禽传染性支气管炎病毒(IBV)是全球家禽业最重要的呼吸道病原体之一,其疫苗生产长期依赖SPF鸡胚,面临供应受限、批次差异大及外源因子污染等瓶颈。尽管哺乳动物细胞培养技术已在多种疫苗生产中广泛应用,IBV因严格的宿主嗜性——仅能感染禽源细胞,无法在任何哺乳动物细胞中有效复制——始终未能实现细胞培养疫苗的转化。
2026年4月9日,中国农业大学研究团队在PLOS PATHOGENS上发表题为“Accumulation of mutations in nsp4, E, and the S2 subunit underlies mammalian cell tropism expansion and virulence attenuation of avian coronavirus”的最新研究论文。研究结果表明,IBV中的哺乳动物细胞嗜性扩增与预先存在的变体的逐步选择有关,导致S蛋白和其他结构和非结构蛋白的协调突变,从而降低致病性
IF:5.5 中科院1区 | JCR/Q1 微生物学 参考译文:Nsp4、E和S2亚基突变的积累是禽冠状病毒哺乳动物细胞嗜性扩增和毒力减弱的基础 第一作者:Yingfei Li1,Xuehui Zhang 通讯作者:Guozhong Zhang , Jing Zhao |
1. IBV对哺乳动物细胞的适应
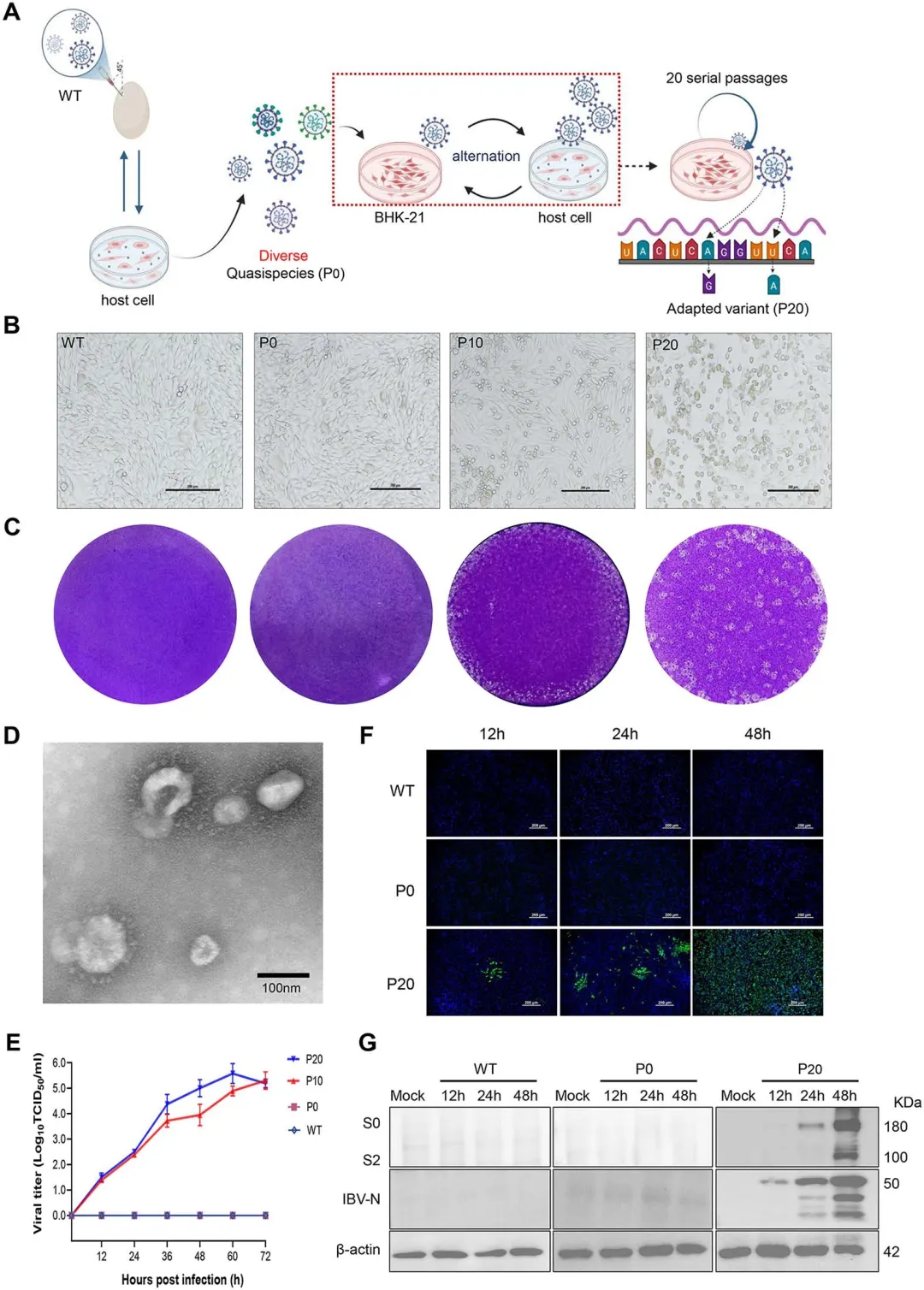
野生型IBV先在鸡胚和原代鸡胚肾(CEK)细胞中连续传代15次,构建遗传多样性丰富的准种群体(P0);随后将该准种在BHK-21细胞与CEK细胞之间交替传代,利用CEK细胞的扩增优势挽救BHK-21细胞中有限吸附的病毒粒子,防止病毒在非宿主细胞中丢失。至第10代(P10)时,BHK-21细胞开始出现细胞病变效应(CPE),表现为细胞圆缩;至第20代(P20)时CPE显著加剧并伴随细胞脱落。