
请关注我们的姊妹号,更加全面报道植物科学前沿领域研究
近日,云南农业大学园林园艺学院敖平星博士团队在国际权威期刊Food Research International(中科院1区TOP;IF5yeah = 8.5)发表题为“Integrative multi-omics combined with molecular docking and molecular dynamics simulations elucidates the mechanism driving the transition from green notes to fruity aroma during Musang King durian ripening”的研究论文,该研究系统分析了猫山王榴莲在成熟过程中风味的演变,揭示了猫山王榴莲成熟过程中挥发性代谢物的动态重组规律、合成途径及其调控机制,研究成果不仅深化了对榴莲成熟生物学的理解,也为榴莲品种改良提供理论基础。
研究背景
榴莲属锦葵科巨型常绿乔木,树高15~20m,原产于马来西亚、文莱和印尼,目前主产国有泰国、马来西亚、印尼、越南和菲律宾等国家,我国海南、云南、广西、广东和台湾均有种植。我国是世界上最大的榴莲消费国和进口国,2024年和2025年我国分别进口156万吨、180余万吨。
榴莲是典型热带水果,生长条件苛刻,对温湿度异常敏感,果皮坚实密生三角形刺,果肉是由假种皮的肉包组成,风味独特,替代性弱,消费者黏性高,“爱之者赞其香,厌之者怨其臭”,正是这些物质混合在一起形成了榴莲独有的风味。榴莲风味主要由挥发性有机化合物和非挥发性化合物共同决定,成熟过程中,这些化合物的种类和含量会发生复杂变化,进而影响其风味特征。