J Virol.|安徽农业大学孙裴教授团队最新研究:HSPA5通过PERK-eIF2α通路诱导自噬靶向VP2,抑制塞内卡病毒SVA复制
2026年4月13日,《Journal of Virology》在线发表了题为“HSPA5 induces autophagy targeting VP2 through the PERK-eIF2α signaling pathway to inhibit SVA replication”的研究论文。安徽农业大学动物医学院团队系统阐明了热休克蛋白HSPA5在塞内卡病毒(Senecavirus A,SVA)复制过程中的抗病毒作用。该研究首次证明,HSPA5可通过激活未折叠蛋白反应(UPR)的PERK-eIF2α分支诱导自噬,特异性降解病毒结构蛋白VP2,从而显著抑制SVA在宿主细胞中的复制。这一发现为理解SVA与宿主相互作用提供了新机制,也为当前缺乏商品化疫苗的猪病防控研究提供了重要科学依据。
SVA为何成为猪业威胁?病毒结构与当前防控困境
塞内卡病毒A(SVA),又称塞内卡谷病毒(SVV),是一种单股正链RNA病毒,属于小RNA病毒科塞内卡病毒属。其基因组全长约7.2 kb,含有一个大开放阅读框,编码一个多聚蛋白,经蛋白酶切割后产生L蛋白、4种结构蛋白(VP1、VP2、VP3、VP4)和7种非结构蛋白(2A、2B、2C、3A、3B、3C、3D)。其中VP2蛋白位于病毒衣壳表面,含有多个抗原表位,不仅参与宿主细胞受体识别,还是诱导中和抗体的关键靶标,已被视为SVA疫苗开发的优先候选。
近年来,SVA在巴西、中国、美国等主要养猪国家快速传播,导致新生仔猪急性死亡、母猪繁殖障碍及猪群水疱性病变,给全球猪业造成重大经济损失。目前,尚无针对SVA的商品化疫苗,防控主要依赖严格的生物安全措施。病毒复制的分子机制仍不完全清楚,宿主细胞如何天然限制SVA复制,成为研究热点。热休克蛋白(HSPs)家族作为细胞内重要的分子伴侣,广泛参与蛋白质折叠、运输和降解,在多种病毒感染中发挥调控作用。安徽农业大学研究团队以此为切入点,系统筛选HSPs家族成员对SVA增殖的影响。