蚀斑试验显示P20病毒蚀斑较P10更大且边界清晰,透射电镜观察到典型冠状病毒形态颗粒。P20病毒在感染后60小时达到复制峰值,滴度达10^5.57 TCID50/ml,而野生型及P0准种在BHK-21细胞中均无 detectable 复制。间接免疫荧光和Western blot证实P20病毒S2及N蛋白表达随时间递增,野生型则无表达。
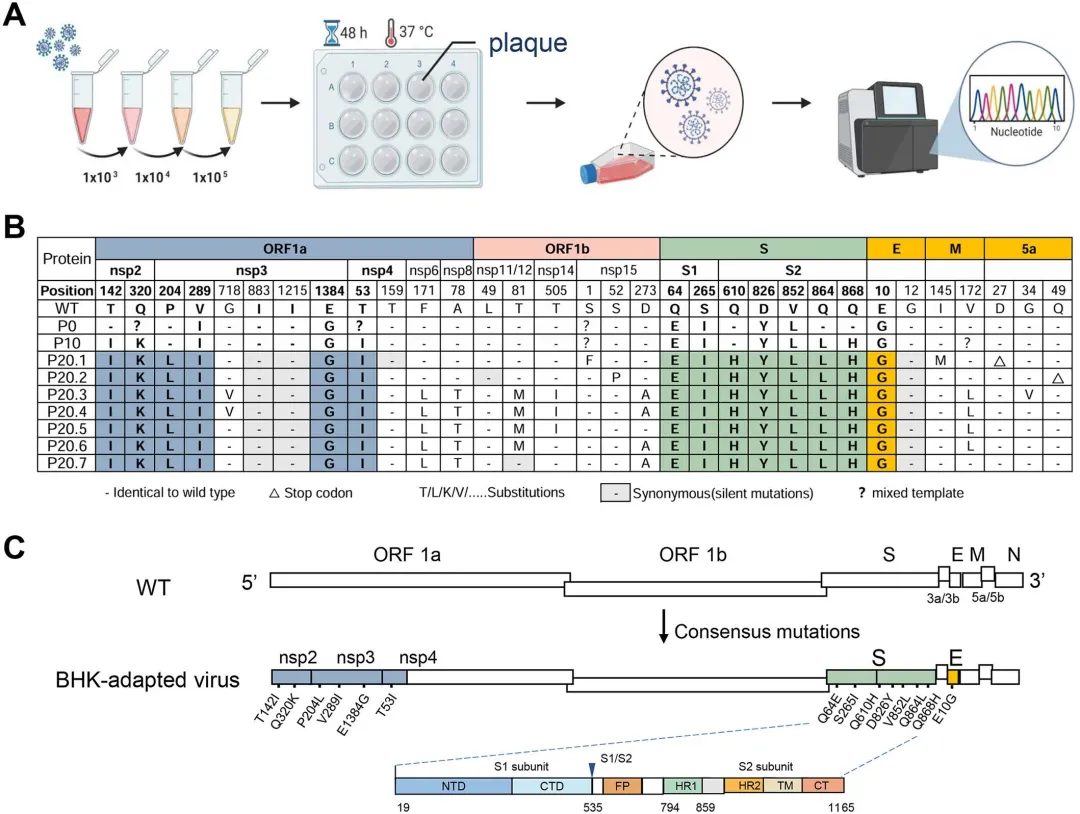
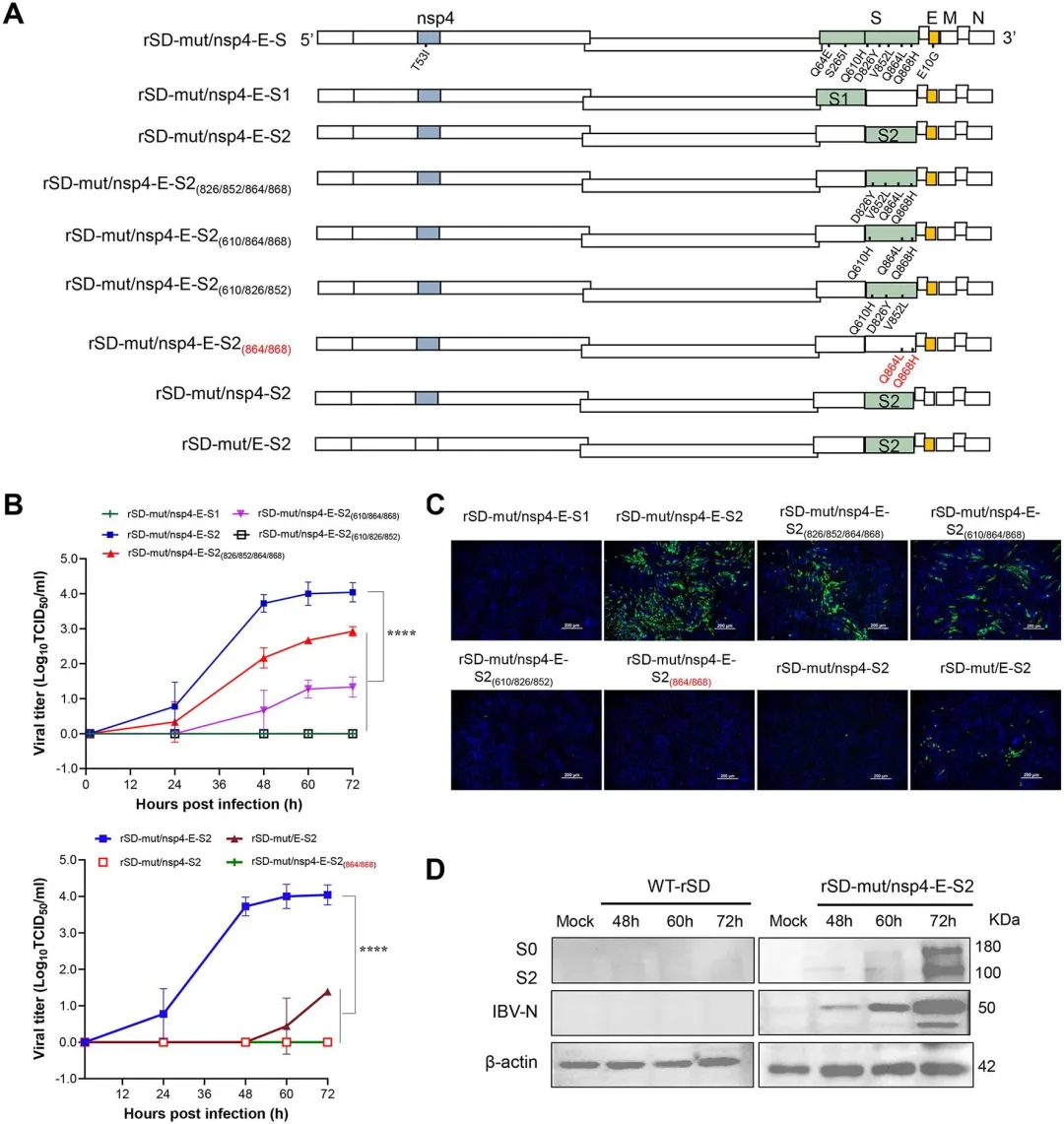
为精确定位与细胞嗜性相关的关键突变,研究者通过三轮蚀斑纯化获得7个单克隆病毒株(P20.1-P20.7),经全基因组PCR扩增和Sanger测序后,与野生型及P0准种进行比对分析。结果显示,从WT到P0阶段已在胚胎蛋和CEK细胞传代过程中积累了大量突变;7个蚀斑克隆携带分布于非结构蛋白和结构蛋白(除N蛋白外)的不同突变组合,其中5a辅助基因突变导致翻译提前终止。进一步分析鉴定出14个共有突变,分布于nsp2(2个)、nsp3(3个)、nsp4(1个)、S基因(7个)及E基因(1个),其中S蛋白尤其是S2亚单位成为突变富集的热点区域。图2.IBV对BHK-21细胞适应性的全基因组突变分析3.S蛋白对于IBV细胞适应是必需的,但不是充分的,需要与Nsp4(T53I)和E(E10G)合作
将P20的全部14个共有突变引入野生型SD骨架获得rP20,该病毒可在BHK-21细胞高效复制(10^4.8 TCID50/ml);而仅替换S基因的嵌合病毒rSD-SP20或保留非S区突变的rP20-SSD均无法复制,证明S蛋白单独不足以驱动IBV跨宿主适应。为筛选关键非S蛋白突变,研究者对nsp2(T142I, Q320K)、nsp3(P204L, V289I, E1384G)、nsp4(T53I)及E(E10G)进行单点回复突变:nsp4回复至野生型T53使病毒滴度降低约40倍,E蛋白回复至E10使滴度下降约79倍,而nsp2和nsp3的回复不影响复制能力。