然而,目前对于榴莲成熟过程中风味化合物的变化及分子调控机制仍是未知,本研究以猫山王榴莲为研究对象,综合运用挥发性代谢组学、转录组学、加权基因共表达网络分析、分子对接及分子动力学模拟等技术手段,该研究系统分析了猫山王榴莲在成熟过程中的风味演变,揭示了猫山王榴莲成熟过程中挥发性代谢物的动态重组规律、合成途径及其调控机制。研究成果不仅深化了对榴莲成熟生物学的理解,也为榴莲品种改良提供理论基础。
研究结果
1.猫山王榴莲成熟过程中不同阶段的挥发性代谢物概况分析
研究者利用HS-SPME–GC–MS技术,对猫山王榴莲成熟过程中的4个阶段果肉气味成分进行了系统分析(图1A)。结果发现,PCA分析表明不同阶段的果肉可以明显区分开来,说明榴莲在成熟过程中,风味成分始终处于持续变化之中(图1B)。热图分析也显示,不同阶段具有各自代表性的挥发性代谢物组成,表明榴莲风味在成熟过程中不断重塑(图1C)。
在成熟过程中,检测到420种挥发性代谢物。其中,酯类数量最多,此外还包括酯类、酮类、烃类、酸类、醛类、醇类、杂环化合物和硫类化合物等14类物质,这说明榴莲风味并不是由几种成分决定的,而是由复杂的代谢物体系共同构成,但本研究只解析了挥发性代谢物与风味的关系(图1D)。
研究者发现在猫山王榴莲成熟过程中,成熟早期阶段(S1-S2)挥发性成分以醛类为主,后期酯类和硫类化合物是主要的挥发性物质,其中壬醛、己醛等醛类在成熟初期含量高,赋予果实青草味。随着果实成熟,丁酸乙酯和己酸乙酯等酯类增多,带来浓郁果香;同时异丁基硫醚、二硫化物等硫类挥发物增多,它们参与塑造了榴莲独特而浓烈的硫味,也就是人们常说的“榴莲都很臭,硫类化合物是榴莲臭味的来源”(图1E-k)。此外,研究团队还对猫山王榴莲成熟过程中的4个阶段进行了液相代谢组分析。研究发现,色氨酸、酪氨酸、苯丙氨酸和精氨酸等氨基酸,以及根皮素和柚皮素等糖苷类物质,是猫山王榴莲这个品种独特苦味的主要来源;葡萄糖-6-磷酸则可能与甜味形成密切相关(相关研究内容正在投稿中)。这些挥发性化合物和非挥发性化合物最终形成大家熟悉的榴莲风味。
