合作文章丨华南农业大学唐湘如团队揭示了纳米氧化锌对水稻镉胁迫的缓解作用
- 2026-06-21 10:58:20

英文题目:ZnO nanoparticle-based seed priming modulates early growth and enhances physio-biochemical and metabolic profles of fragrant rice against cadmium toxicity
中文题目:纳米氧化锌诱导水稻早期生长及增强水稻抗镉胁迫的生理生化和代谢特征
期刊名称:Journal of Nanobiotechnology
影响因子:10.6
作者单位:华南农业大学
普奈斯提供服务:透射电镜(TEM)检测
DOI号:https://doi.org/10.1186/s12951-021-00820-9
前言
本研究在正常和Cd胁迫条件下(0和100mg·L-1)应用四种水平的ZnO NPs(0、25、50和100mg·L-1)于两个香米品种(象牙香占和玉香油占)。ZnO NPs对种子萌发无显著影响(p>0.05),但显著提高了Cd胁迫下幼苗的生长及相关生理指标,施用ZnO NPs后,地上部和全苗的平均鲜重分别增加了16.92-27.88%和16.92-27.88%,也显著提高了根鲜重、根冠长。此外,ZnO NPs的应用诱导了生理和生化属性的调节,主要表现在低浓度的ZnO NPs处理提高了根和地上部的超氧化物歧化酶(SOD)活性、根中的过氧化物酶(POD)活性和金属硫蛋白含量。在Cd胁迫下,施用ZnO NPs提高α-淀粉酶和总淀粉酶的活性。此外,还检测到了幼苗中Zn浓度和ZnO NPs吸收的调节作用。代谢组学分析表明,丙氨酸、天冬氨酸和谷氨酸代谢、苯丙烷类化合物合成、牛磺酸和亚牛磺酸代谢等多种途径可能是水稻对ZnO NPs和Cd胁迫响应的重要途径。
研究路线
研究结果
01
发芽率和种子活力指数