进一步将nsp4(T53I)、E(E10G)与S突变共同引入rSD骨架构建rSD-mut/nsp4-E-S,其复制效率恢复至rP20水平。免疫荧光试验证实rP20-nsp4^SD和rP20-E^SD荧光信号显著减弱。图3.结构蛋白和非结构蛋白在IBV细胞嗜性中的作用4.在nsp4(T53I)和E(E10G)突变的背景下,S2亚基决定IBV细胞嗜性
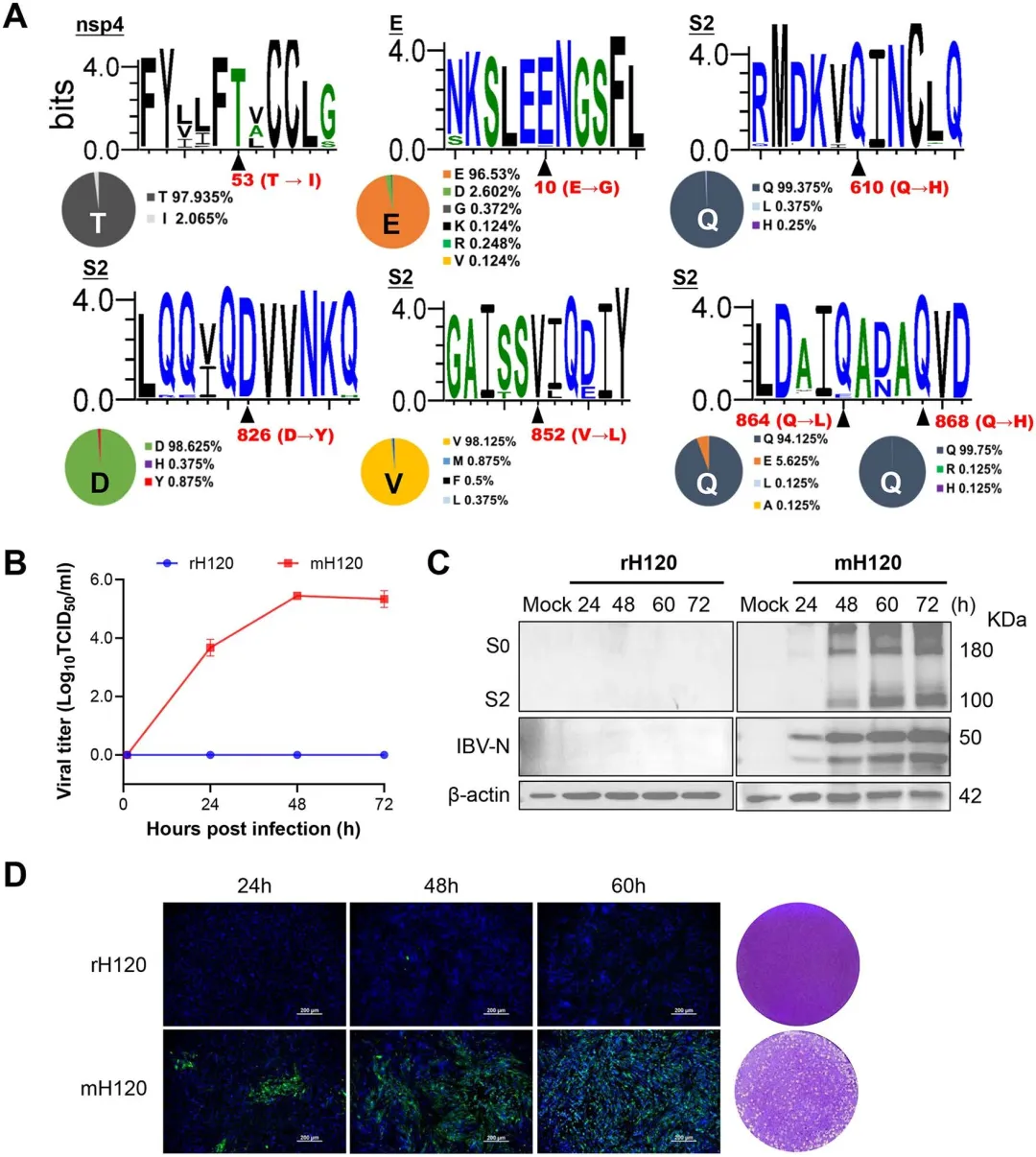
在确定nsp4(T53I)和E(E10G)为必需协同因子后,研究者进一步解析S蛋白内部决定细胞嗜性的功能域。以rSD为骨架保留nsp4(T53I)和E(E10G)突变,分别引入S1亚单位或S2亚单位突变构建重组病毒:rSD-mut/nsp4-E-S2(含5个S2突变:Q610H、D826Y、V852L、Q864L、Q868H)可在BHK-21细胞达到约10^4 TCID50/ml,而rSD-mut/nsp4-E-S1无 detectable 复制,证明S2亚单位而非S1是嗜性决定的关键区域。对S2突变进行时序分析显示,Q610H出现于P0-P10阶段,D826Y和V852L产生于WT-P0阶段,Q864L和Q868H则出现于P0-P10阶段。逐组回复实验表明:单独回复Q610H或同时回复D826Y/V852L均显著降低滴度,而回复Q864L/Q868H完全 abolished 复制能力。进一步组合分析显示,仅保留E+S2或nsp4+S2突变均导致复制显著下降或丧失,nsp4+E+S2(864/868)组合亦无法有效复制。图4.nsp4(T53I)、E(E10G)和S2亚基共同决定IBV细胞嗜性5.