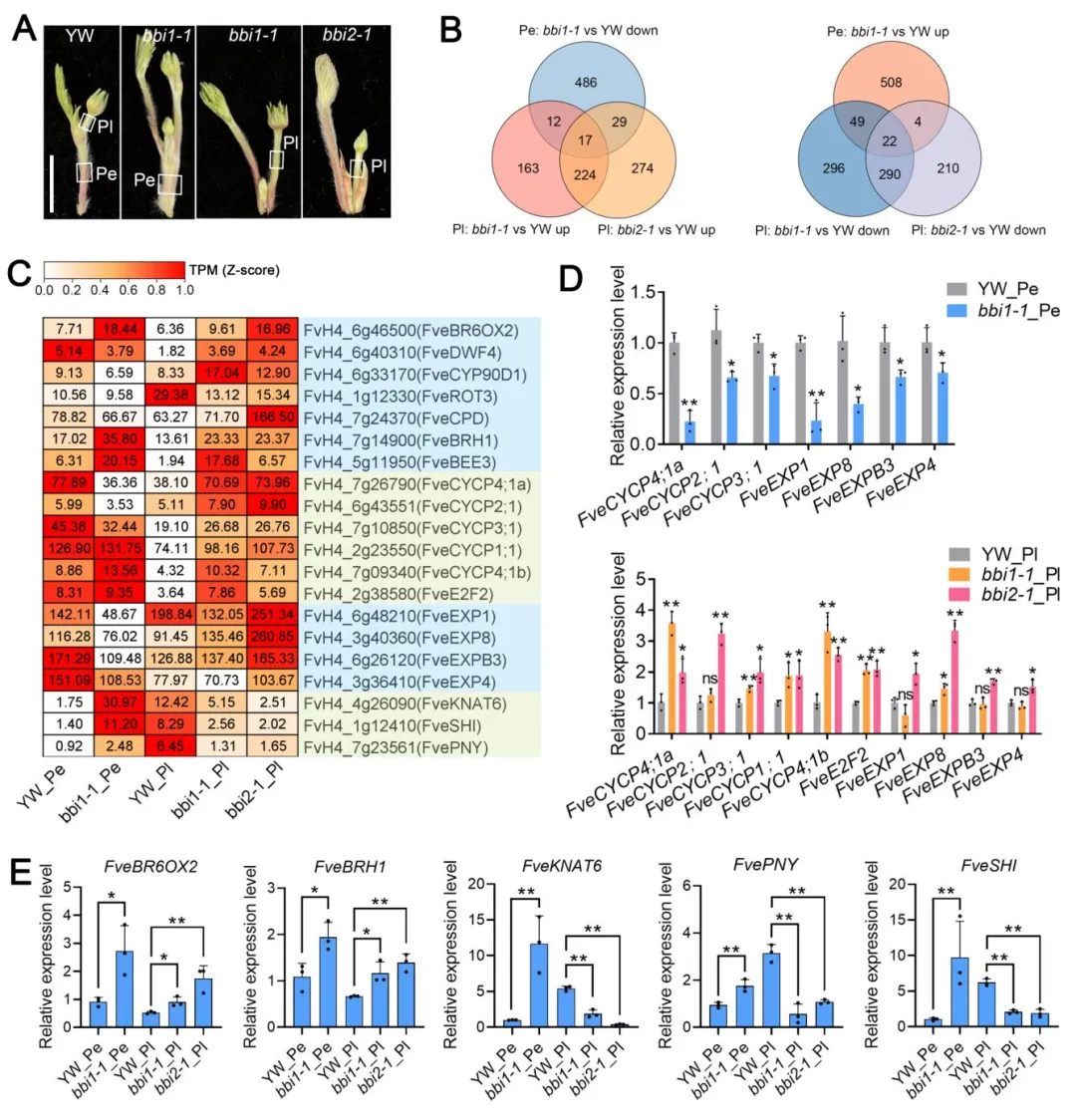
对野生型和突变体的发育中花梗、花柄进行转录组测序,发现bbi1和bbi2突变体的花柄中共有52%的差异表达基因重叠,说明二者共享大量下游调控靶点。其中BR合成与响应基因在两个突变体中均显著上调,与生理实验结果一致;细胞周期基因(如CYCP家族)和细胞壁扩张基因(如EXP家族)在突变体花梗中下调、花柄中上调,对应了二者细胞发育的差异;此外,调控节间伸长的关键转录因子FveKNAT6、FvePNY和FveSHI在突变体花梗中上调、花柄中下调,可能是介导组织特异性伸长的关键调控因子。
图8 花梗与花柄的转录组比较分析
▎8. 草莓花序结构调控的分子模型
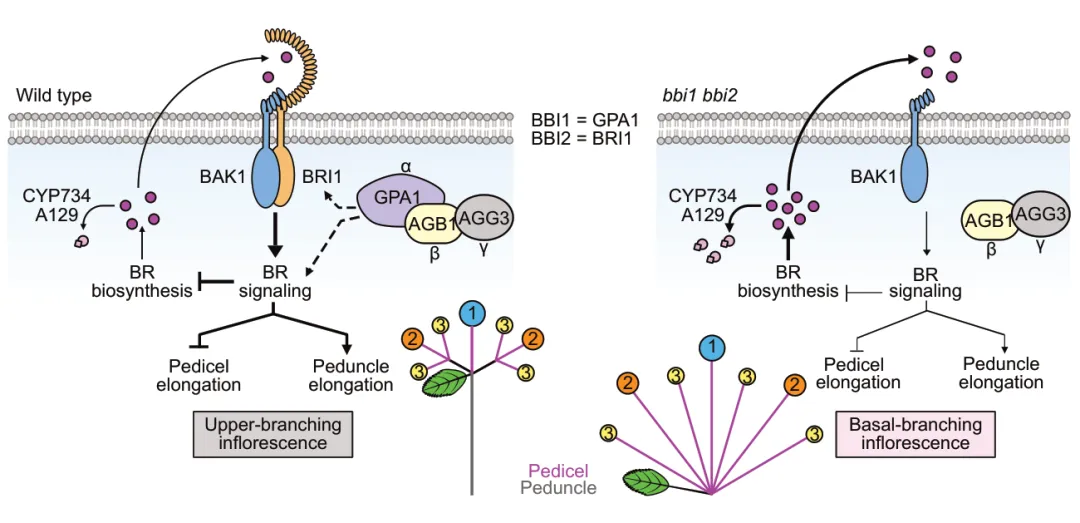
基于上述结果,研究团队提出了草莓花序结构的调控模型:BR信号通路通过受体FveBRI1调控花序节间伸长,异三聚体G蛋白复合体(FveGPA1-FveAGB1-FveAGG3)通过与BR通路的组分互作,精细调控BR信号输出,促进花梗伸长、抑制花柄伸长,最终形成上部分枝花序;当FveGPA1或FveBRI1功能缺失时,BR信号受损,花梗伸长受抑、花柄过度伸长,花序转变为基部分枝型。
图9 FveGPA1与FveBRI1调控草莓花序结构的工作模型