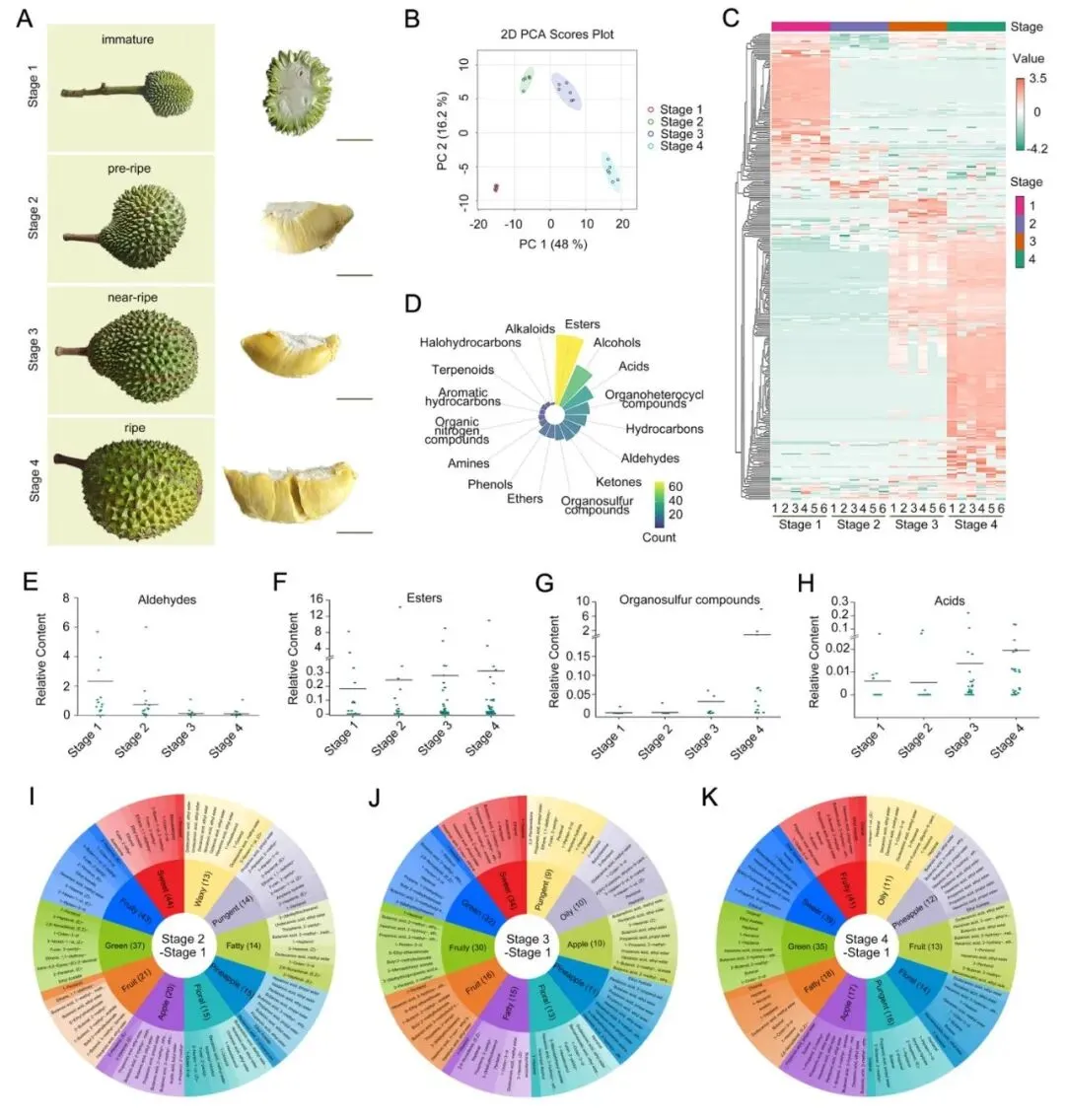
|
图1 猫山王榴莲成熟过程中不同阶段的挥发性代谢物概况分析 |
2.猫山王榴莲成熟过程中不同阶段挥发性物质的动态变化
为进一步弄清榴莲风味在什么时候变化最明显,对不同成熟阶段的挥发性代谢物进行了系统比较研究,共鉴定出394种差异积累挥发性代谢物。从数量变化来看,与未熟期相比,预成熟期、接近熟期和完全成熟期分别有220、60和121种挥发性代谢物显著上调(图2A-C)。这说明榴莲风味变化发生在最关键的两个阶段,一个是从未熟向预成熟过渡的阶段,另一个是从接近成熟到完全成熟的阶段,而中间的阶段则相对稳定。
进一步研究分析还发现,虽然各阶段共享一部分关键核心挥发性代谢物,但完全成熟阶段会出现一些前期没有检测到的特征性成分,其中不少正是赋予榴莲独特气味的含硫化合物。相比之下,未熟期则保留了较多只在早期阶段存在的特异性挥发性醛类代谢物(图2D,E)。
KEGG富集分析显示,不同阶段的差异挥发性代谢物主要涉及氮代谢、丁酸代谢、碳代谢、α-亚麻酸代谢和脂肪酸降解等通路(图2F-H)。其中,α-亚麻酸代谢和脂肪酸降解在各阶段都持续显著变化,提示它们很可能是推动榴莲成熟风味形成的核心代谢通路。
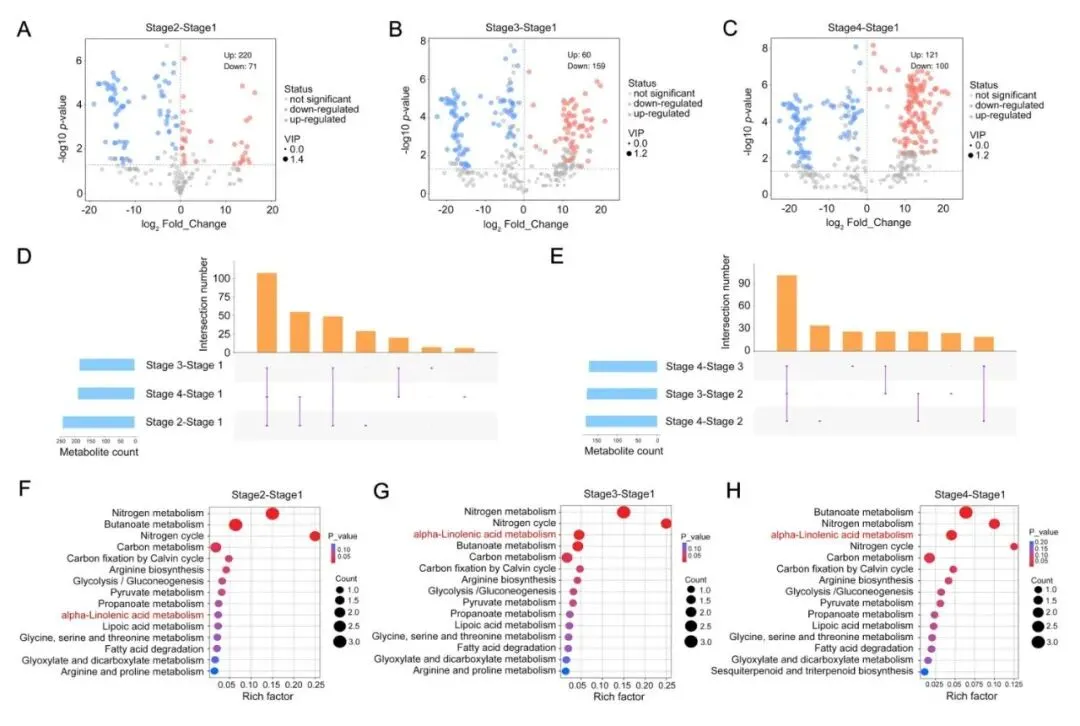
|
图2 猫山王榴莲成熟过程中不同阶段挥发性物质的动态变化 |
3.猫山王榴莲成熟过程中标志性关键风味物质的挖掘