筛选HSPs家族:HSPA5特异性抑制SVA复制
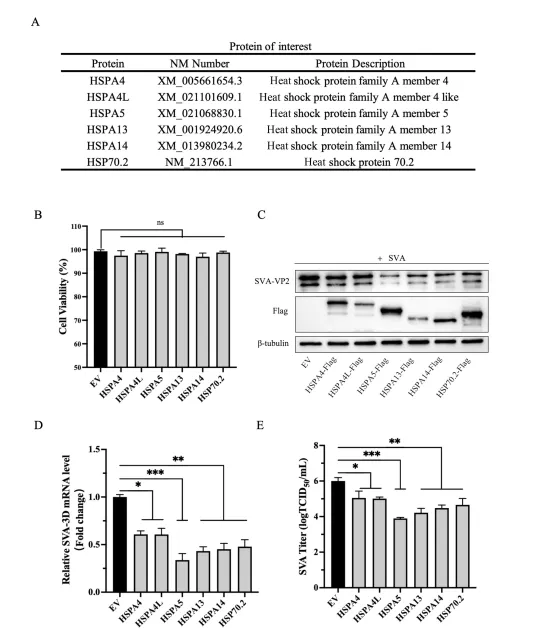
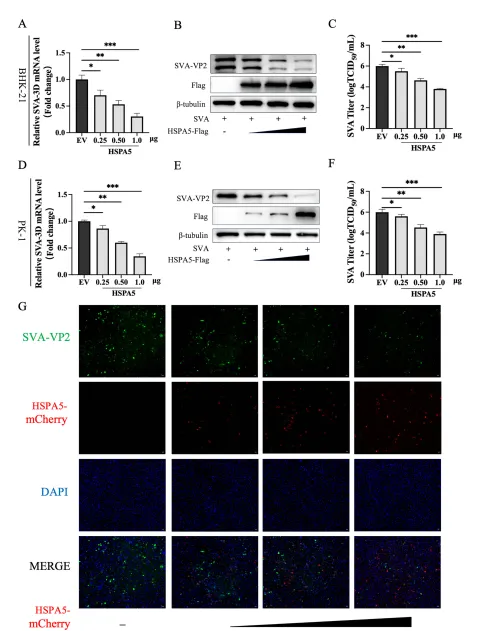
研究团队选取了猪源HSP家族多个代表性成员(HSPA4、HSPA4L、HSPA5、HSPA13、HSPA14、HSP70.2等),构建表达质粒并转染BHK-21细胞。细胞活力检测显示,各转染组细胞状态一致。随后以0.01 MOI SVA感染细胞,16小时后收集样本。通过Western blot、RT-qPCR和TCID₅₀测定病毒复制水平,结果显示:仅HSPA5过表达组显著降低SVA VP2蛋白表达、病毒RNA拷贝数及病毒滴度,而其他HSP成员影响不明显或无显著作用。这一抑制效应在PK-1猪肾细胞中同样成立,且呈明显的剂量依赖性:随着HSPA5表达质粒转染量增加,病毒复制指标逐步下降。间接免疫荧光(IFA)也观察到VP2荧光强度随HSPA5表达增强而减弱。
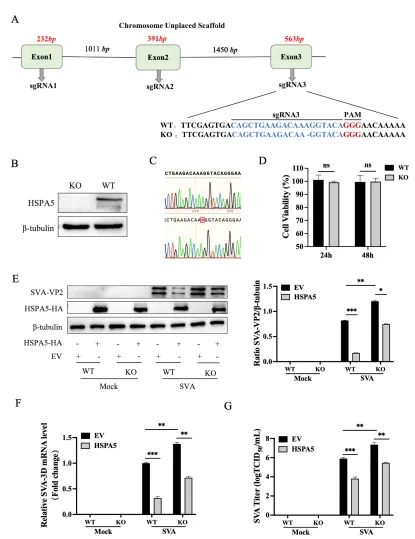
为进一步验证HSPA5的必要性,团队利用CRISPR-Cas9技术构建了HSPA5基因敲除的BHK-21稳定细胞系(BHK-HSPA5-KO)。Western blot和Sanger测序确认敲除成功,细胞活力检测显示敲除不影响细胞增殖。
SVA感染实验显示,与野生型BHK-21细胞相比,敲除细胞中VP2蛋白、3D基因mRNA及病毒滴度均显著升高。转染HSPA5-HA质粒回补后,病毒复制水平恢复至对照组。这些“过表达抑制、敲除促进”的结果,明确HSPA5是限制SVA复制的关键宿主因子。
HSPA5作用于病毒复制阶段,而非吸附或进入
