腐败微生物是鲜肉变质的主要原因。尽管腐败通常始于表面,但随后微生物向内部的渗透是一个关键步骤,它会加速保质期的缩短并促进深层组织的变质。以往的研究大多将渗透能力视为单个菌株的固有特性(Gill 和 Penney,1977;Shirai 等,2017)。然而,在新鲜猪肉系统中,细菌通常以群落形式共存,种间或种内相互作用可能通过改变营养物质的可用性和组织结构,从而重塑穿透过程(Fang 等,2022)。本研究选取了两种运动表型不同的P. lundensis菌株,旨在探究共培养如何影响其从猪肉表面向深层组织的穿透,并进一步分析该过程与腐败发展的关系。
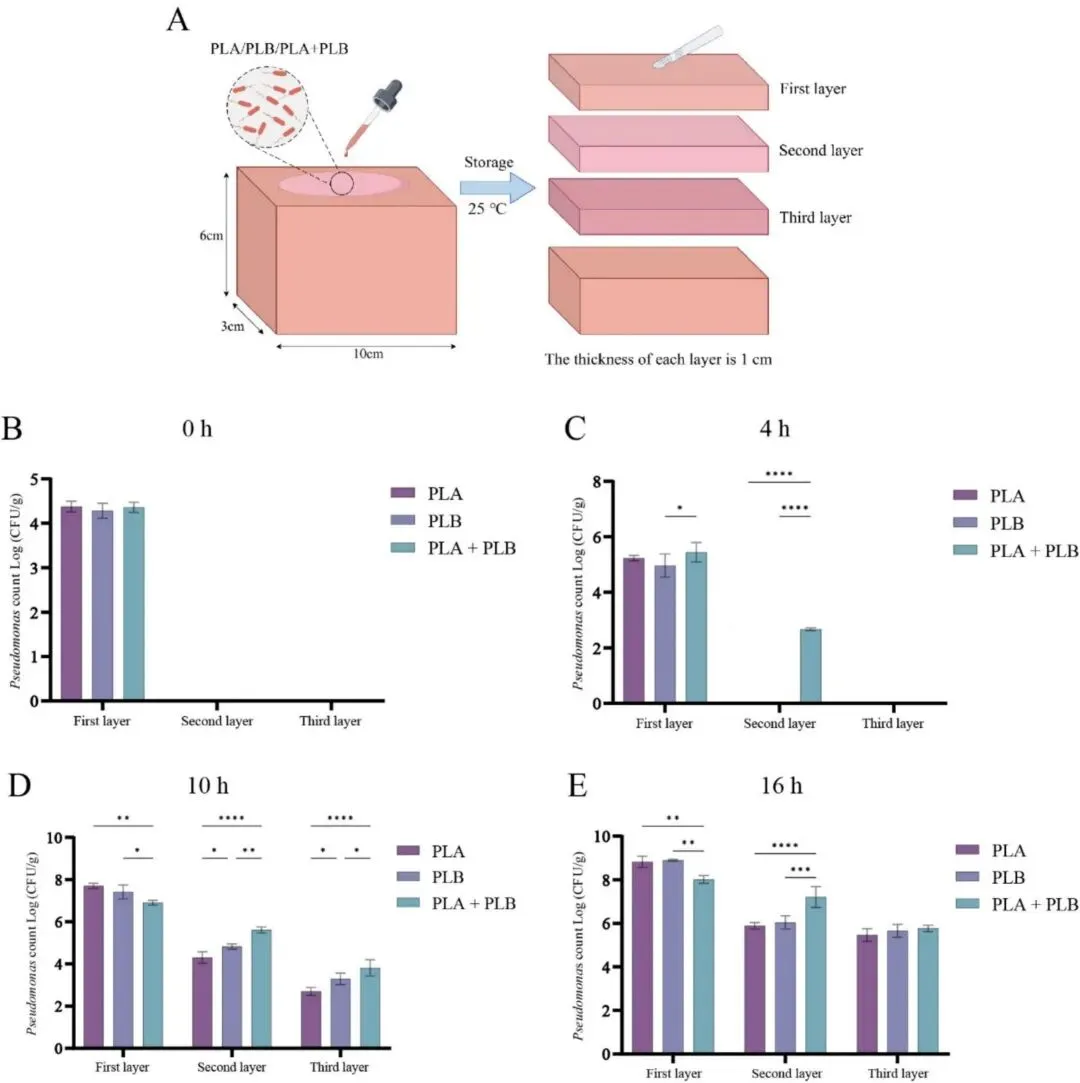
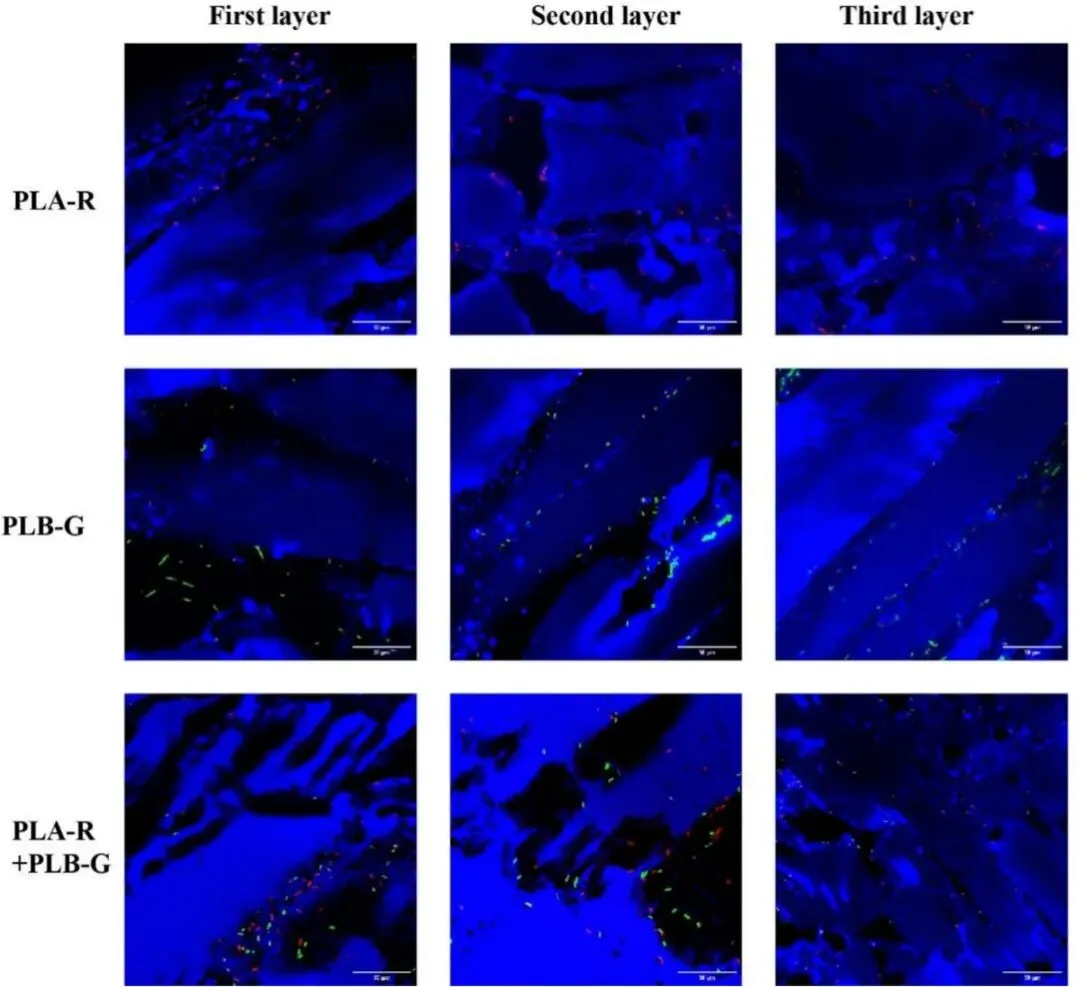
分层计数结果显示,PLA + PLB 组比任何单一培养组更早出现在深层组织中,并在储存期间第二层的细菌计数保持更高,这表明共培养加速了向内的渗透及随后的定植。在其他多物种共培养系统中也报道了类似现象,这表明细菌相互作用可以增强渗透过程(Ali 等,2022)。