榴莲的风味并不是由某一种物质单独决定的,而是在不同成熟阶段由不同类型的挥发性代谢物共同接力完成。通过聚类分析研究发现,榴莲成熟过程中不同挥发性代谢物呈现出动态变化规律(图3A)。
研究最终挖掘了4种关键风味标志物,即(E,Z)-2,6-壬二烯醛、(Z)-3-己烯醛、丁酸乙酯和己酸乙酯。其中,前两者主要在早期赋予榴莲青草风味,后两者主要在中后期赋予榴莲显著的果香和甜香。此外,2-羟乙基异丁基硫醚、甲基乙基二硫化物和二乙基二硫化物等含硫挥发性物质,也共同参与塑造榴莲独特而浓烈的硫味(图3B,C)。
相关性分析显示,果香酯类之间他们相互协同,醇类与其对应的酯类之间也高度相关。相比之下,早期醛类与晚期酯类、硫化物之间多呈负相关,尤其是二乙基二硫化物、甲基乙基二硫化物与丁酸乙酯、己酸乙酯之间存在显著正相关(图3D)。
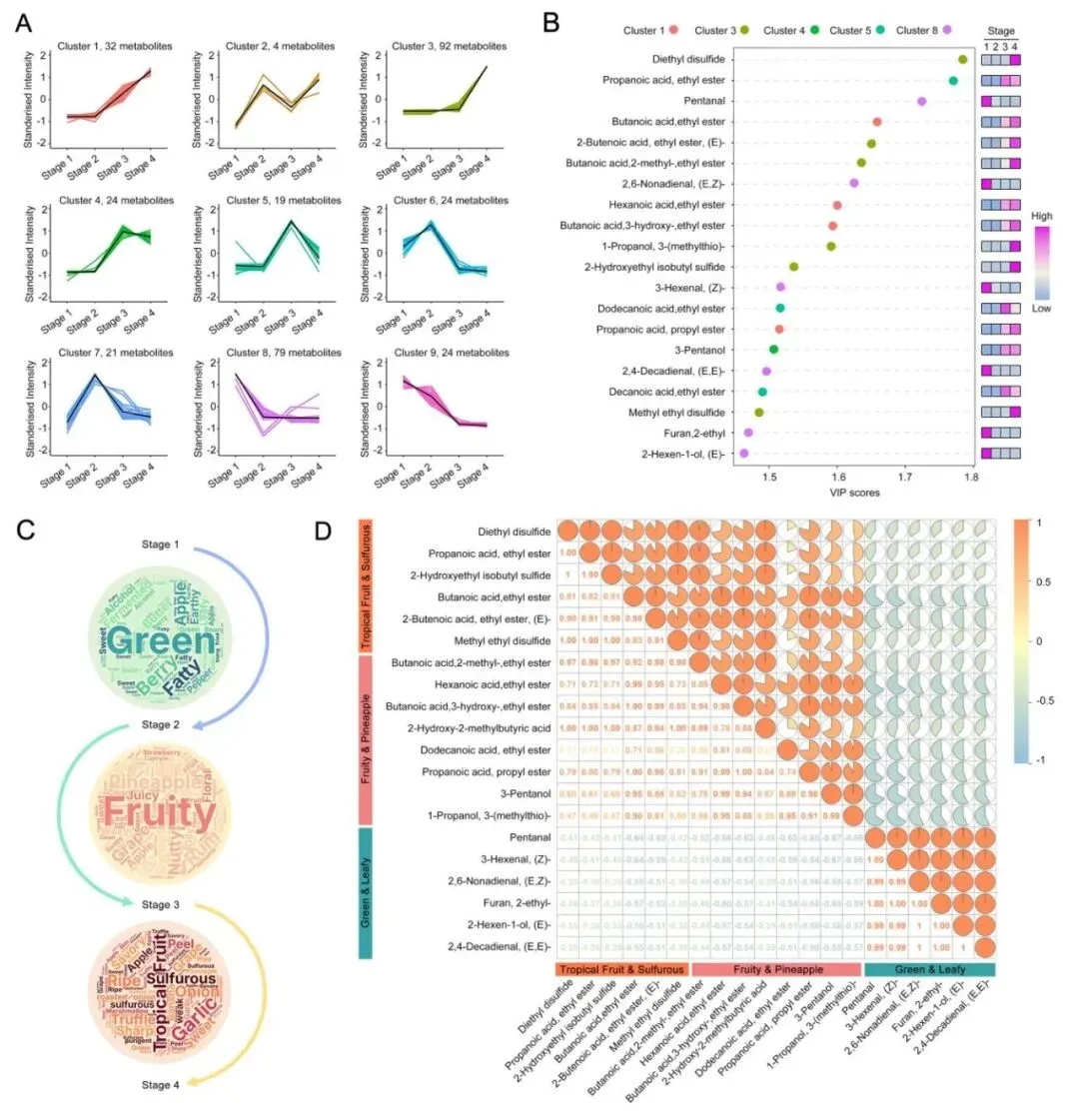
|
图3猫山王榴莲成熟过程中风味物质分析 |
4.猫山王榴莲成熟过程中基因表达分析
风味变化并非孤立发生的表象,其背后是基因表达程序的系统性重构。研究团队对榴莲果实四个发育阶段的果肉进行全转录组测序,共鉴定出33,696个表达基因。
PCA分析显示,不同阶段在基因表达不一样,表明榴莲在成熟过程中,不仅风味成分在持续变化,调控这些变化的基因程序也在同步切换(图4A)。
与未成熟期相比,预成熟期、近成熟期和完全成熟期分别检测到14,705、17,194和17,996个差异表达基因,差异基因的数量随成熟进程逐步扩大(图4B,C)。聚类分析进一步揭示,一些基因在早期高表达,随后表达量逐渐下降,另一些基因则在后期,尤其是近熟期到完全成熟期显著上调,这些后期上调的基因很多属于α-亚麻酸代谢、脂肪酸降解等与风味形成密切相关的代谢通路(图4D,E)。