水稻品种(V)和Cd处理之间有统计学显著差异(p<0.05),而不同剂量的ZnO NPs处理在两种水稻品种的种子萌发中没有显著差异。种子萌发动态研究表明,Cd处理降低了水稻种子的萌发,尤其是在播种后3-5天,Cd和V×ZnO NPs×Cd对种子活力指数有显著影响,与无Cd处理相比,Cd胁迫使发芽率和活力指数分别降低5.59%和52.68%(图1)。
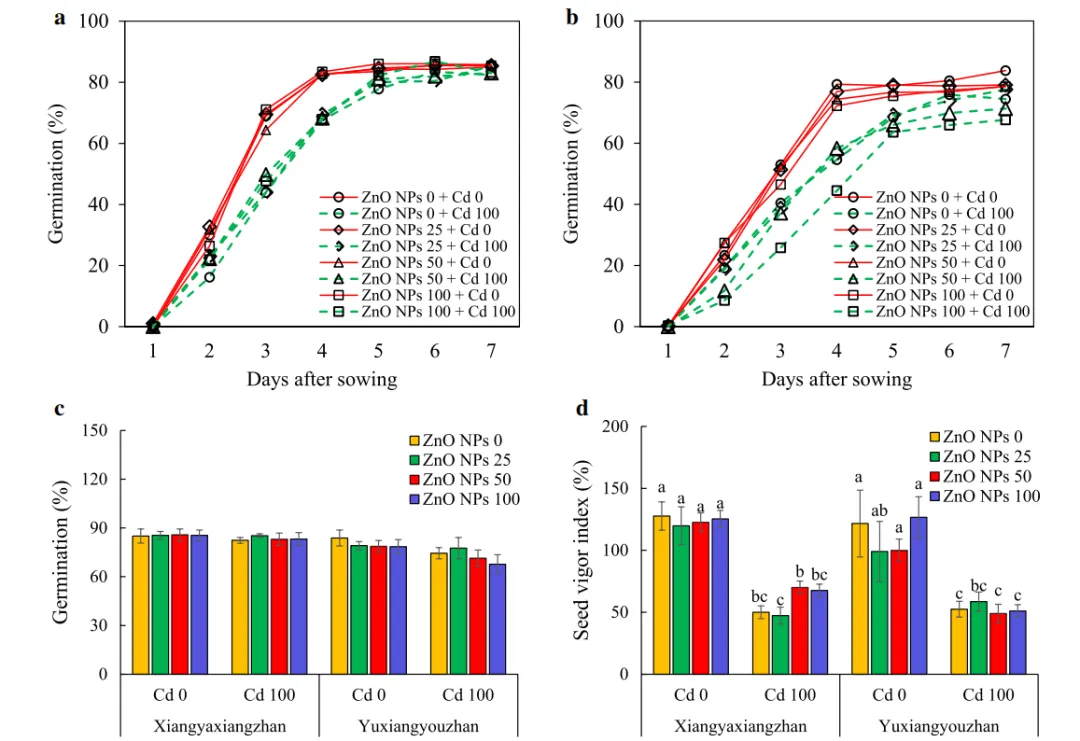
图1.(a)象牙香占和(b)玉香油占中ZnO纳米颗粒对种子萌发的动态影响,(c)象牙香占和玉香油占在Cd胁迫下种子萌发情况,(d)镉胁迫下,氧化锌纳米颗粒0、25、50和100mg·L−1,镉0和100mg·L−1的种子活力指数。表示为平均值±SD(n=4)。根据最小显著性差异检验,品种内不同处理之间的不同小写字母在p<0.05时显示统计学显著性。
02
形态生长属性
Cd、ZnO NPs和ZnO NPs×Cd对茎、根、先出叶、叶、叶鞘和叶片的长度也有显著影响。在正常条件下,ZnO NPs处理的水稻幼苗的茎长没有显著变化。在Cd(100mg·L-1)胁迫下,两个品种的水稻幼苗ZnO NPs处理后,茎长均有所提高。50mg·L-1和100mg·L-1的ZnO NPs处理下,象牙香占水稻幼苗的茎长显著提高(27.20%和26.84%);在玉香油占中,在不同ZnO NPs处理下,茎长增加29.90-34.79%(图2e)。此外,在25和100mg·L-1 ZnO NPs的象牙香占中,水稻幼苗的根长显著增加,在玉香油占中,正常条件下所有ZnO NPs处理都表现出水稻幼苗根长的增加,而基于ZnO NPs的处理对Cd胁迫下的根长没有影响(图2f)。分别在25和50mg·L-1的ZnO NPs施用量下,在有和没有Cd胁迫的情况下,玉香油占的先出叶片长度显著降低(图2g)。在正常条件下,100mg·L-1的ZnO NPs处理导致玉香油占叶鞘长度的显著改善,而在Cd胁迫下,所有ZnO NPs处理均显著增加叶鞘长度(图2h)。在正常条件下,在25和50mg·L-1的ZnO NPs处理下,玉香油占的叶片长度分别显著降低,在Cd胁迫下的象牙香占中,当用50和100mg·L-1 ZnO NPs处理时,叶片长度增加,且在所有ZnO NPs处理下的玉香油占中观察到类似的效果(图2i)。