各层中的假单胞菌数量随时间推移稳步增加,这主要是因为猪肉为细菌的生长和定植提供了营养丰富的环境。随着细菌负荷的增加,假单胞菌很可能分泌了更多的蛋白水解酶,从而更有效地水解肌肉组织中的肌浆蛋白,削弱组织结构,并进一步促进了渗透。共聚焦激光扫描显微镜(CLSM)观察显示,P. lundensis 主要沿肌纤维间隙和结缔组织结构向内渗透,这与先前关于细菌通过肌肉间质空间侵入肉类内部的观察结果一致(Bosse 等,2015)。在 PLA + PLB 组中,这两种菌株经常被检测到位于肌纤维间隙内的相邻区域,表明存在高度协调的空间共定植模式。这种空间共定位可能有利于细菌协同利用局部营养资源,并增强其在组织内的持续定植。
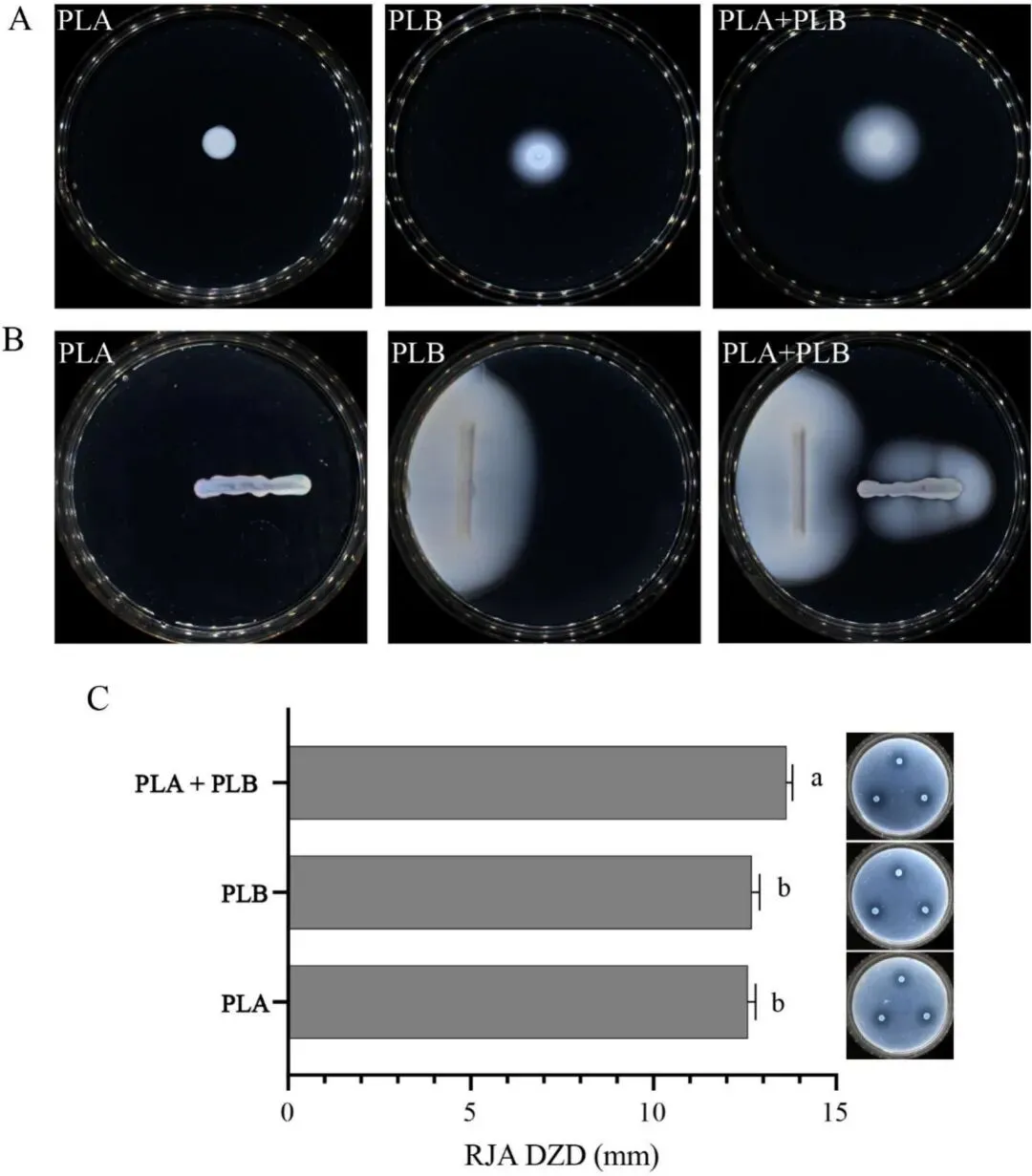
体外表型结果进一步证实了这种共渗透现象。半固体平板上的扩散范围反映了细菌的运动能力,而RJA水解区的直径则反映了细菌降解肉类基质的能力。细菌的渗透与运动能力和蛋白水解活性密切相关。这两项特性使细菌能够穿过肌纤维间的空隙,并通过产生蛋白水解酶来降解结缔组织,从而污染更广、更深的组织区域(Moza 等,2009;SIKES 和 MAXCY,1980)。