本研究突破了以往花序调控研究主要聚焦于分生组织活性的局限,重点解析了节间差异伸长这一决定花序结构的关键维度,首次揭示了异三聚体G蛋白与油菜素内酯通路协同调控草莓花序构型的分子机制。基部分枝花序具有果实大小均匀、成熟一致的优势,更适合商业化生产和机械化采收,本研究鉴定的FveGPA1、FveBRI1等基因为草莓花序构型的定向遗传改良提供了精准靶点。未来可进一步挖掘草莓种质资源中相关基因的自然变异,开发功能分子标记,快速选育具有适宜花序结构的优良草莓品种;同时解析G蛋白与BR通路互作的具体分子细节,明确组织特异性调控的上游信号,为草莓株型和产量性状的协同改良提供理论支撑。
---原文链接---:https://doi.org/10.1093/plcell/koag134
为了能更有效地帮助广大的科研工作者获取相关信息,植物生物技术Pbj特建立微信群,PlantBiotechnologyJournal投稿以及文献相关问题、公众号发布内容及公众号投稿问题都会集中在群内进行解答,同时鼓励在群内交流学术、碰撞思维。为了保证群内良好的讨论环境,请先添加小编微信,扫描二维码添加,之后我们会及时邀请您进群。小提示:添加小编微信时及进群后请务必备注学校或单位+姓名,PI在结尾注明,我们会邀请您进入PI群。