GSEA分析进一步支持了这一结论:脂肪酸降解通路在完全成熟期显著增强,α-亚麻酸代谢也表现出明显的阶段性变化(图4F-H)。综合来看,榴莲成熟过程中的风味变化并非随机发生,而是由一套有序切换的基因表达程序精确调控的结果。
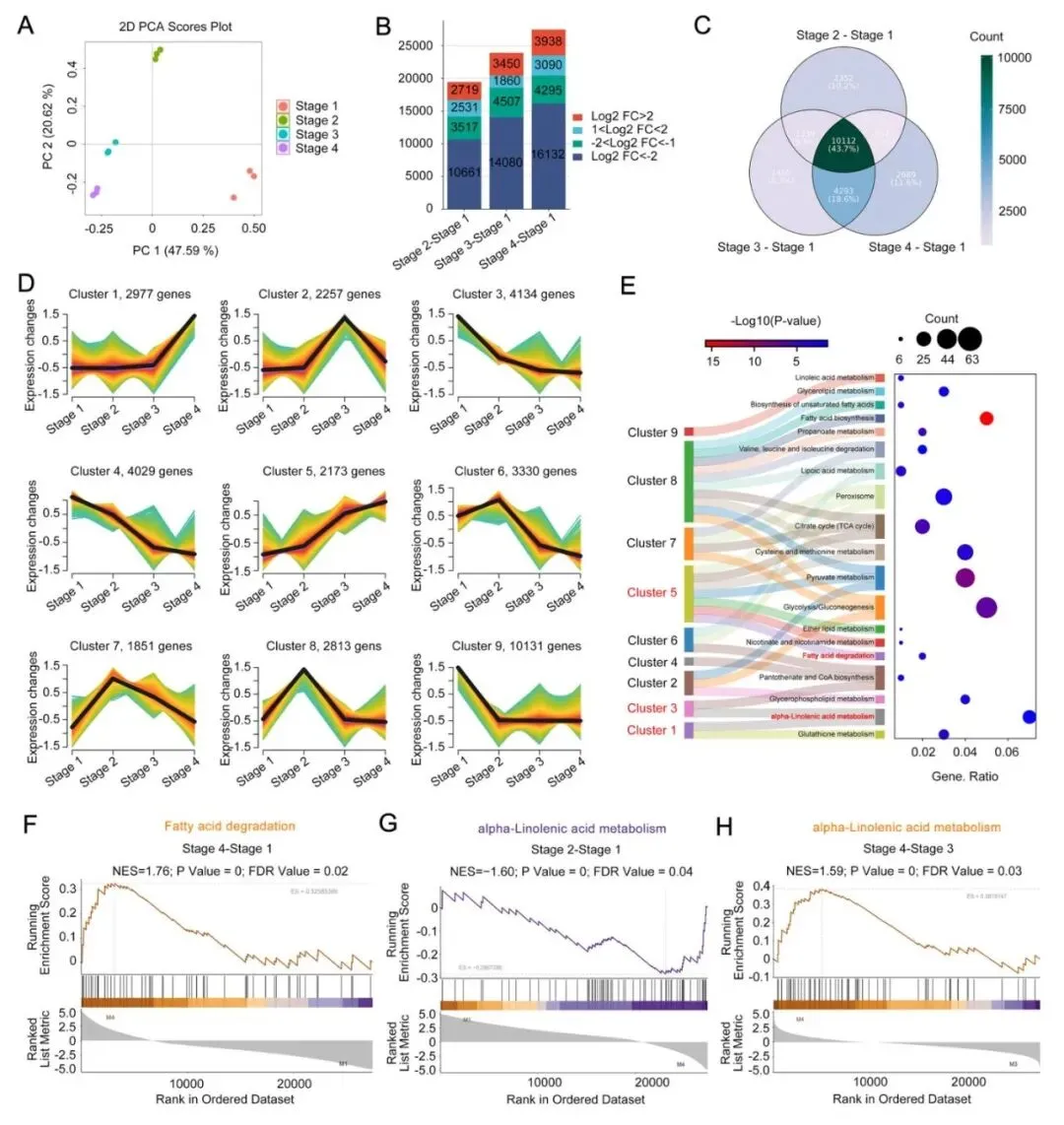
|
图4 猫山王榴莲成熟过程中基因表达分析 |
5.猫山王榴莲成熟过程中脂肪酸衍生挥发性代谢物的合成分析
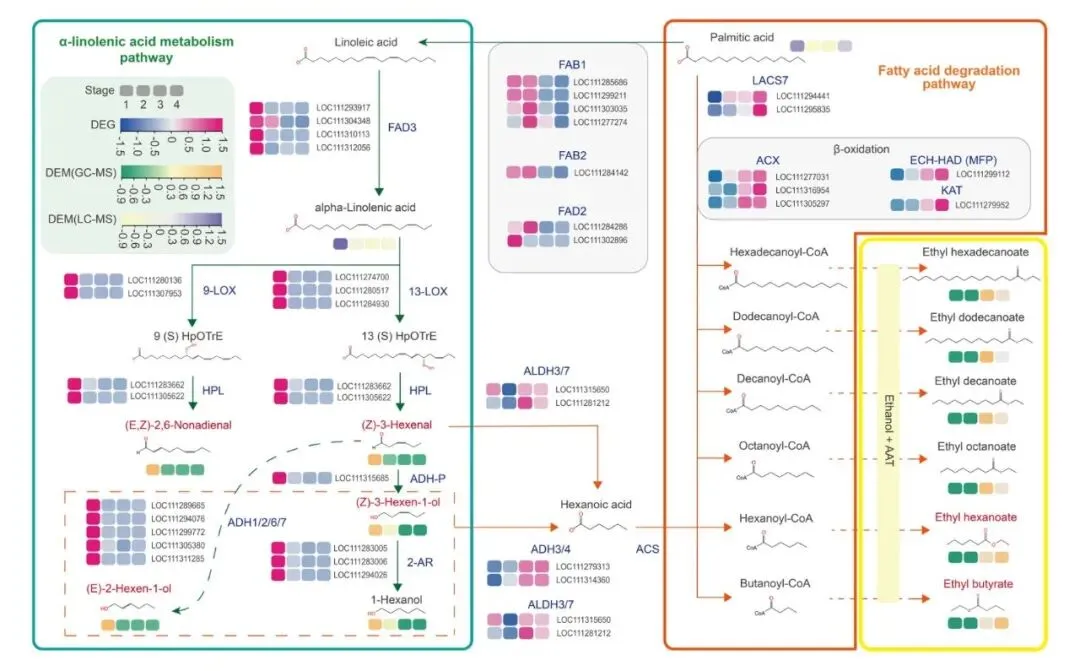
在整合代谢组和转录组数据后,研究者进一步重建了榴莲成熟过程中脂肪酸衍生挥发性代谢物的形成路径,解析了青草味如何一步步转变成果香味(图5)。
在α-亚麻酸代谢模块中,亚油酸经FAD3转化为α-亚麻酸后,可通过9-LOX/13-LOX及HPL等酶促途径,生成一系列C6/C9醛类物质,包括(E,Z)-2,6-壬二烯醛与(Z)-3-己烯醛。这些醛类正是成熟早期青草气味的主要来源。随后,这些醛类可进一步还原为相应的醇类,或继续氧化为酸类,为成熟后期酯类合成提供前体。
另一方面,在脂肪酸降解模块中,棕榈酸等脂肪酸经过一系列关键酶的作用,生成丁酰辅酶A、己酰辅酶A等中间产物,进而在AAT等酶的催化下与乙醇结合,最终形成丁酸乙酯、己酸乙酯、辛酸乙酯等典型果香酯类物质。
从基因表达图谱来看,调控早期青草气味形成的LOX–HPL模块及相关ADH基因,在成熟早期表达水平较高;而与后期果香形成密切相关的β-氧化基因及酯合成基因,则在成熟后期显著增强。
|
图5 脂肪酸衍生挥发性代谢物的合成路径 |
6.榴莲风味形成相关基因模块及核心调控网络分析
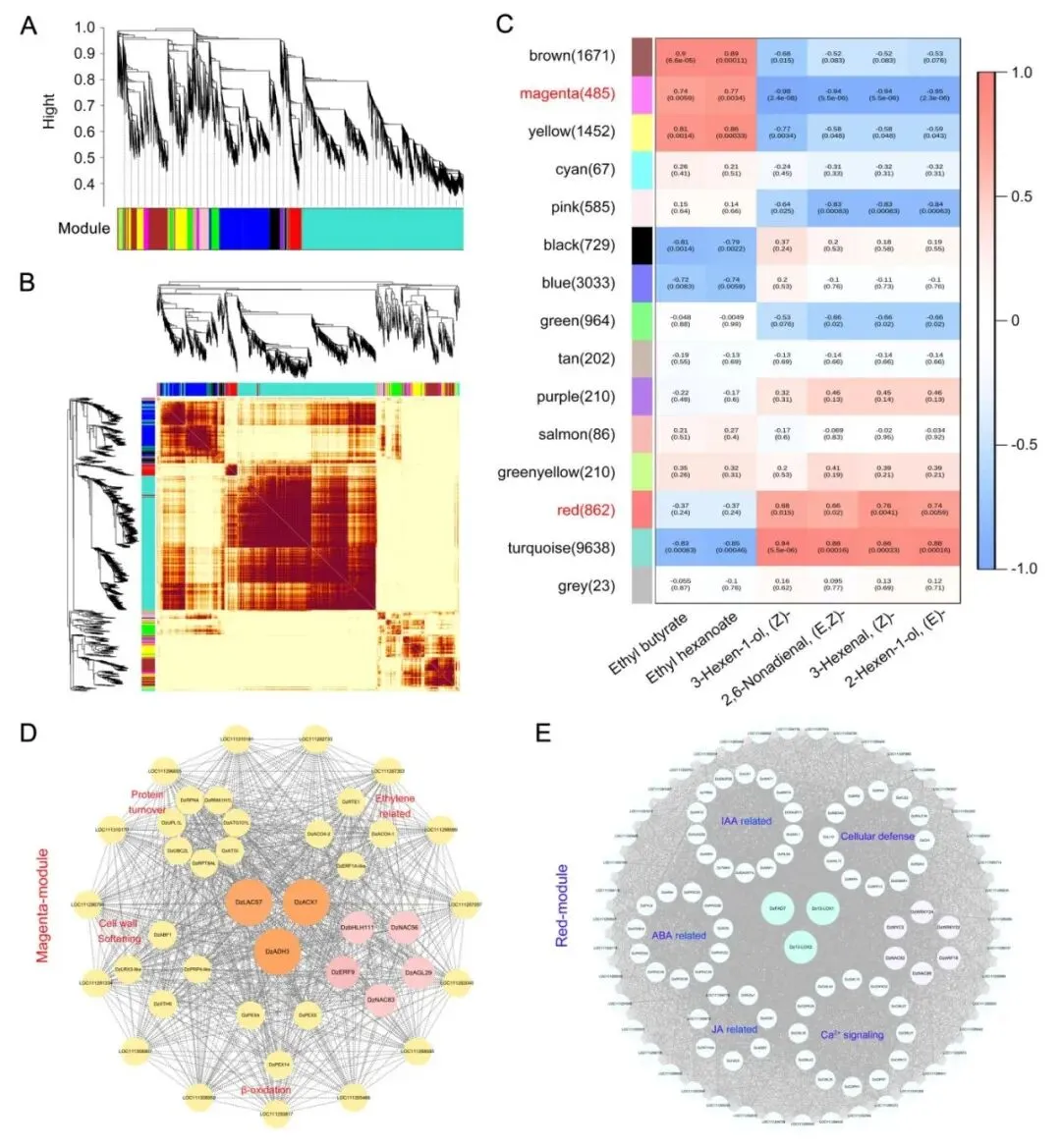