Hagai 等(2013)报道称,在设计相似的平板共培养系统中,假单胞菌被黄单胞菌吸引,导致扩散区域扩大。同样,本研究中共培养条件下 PLA 扩散范围的增加表明,菌株间相互作用改变了细菌的迁移行为。我们的结果还显示,共培养产生了更大的RJA水解区,表明混合培养中的蛋白水解活性更强,这与Dou等人(2025)的发现一致。综合来看,这些特征可能是共培养组在猪肉模型中表现出更强渗透能力的基础。
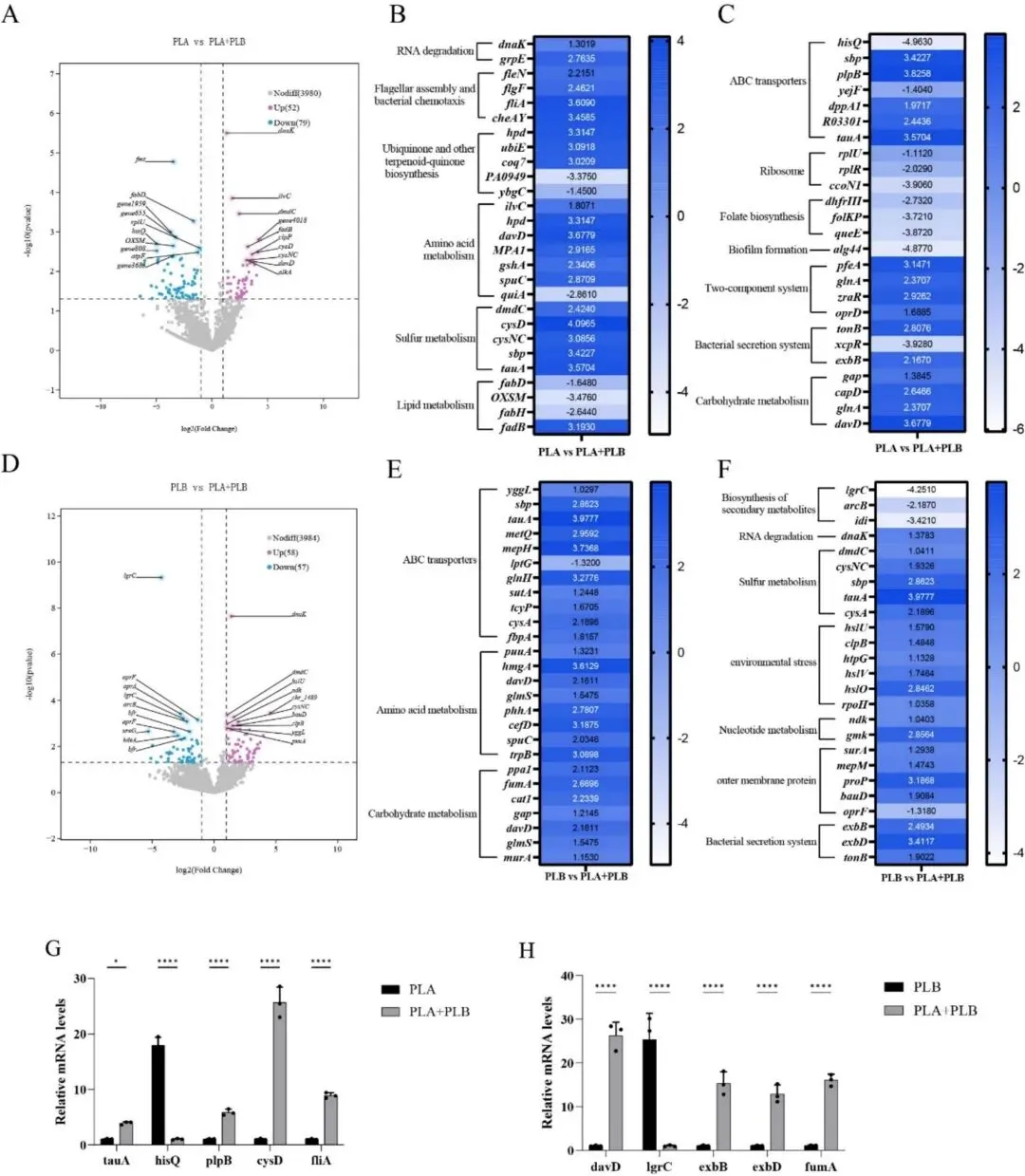
为了阐明共培养条件下观察到的穿透能力增强背后的潜在分子机制,进一步进行了转录组分析。一方面,共培养显著上调了PLA中与运动性和趋化性相关的基因,包括鞭毛组装基因fleN、flgF和fliA,以及趋化性基因cheA和cheY。这些基因编码的蛋白质不仅是鞭毛装置的关键结构组分,对营养获取和细胞表面相互作用也至关重要(Khan 等,2020)。这些基因的上调表明PLA中的运动能力和营养梯度感知均被激活,这与体外观察到的扩散增强表型一致。