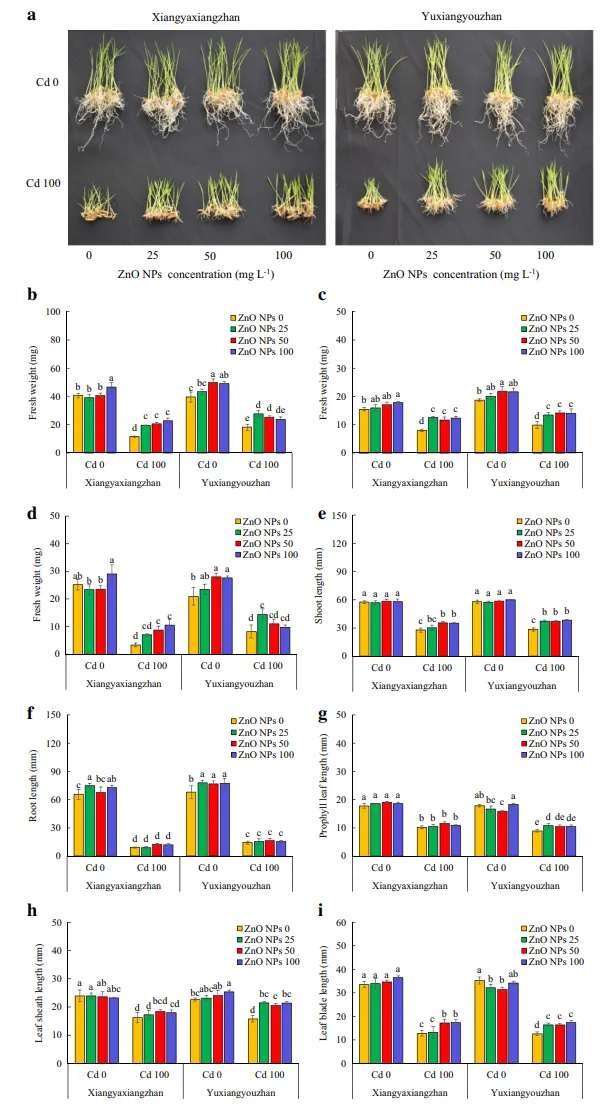
图2.水稻幼苗的生长和鲜重。水稻幼苗的生长(a),总鲜重(b),芽鲜重(c),根鲜重(d),芽长(e),根长(f),先出叶长(g),叶鞘长(h)和叶片长(i)。
03
水稻幼苗的淀粉酶活性
ZnO NPs和Cd处理显著影响了α-淀粉酶和总淀粉酶活性。对于玉香油占来说,在正常条件下利用不同剂量的ZnO NPs处理后,种子中的α-淀粉酶活性增加了30.29%,而在Cd胁迫(100mg·L-1)下,分别利用25和100mg·L-1的ZnO NPs处理后,种子中的α-淀粉酶活性显著增加。在正常条件下,50和100mg·L-1的ZnO NPs处理后,象牙香占水稻幼苗的α-淀粉酶活性均增加。两个水稻品种的α-淀粉酶活性在所有ZnO NPs处理下均显著增加,分别增加了17.45%-26.56%和10.56%-13.71%(图3a和b)。Cd胁迫及ZnO NPs处理对β-淀粉酶的活性没有显著影响(图3c和d)。在Cd胁迫下,分别在25和100mg·L-1 ZnO NPs处理下检测到象牙香占和玉香油占种子中总淀粉酶的最高活性。在正常条件下,在25、50和100mg·L-1 ZnO NPs下,玉香油占种子中的总淀粉酶活性显著增加了24.23%、16.20%和26.88%。此外,在Cd胁迫下,象牙香占和玉香油占的总淀粉酶活性分别增加了7.53-13.57%和9.58-16.64%,并且除了象牙香占中的100mg·L-1 ZnO NPs外,所有ZnO NPs处理均检测到统计学显著变化(图3e和f)。
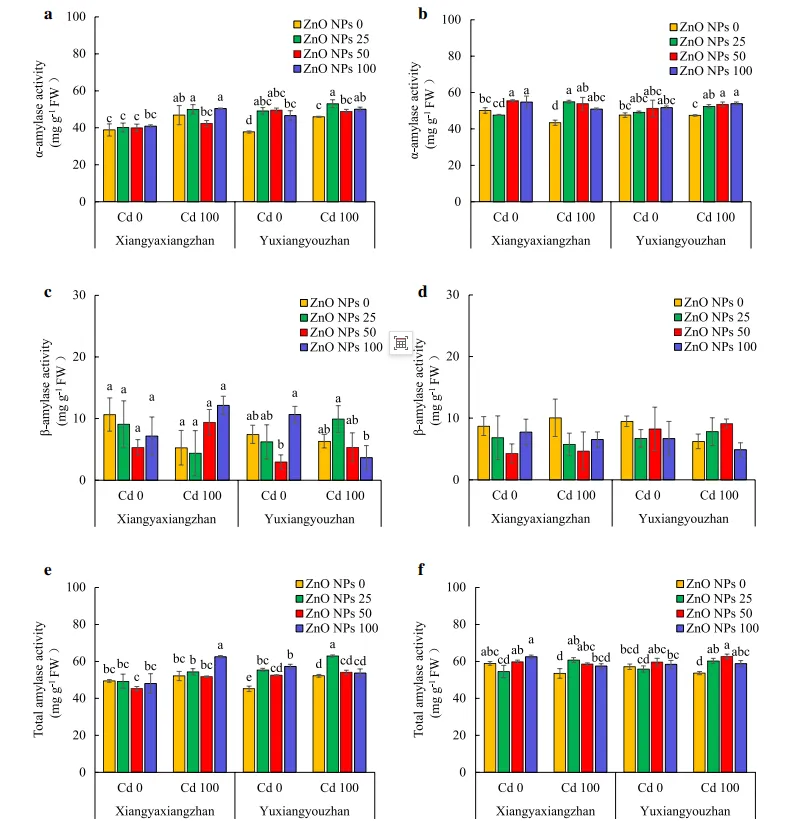