为了进一步解析驱动榴莲风味形成的核心调控网络,研究者开展了加权基因共表达网络分析(WGCNA),将成熟过程中的表达基因划分为15个共表达模块,并结合代谢物数据,分析各模块与风味性状之间的关联(图6A,B)。结果发现红色模块与醛类挥发性代谢物呈显著正相关,而品红色模块则与脂肪酸衍生酯类显著相关。这表明不同模块分别对应榴莲成熟早期的青草气味形成与成熟后期的果香积累过程(图6C)。
进一步分析各模块中的核心基因发现,品红色模块中富集了多个与过氧化物酶体及脂质代谢相关的重要因子,尤其是ACX1和LACS7,它们与β-氧化、果肉软化及成熟后期的果香形成密切相关。同时,乙烯合成与信号转导相关基因也大量聚集于该模块(图6D)。相比之下,红色模块中则富集了13-LOX等脂质氧化相关基因,以及生长素、茉莉酸和脱落酸信号通路的相关组分,与其在早期醛类形成中的功能高度一致(图6E)。 以上结果说明,榴莲风味的形成并非由单一结构基因独立驱动,而是多个基因模块协同配合的结果。
|
图6 榴莲风味形成相关基因模块及核心调控网络 |
7.生长素和乙烯调控榴莲风味合成关键基因分析
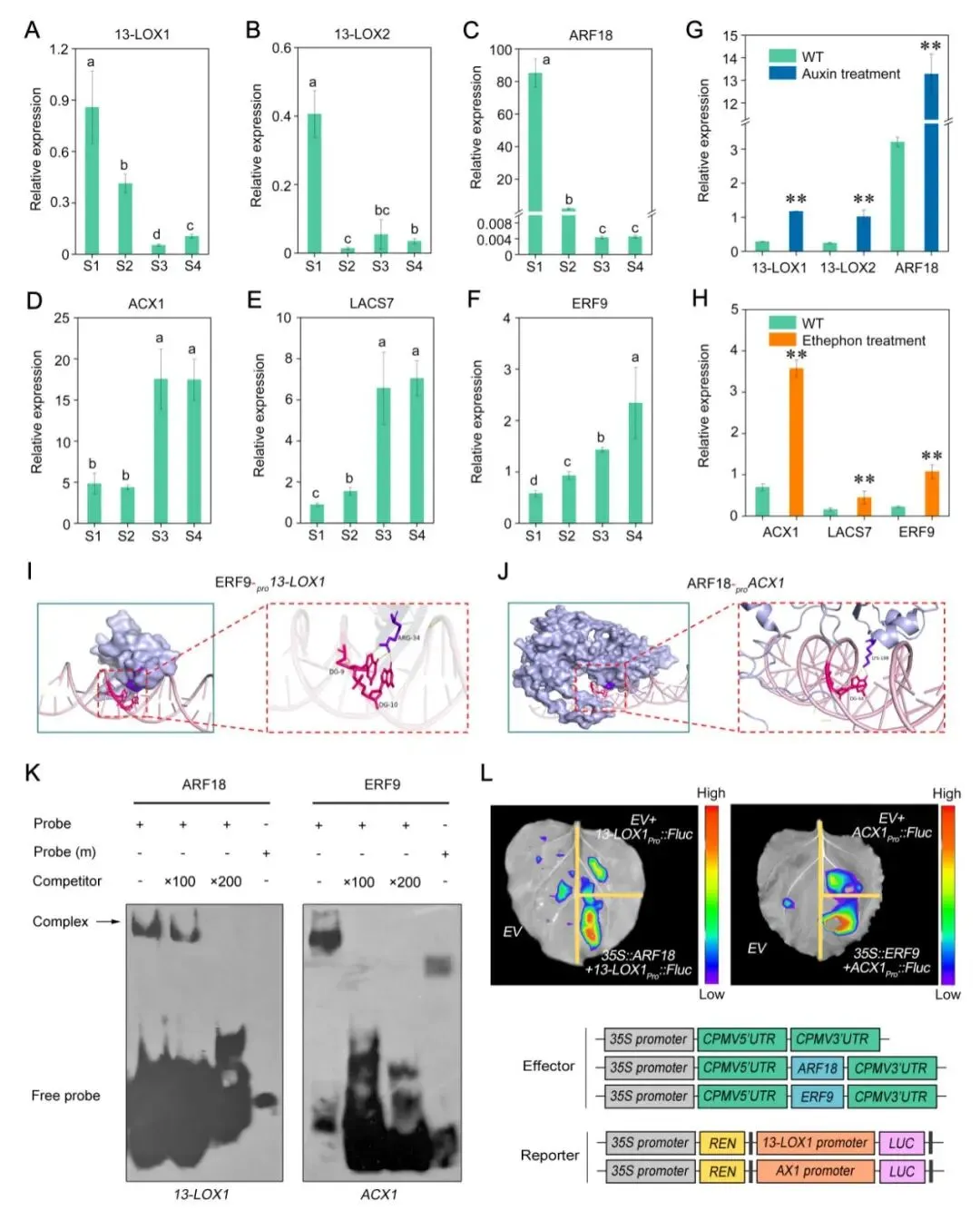
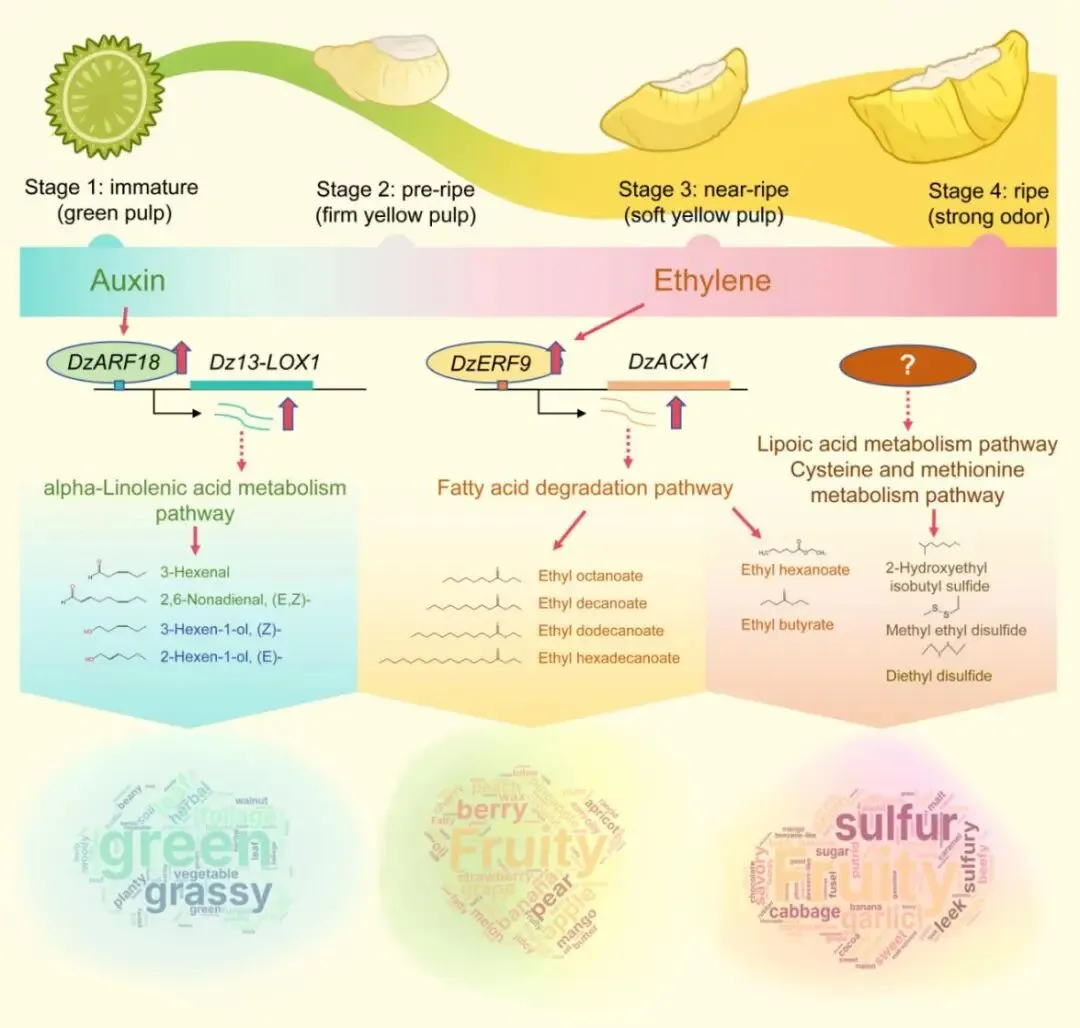
为验证关键风味途径基因的调控机制,研究者选取了Dz13-LOX1、Dz13-LOX2、DzACX1、DzLACS7、DzARF18和DzERF9进行RT-qPCR验证。结果显示,Dz13-LOX1、Dz13-LOX2和DzARF18在成熟过程中总体呈下降趋势,而DzACX1、DzLACS7和DzERF9则随成熟进程逐渐升高,与转录组数据一致(图7A-F)。