另一方面,在两种菌株共培养期间,与碳水化合物代谢、硫代谢、氨基酸代谢以及多种ABC转运系统相关的基因均呈现上调。由于原核生物的ATP生成主要依赖于糖酵解过程中的底物级磷酸化和氧化磷酸化(Wang et al., 2021),这些代谢和转运相关基因的上调表明共培养条件下能量需求更高,这与本研究中观察到的更旺盛的生长和菌落计数结果一致。ABC转运系统是微生物中普遍存在的跨膜转运系统,在营养物质的摄取和利用中发挥关键作用(Ekiert 等,2022)。因此,这些转录变化表明,在肉类组织内侵入及随后的生长过程中,这两种菌株可能具备更强的营养获取和代谢适应能力。
作为蛋白质的基本组成单位,氨基酸会发生脱氨反应,这是腐败菌降解和利用肉源氨基酸的重要途径(Zhuang 等,2021)。共培养条件下氨基酸代谢相关基因的广泛上调表明,PLA + PLB 组中的蛋白质降解和氨基酸异化作用更为活跃。与此一致,Zhang等(2025)报道称,将两种假单胞菌菌株共同接种至新鲜猪肉中,导致尿苷等代谢物显著积累,表明这两种腐败菌通过包括组氨酸代谢在内的多种氨基酸分解代谢途径共同降解营养物质。