最小突变足以赋予GI-1 rH 120菌株哺乳动物细胞嗜性为验证适应性突变的保守性和跨基因型普适性,研究者对NCBI数据库中800余条IBV全基因组序列进行比对分析。结果显示,nsp4(T53)、S2亚单位关键位点(Q610、D826、V852、Q868)及E(E10)在IBV各基因型中高度保守(≥95%序列一致性),仅Q864保守率略低(94.125%)。基于该分析,研究者将nsp4(T53I)、E(E10G)及5个S2突变(Q610H、D826Y、V852L、Q864L、Q868H)这一最小突变组合引入商业化疫苗株H120(GI-1基因型)骨架,构建重组病毒mH120。令人瞩目的是,mH120成功获得BHK-21细胞复制能力,滴度高达10^5.4 TCID50/ml,而野生型rH120完全无法复制。Western blot显示mH120感染细胞在48-60小时达蛋白表达峰值,免疫荧光呈现强阳性信号并形成清晰蚀斑。将mH120在BHK-21细胞连续传代8次后Sanger测序证实所有适应性突变稳定维持,未出现额外替换。图5.适应性突变nsp 4(T53 I)、E(E10 G)和S2亚基对rH 120(GI-1)的影响6.BHK适应增强CEK细胞的复制和细胞-细胞融合,而不影响早期进入为评估BHK-21适应株在原代鸡细胞中的复制特性,研究者比较了rSD与mSD、rH120与mH120在CEK细胞中的生长动力学。结果显示,mSD在CEK细胞中的复制滴度显著高于rSD,而rH120与mH120的峰值水平无显著差异,表明BHK-21适应增强了QX型毒株在天然宿主细胞中的复制效率。为探究S2突变的功能机制,研究者构建仅含S2突变的重组病毒rSD-S2^rP20,通过内化试验检测早期入侵步骤:在CEK和BHK-21细胞中,rSD、rSD-S2^rP20与mSD的病毒基因组内化量无显著差异,表明S2突变不影响病毒吸附与内化。