激素处理实验进一步表明,一定浓度的生长素可显著诱导Dz13-LOX1、Dz13-LOX2和DzARF18的表达,而1 mM乙烯利则显著促进DzACX1、DzLACS7和DzERF9的转录积累,说明生长素和乙烯分别参与调控青草气味与果香酯类形成相关通路(图7G,H)。
|
图7 生长素和乙烯调控榴莲风味合成关键基因分析 |
蛋白-DNA分子对接、EMSA及双荧光素酶报告实验共同提供了直接证据:DzARF18能够结合并转录激活Dz13-LOX1启动子,DzERF9则能够结合并激活DzACX1启动子(图7K,L)。这表明,榴莲在成熟过程中至少存在两条重要的调控通路,一条偏向早期青草气味的形成,另一条偏向后期果香的积累,二者协同推动风味由青草气向成熟果香转换。
8. 榴莲关键气味分子与人类嗅觉受体互作的分子模拟分析
除了研究榴莲风味是怎样形成的,团队还进一步研究了这些关键挥发性代谢物如何被人的嗅觉系统识别。
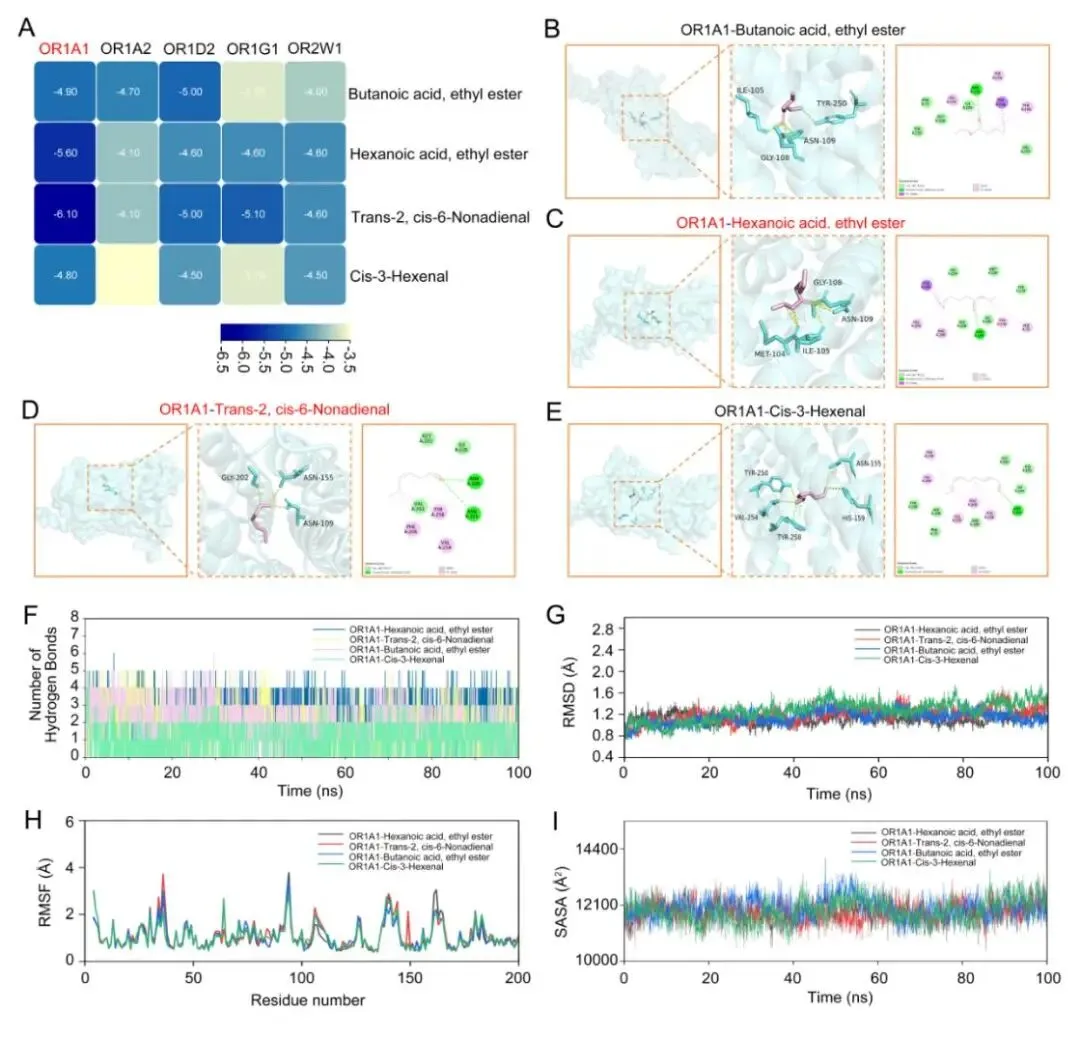
研究者选取了4种代表性榴莲风味分子,并将它们与5个嗅觉受体进行分子对接分析。结果显示,OR1A1对这些配体表现出相对更高的结合亲和力,其中trans-2,cis-6-壬二烯醛与OR1A1的结合最强,其次是己酸乙酯,而丁酸乙酯和cis-3-己烯醛也表现出较稳定的结合潜力(图8A)。
进一步的结合位点分析表明,这些醛类和酯类分子可以进入OR1A1的核心结合口袋,并与其中的极性和疏水残基形成相互作用(图8B-E)。100 ns分子动力学模拟结果也显示,这些复合物在模拟过程中整体保持稳定,说明它们与受体之间具有较好的结合稳定性(图8F-I)。
|
图8 榴莲关键气味分子与人类嗅觉受体互作的分子模拟分析 |