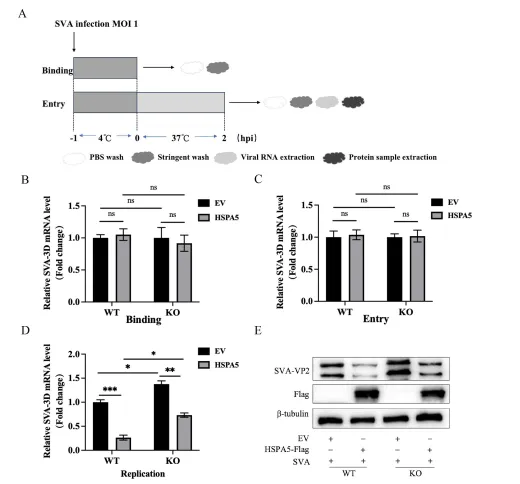
病毒生命周期包括吸附、进入、复制、组装与释放等环节。研究团队采用吸附-进入实验设计(4℃允许结合、37℃允许内化),通过RT-qPCR检测病毒3D mRNA水平,发现HSPA5过表达或敲除均不影响SVA的吸附和进入过程。随后聚焦复制阶段,结果显示:在HSPA5过表达细胞中,8小时感染后病毒RNA合成显著受抑;敲除细胞则RNA水平升高。Western blot检测VP2蛋白也呈现一致趋势。这一发现表明,HSPA5主要在病毒复制阶段发挥抑制作用,而非阻断病毒早期入侵。
HSPA5与VP2直接结合,通过自噬-溶酶体途径实现降解
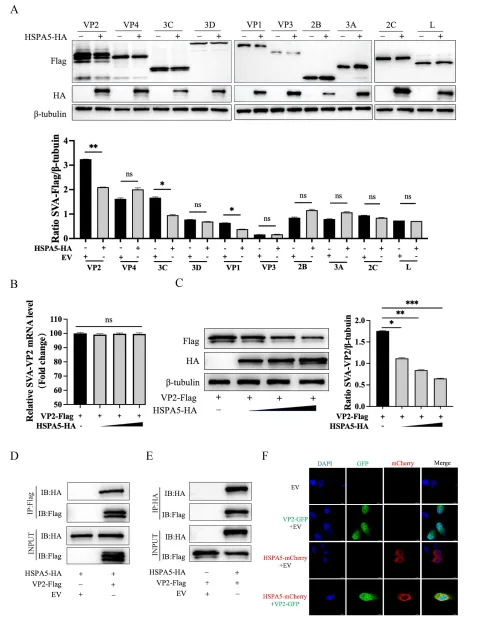
为解析HSPA5的分子作用靶标,团队将HSPA5-HA质粒与SVA各编码蛋白(VP1、VP2、VP3、VP4、L、2B、2C、3A、3C、3D)的Flag标签质粒共转染HEK-293T细胞。Western blot筛查显示,HSPA5可降解VP1、VP2和3C蛋白,其中对VP2的降解作用最为显著,且呈剂量依赖性,同时不影响VP2 mRNA转录水平。免疫共沉淀(Co-IP)实验证实HSPA5与VP2存在物理相互作用;激光共聚焦显微镜观察到两者在细胞质中共定位。SVA实际感染条件下,过表达HSPA5仍能与VP2结合并共定位,进一步验证了这一相互作用的生理相关性。
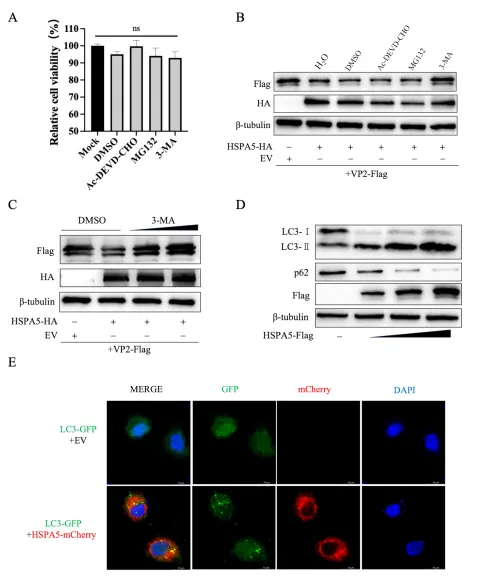
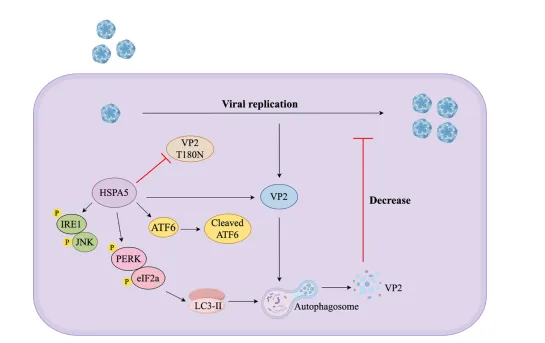
细胞内蛋白降解主要通过凋亡、自噬-溶酶体和泛素-蛋白酶体三条途径。研究团队分别使用凋亡抑制剂Ac-DEVD-CHO、自噬抑制剂3-甲基腺嘌呤(3-MA)和蛋白酶体抑制剂MG132处理共转染细胞。细胞活力检测确认各抑制剂无明显毒性。结果显示,仅3-MA能剂量依赖性地恢复VP2蛋白水平,提示HSPA5通过自噬-溶酶体途径降解VP2。进一步实验发现,HSPA5过表达显著促进LC3-I向LC3-II转化、降低自噬底物p62水平,并增加LC3-GFP斑点形成,证实HSPA5可诱导自噬发生。