Western blot进一步证实,在CEK和BHK-21细胞中表达野生型S蛋白或S2突变型S蛋白时,S2及S2'切割模式一致,说明S2突变不改变S蛋白切割效率。然而,在CEK细胞中S2突变显著增强了细胞-细胞融合(合胞体形成),而在BHK-21细胞中无论S蛋白质粒转染或mSD感染均未观察到合胞体,提示不同细胞类型存在融合机制差异。蛋白酶抑制剂试验显示,Camostat(TMPRSS家族丝氨酸蛋白酶抑制剂)显著抑制CEK细胞中病毒复制,而E64d(组织蛋白酶抑制剂)显著降低BHK-21细胞滴度,表明IBV在禽源与哺乳动物细胞中利用不同的内吞入侵途径。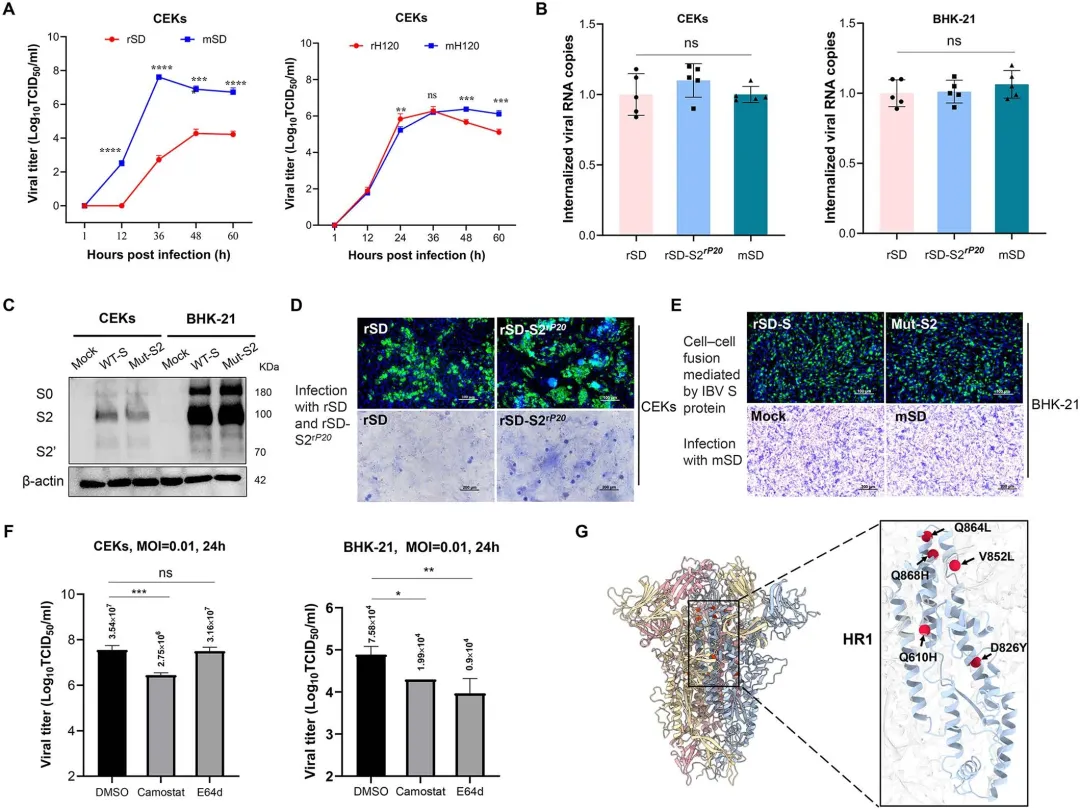 评估哺乳动物细胞适应突变对IBV天然宿主致病性的影响,研究者对1日龄SPF鸡进行攻毒试验并监测14天。野生型rSD株表现出强毒力特征,死亡率高达70%,而mSD突变株仅导致30%死亡且死亡时间延迟;rH120疫苗株及其突变株mH120均未引起死亡。临床症状观察显示,rSD感染鸡在3 dpi即出现严重呼吸道症状,8 dpi达发病高峰;mSD感染鸡仅表现轻微症状,10 dpi才逐渐明显;rH120组有轻度张口呼吸,mH120组基本无症状,所有存活鸡14 dpi均恢复。气管纤毛活性作为敏感感染指标显示,mSD和mH120组纤毛摆动基本保留,而rSD和rH120组出现显著纤毛功能障碍和静止。剖检病变与临床表现一致:rSD组2/6鸡气管积黏液伴出血点、2/6鸡肾脏尿酸盐沉积;mSD组无气管病变、仅1/6鸡轻度肾肿胀;rH120组3/6鸡气管黏液,mH120组无病变。病毒滴定显示rSD在气管组织中显著高于mSD,肾脏无显著差异,脑组织未检测到感染性病毒;rH120组气管滴度显著高于mH120,肾脏和脑组织均为阴性。组织病理学揭示rSD和rH120感染鸡气管上皮严重脱落、基膜裸露,rSD组肾小管伴炎症浸润;而mSD和mH120组气管纤毛完整、肾脏炎症细胞浸润轻微,各组脑组织均无病变。图7.用rSD、rH 120及其突变体感染1日龄SPF鸡