这一结果不仅解释了榴莲风味是怎样形成的,也进一步回答了这些关键气味分子为什么能够被人体有效感知,为理解榴莲标志性风味的感知机制提供了新的理论支持。
研究结论
本研究较系统地揭示了猫山王榴莲成熟过程中风味形成的物质基础和分子调控机制,回答了榴莲风味从何而来、如何形成以及如何随成熟进程动态变化等问题。
研究表明,榴莲风味在成熟过程中会持续发生重塑。早期以青草气息为主,中后期逐渐转向果香和甜香,到了完全成熟阶段,则进一步形成典型而浓郁的果香与硫香复合风味。这一过程并非由某一种成分单独决定,而是多类挥发性物质协同积累、多个代谢通路连续衔接以及关键激素信号精准调控共同作用的结果。
这项研究不仅加深了人们对榴莲气味形成规律的认识,也为今后提升榴莲果实品质、优化采后管理以及选育优质品种提供了理论依据。
|
图9 榴莲成熟过程中风味由青草味向果香和硫味转变的调控模型 |
云南农业大学园林园艺学院教师敖平星博士为论文通讯作者,课题组在读研究生李中艳为第一作者,闫天赐博士及许彬老师为共同第一作者。该研究工作得到热带特优果树种质资源利用专项(No. A3012024043001)、云南农业大学榴莲种质资源圃建设项目(No. A2032025197)及云南农业大学省部共建云南生物资源保护与利用国家重点实验室开放课题(No. KX14147300003)资助。
论文链接:
https://doi.org/10.1016/j.foodres.2026.119168

植物科学最前沿,专注于植物科学前沿进展、资讯、招聘信息的发布及方法软件共享等。投稿及招聘请后台回复“投稿”,均为无偿;商务合作请联系微信ID:zwkxqy ;

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