Zhou 等(2024)还发现,P. fragi 与 Brochothrix thermosphacta 的共培养可能通过组氨酸和嘌呤代谢产生协同效应,从而促进氨基酸和核苷酸的降解,增加微生物负荷和 TVB-N 含量,并加速变质。综合来看,转录组学证据表明,共培养赋予了 P. lundensis 更强的运动能力以及获取和利用蛋白质降解产物的更大能力,从而支持其在肉质基质中的快速增殖和持续代谢活动。
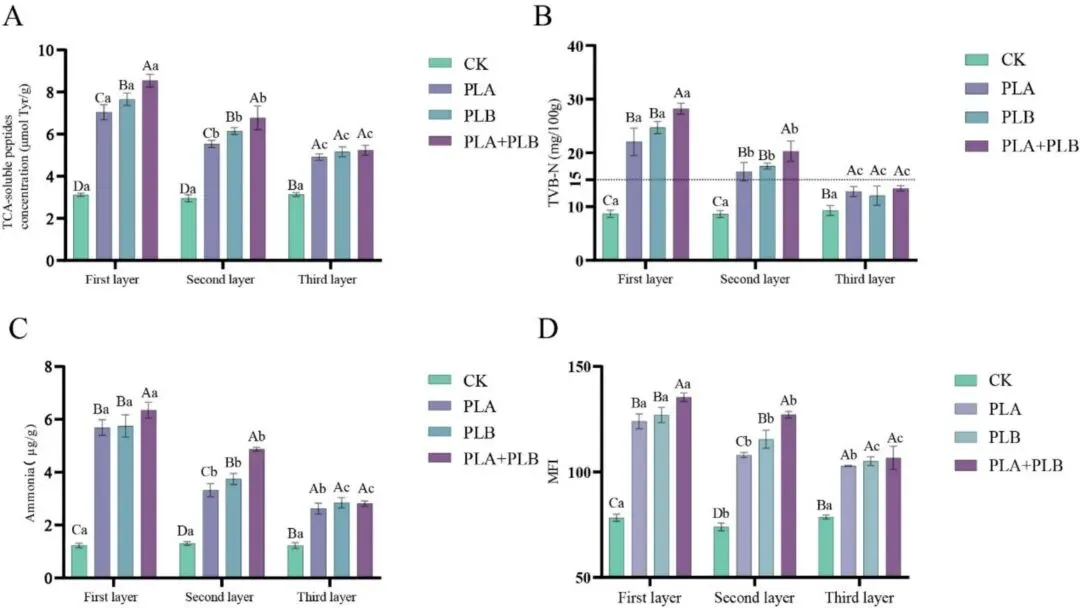
因此,在渗透期结束时,对猪肉中的蛋白质利用率和肌原纤维损伤进行了评估。与任何单一培养组相比,PLA + PLB 组表层中的 TCA 可溶性肽、TVB-N 和氨含量均显著更高,这表明 PLA + PLB 组中与蛋白质转运和氨基酸代谢相关的基因上调伴随着肉中可溶性降解产物和氨的大量积累。MFI结果进一步表明,共培养对肌原纤维的破坏作用更强,这与其更强的腐败潜力相一致。Jia 等(2024)也报告称,在嗜冷假单胞菌与腐败链球菌共培养期间,微生物负荷增加和蛋白酶分泌破坏了肌原纤维的横向交联和结构完整性,导致碎片化指数升高。本研究的扫描电子显微镜(SEM)观察结果进一步支持了这一结论。这些发现表明,随着蛋白质被水解及组织结构逐渐松弛,猪肉组织的结构抗性降低,这可能进一步促进细菌向深层区域迁移。因此,组织降解不仅是细菌定植的结果,还可能进一步助长了细菌的向内扩散。
综合来看,分层计数、共聚焦激光扫描显微镜(CLSM)观察、体外表型及转录组学数据均表明,这两种P. lundensis菌株的共渗透是由多种协调过程驱动的。在共培养条件下,猪肉表面的空间共存伴随着更强的迁移表型和更明显的空间共定植。与此同时,与营养获取、跨膜转运及能量代谢相关的变化,可能促进了该菌在组织内的持续生长和适应。蛋白质降解产物的积累以及更明显的组织损伤进一步表明,共侵过程与加剧的腐败活动紧密相关。因此,PLA与PLB之间的协同效应不太可能体现在单一性状的增强上;相反,它更可能源于同一肉类微环境中渗透、定植和代谢利用之间的协调相互作用。未来的研究应验证参与趋化行为、营养物质摄取和蛋白酶活性的关键基因的功能,以进一步阐明这些过程在共渗透中的具体作用及其协同关系。