评估哺乳动物细胞适应突变对IBV天然宿主致病性的影响,研究者对1日龄SPF鸡进行攻毒试验并监测14天。野生型rSD株表现出强毒力特征,死亡率高达70%,而mSD突变株仅导致30%死亡且死亡时间延迟;rH120疫苗株及其突变株mH120均未引起死亡。临床症状观察显示,rSD感染鸡在3 dpi即出现严重呼吸道症状,8 dpi达发病高峰;mSD感染鸡仅表现轻微症状,10 dpi才逐渐明显;rH120组有轻度张口呼吸,mH120组基本无症状,所有存活鸡14 dpi均恢复。气管纤毛活性作为敏感感染指标显示,mSD和mH120组纤毛摆动基本保留,而rSD和rH120组出现显著纤毛功能障碍和静止。剖检病变与临床表现一致:rSD组2/6鸡气管积黏液伴出血点、2/6鸡肾脏尿酸盐沉积;mSD组无气管病变、仅1/6鸡轻度肾肿胀;rH120组3/6鸡气管黏液,mH120组无病变。病毒滴定显示rSD在气管组织中显著高于mSD,肾脏无显著差异,脑组织未检测到感染性病毒;rH120组气管滴度显著高于mH120,肾脏和脑组织均为阴性。组织病理学揭示rSD和rH120感染鸡气管上皮严重脱落、基膜裸露,rSD组肾小管伴炎症浸润;而mSD和mH120组气管纤毛完整、肾脏炎症细胞浸润轻微,各组脑组织均无病变。图7.用rSD、rH 120及其突变体感染1日龄SPF鸡8.通过RNA测序(RNA-seq)分析感染野生型和突变型IBV的鸡的差异表达基因(DEG)
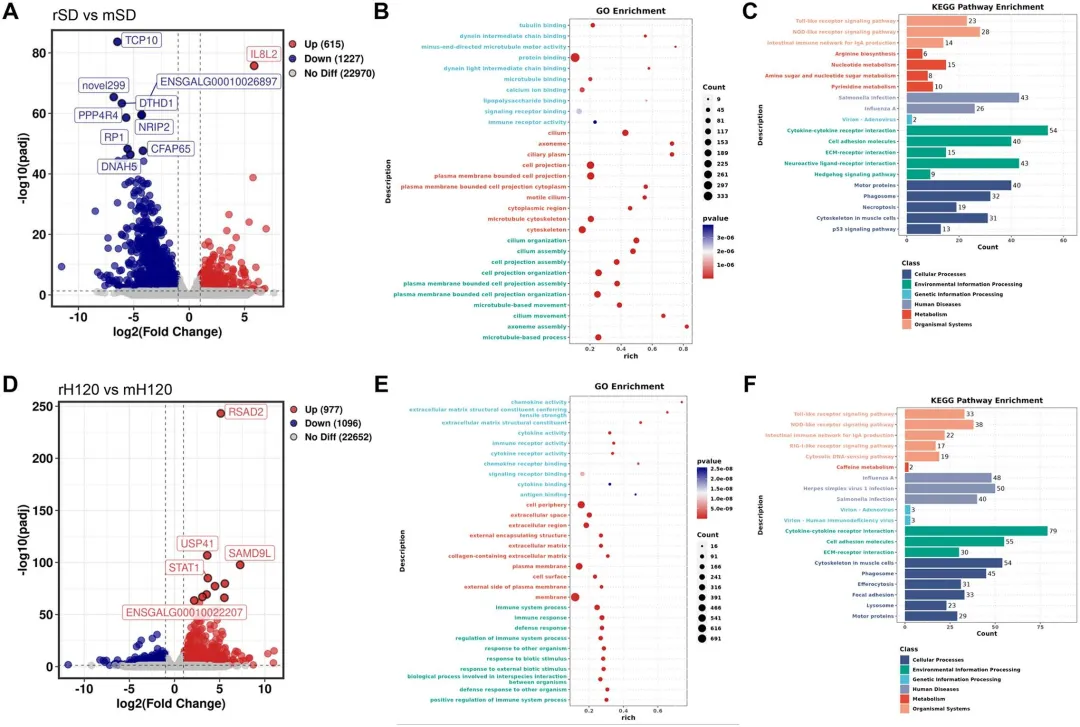