PERK-eIF2α通路是HSPA5诱导自噬的关键开关
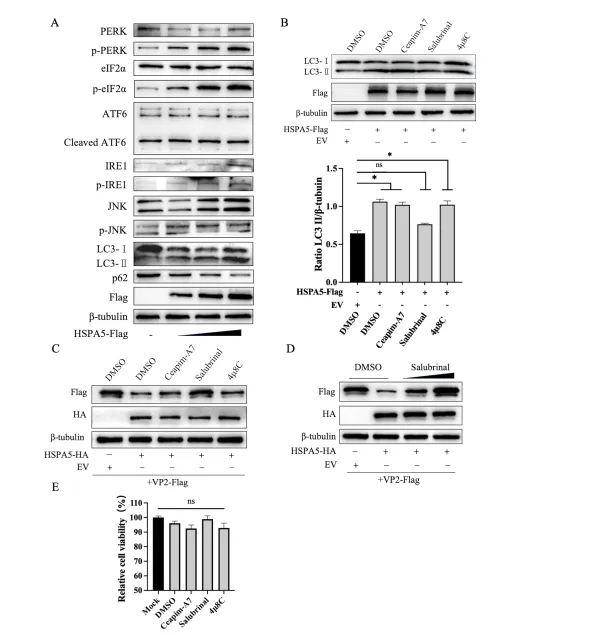
HSPA5作为内质网分子伴侣,是未折叠蛋白反应(UPR)的核心调控因子。UPR包括IRE1、PERK和ATF6三条分支。Western blot分析显示,HSPA5过表达剂量依赖性地激活PERK磷酸化及其下游eIF2α磷酸化,同时IRE1、JNK和ATF6分支也有一定激活,但PERK通路最为显著。使用PERK特异性抑制剂Salubrinal、IRE1抑制剂4μ8C和ATF6抑制剂Ceapim-A7处理后,仅Salubrinal能有效阻断HSPA5诱导的LC3-II增加,并恢复VP2蛋白水平,且呈剂量依赖性。细胞活力检测排除抑制剂毒性影响。这些结果明确,HSPA5通过激活UPR的PERK-eIF2α通路诱导自噬,进而介导VP2降解。
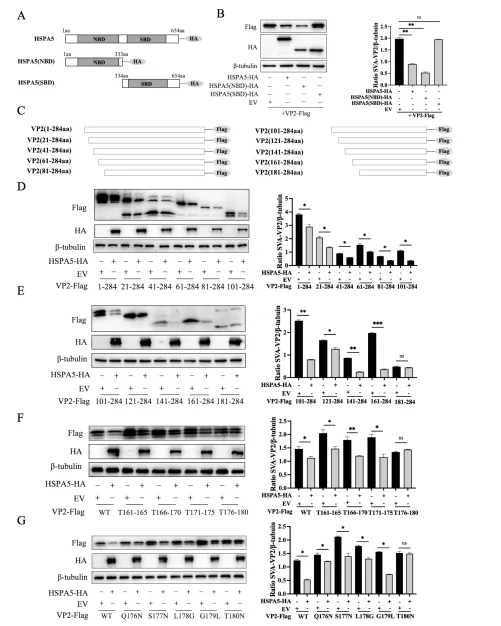
NBD结构域与VP2 Thr180位点是功能关键
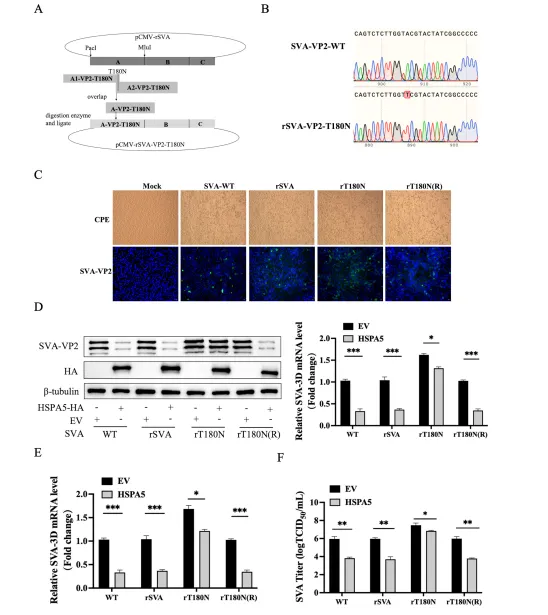
HSPA5包含N端核苷酸结合域(NBD,aa 1-333)和C端底物结合域(SBD,aa 334-652)。构建截短体实验显示,仅NBD片段可有效降解VP2,SBD则无此功能,表明NBD是执行降解功能的关键结构域。反向针对VP2,研究团队构建了一系列20个氨基酸步进的截短体及丙氨酸扫描突变体,最终将关键位点定位于VP2第176-180氨基酸区域,进一步通过点突变鉴定出Thr180为HSPA5介导降解的关键残基。T180N突变完全恢复VP2蛋白水平,但不影响与HSPA5的结合,说明该位点特异性参与降解过程而非结合。
T180N突变病毒对HSPA5产生抵抗,证实位点功能
为验证体内意义,团队利用反向遗传学技术,以pCMV-rSVA为骨架构建携带VP2 T180N突变的感染性克隆,并拯救出rT180N病毒及修复病毒rT180N(R)。免疫荧光和测序确认病毒成功拯救。在HSPA5过表达的BHK-21细胞中,野生型SVA、rSVA及rT180N(R)复制均被显著抑制,而rT180N病毒的VP2蛋白、RNA水平及滴度抑制效果明显减弱,表明T180N突变赋予病毒对HSPA5抗病毒活性的抵抗能力。
研究意义与展望:为SVA防控提供新分子靶标
本研究系统揭示了HSPA5作为抗病毒因子的作用机制:通过PERK-eIF2α信号通路诱导自噬,特异性靶向并降解SVA VP2蛋白,从而抑制病毒复制。NBD结构域的必需性及VP2 Thr180位点的关键作用。该发现不仅拓展了HSPA5在病毒-宿主互作中的抗病毒谱(此前已在HBV、ZIKV等病毒中观察到类似调控),也深化了对SVA复制调控的认识。SVA目前仍缺乏有效疫苗,本研究为开发基于HSPA5通路的抗病毒策略或增强宿主天然免疫的防控手段提供了重要线索。未来,可进一步在原代猪细胞和活体模型中验证机制安全性,并探索模拟NBD功能的小分子或基因编辑育种等应用路径。这一成果为猪病基础研究与产业应用架起了桥梁,有助于推动SVA防控技术的进步。
参考文献:https://doi.org/10.1128/jvi.02103-25注:文中插图源于Journal of Virology,欢迎关注交流。
本文为原创编译,仅代表对原文内容的整理与解读,原文版权归原作者及出版方所有。本文仅用于学术交流与分享,旨在促进研究与学习。如内容存在错误或涉及侵权,请通过后台私信联系我们进行更正或删除。(当前本公众号的原创文章均为对原文的整理与解读,原文版权归原作者及出版方所有)