为阐明适应株致病性衰减的分子机制,研究者对感染后5天采集的气管组织进行RNA测序分析。rSD与mSD比较鉴定出1,842个差异表达基因(DEGs),其中rSD感染显著上调促炎细胞因子IL8L2,同时下调凋亡抑制基因TCP10、纤毛组装相关基因PPP4R4、离子稳态调节基因DTHD1及多个未注释基因。GO富集分析显示DEGs主要集中于细胞骨架组织、微管动力学、纤毛结构及纤毛运动等通路,与强毒rSD造成不可逆气管损伤、而减毒mSD保留纤毛完整性的表型高度吻合。rH120与mH120比较获得2,073个DEGs,rH120感染最显著上调抗病毒与先天免疫应答基因如RSAD2和STAT1。GO富集表明DEGs主要关联免疫识别过程,这与rH120作为商业化疫苗株具有强先天免疫激活能力的特性一致。KEGG分析显示rH120/mH120在免疫防御通路方面的差异较rSD/mSD更为显著,同时rSD还激活了多条核苷酸和氨基酸代谢通路,提示QX型野毒株引发更复杂的病理状态和更严重的纤毛损伤。整体转录组数据表明,rSD感染触发较mSD更剧烈的炎症相关通路,而哺乳动物细胞适应的mH120株对抗病毒和先天免疫应答的诱导显著减弱,图8.野生型和突变型IBV毒株感染鸡气管组织的差异表达谱9.用BHK适应性病毒感染可防止WT再攻击
评估BHK-21适应株的免疫保护效力,研究者设计了攻毒保护试验。1日龄SPF雏鸡分别免疫10^3.5 EID50的rSD或mSD,14天后以10^6.0 EID50野生型病毒进行攻毒。免疫后7天血清学检测显示,mSD组IL-1β和IFN-β水平较rSD组呈降低趋势,但无统计学差异。14 dpi时ELISpot检测发现,rSD和mSD均可诱导适度的细胞免疫应答,两组间无显著差异。中和抗体测定表明rSD诱导的中和抗体滴度约为mSD的1.5倍。
攻毒后5天剖检结果显示,未免疫攻毒对照组出现明显气管黏液蓄积和肾脏尿酸盐沉积,而rSD和mSD免疫组均无肉眼可见病变,与未感染对照组一致。病毒滴定显示,两免疫组气管和肾脏病毒载量均显著低于攻毒对照组,rSD与mSD组间无显著差异。最关键的是,rSD和mSD免疫组攻毒后死亡率均为0%,而攻毒对照组死亡率达40%。
本研究首次以当前全球流行的QX型(GI-19)IBV为模型,通过宿主-非宿主细胞交替传代策略成功获得BHK-21细胞高效适应株,系统解析了病毒跨宿主适应的分子机制。研究突破传统认知,证明IBV哺乳动物细胞嗜性扩展并非由S蛋白单独决定,而是需要nsp4(T53I)、E(E10G)与S2亚单位(Q610H、D826Y、V852L、Q864L、Q868H)突变的协同作用;其中S2亚单位而非S1是嗜性决定的关键区域,且5个S2突变具有功能互补性。这一最小突变集在GI-1疫苗株H120骨架中同样有效,证实其跨基因型普适性,为理性设计广适性细胞培养疫苗株提供了核心分子工具。
研究揭示IBV跨宿主适应遵循"预存变异-逐步选择"的进化模式:早期在鸡胚和CEK细胞中积累的nsp4、E及S2突变构成禽源适应背景,为后续BHK-21细胞适应奠定遗传基础;进一步传代中S2亚单位额外突变逐步富集,最终实现哺乳动物细胞高效复制。该发现强调维持准种多样性在病毒适应进化中的关键作用,也为理解冠状病毒跨物种传播的共性规律提供了重要参考。值得注意的是,适应株在哺乳动物细胞中利用组织蛋白酶依赖的内吞途径入侵,与禽源细胞的丝氨酸蛋白酶途径截然不同,这解释了S2突变通过调控HR1构象改变膜融合活性的结构机制。
源论文链接:https://doi.org/10.1371/journal.ppat.1014147
编辑:九九
投稿合作:shouyiyanquan@163.com
可为课题组代发文章宣传、新闻通稿、招聘等
| 标注“原创”仅代表原创编译,原文版权归原作者所有。本文仅用作学术交流分享,如有错误或侵权请后台私信订正或删除。 |


 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