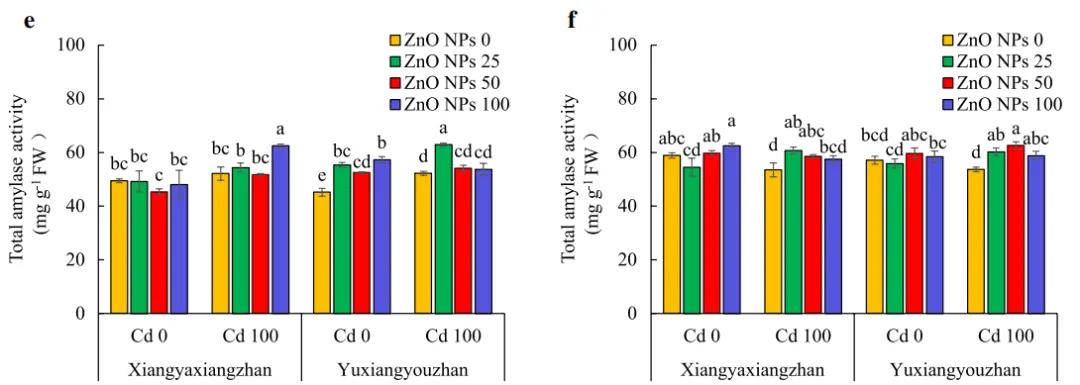
图3.水稻幼苗的α-淀粉酶、β-淀粉酶和总淀粉酶活性。地上部α-淀粉酶活性(a),幼苗α-淀粉酶活性(b),地上部β-淀粉酶活性(c),幼苗β-淀粉酶活性(d),地上部总淀粉酶活性(e),幼苗总淀粉酶活性(f)。
04
抗氧化酶活性
Cd和ZnO NPs显著影响了幼苗和地上部SOD、POD和CAT(过氧化氢酶)的活性。ZnO NPs×Cd显著影响地上部以及整个幼苗中的POD和CAT的活性。Cd胁迫下,25、50和100mg·L-1ZnO NPs处理的玉香油占地上部的SOD活性分别比对照提高了36.99%、51.37%和32.79%。在正常和Cd胁迫下,ZnO NPs处理组对象牙香占地上部SOD活性无显著影响。Cd胁迫下,水稻幼苗SOD活性略有升高,而正常和Cd胁迫下,ZnO NPs处理组的SOD活性无显著变化(图4a和b)。在无Cd胁迫条件下,50和100mg·L-1ZnO NPs处理的水稻幼苗地上部POD活性分别显著提高了36.23%和38.56%。在Cd胁迫下,25mg·L-1的ZnO NPs使Cd胁迫下水稻幼苗地上部POD活性提高了43.06%。在无Cd胁迫条件下,100mg·L-1的ZnO NPs显著降低了POD活性。在Cd胁迫下,25、50和100mg·L-1的ZnO NPs处理使玉香油占幼苗的POD活性分别提高了29.61%、43.05%和61.97%。在正常和Cd胁迫条件下,ZnO NPs处理组的玉香油占地上部和香芽香占幼苗的POD活性没有显著变化(图4c和4d)。正常条件下在象牙香占地上部利用50mg·L-1 ZnO NPs处理组的CAT活性显著升高,而在Cd胁迫下,各ZnO NPs处理组的CAT活性均显著降低。在正常条件下,25和50mg·L-1的ZnO NPs处理显著降低了象牙香占幼苗的CAT活性,而提高了玉香油占幼苗的CAT活性,而所有ZnO NPs处理组中的象牙香占以及25和100mg·L−1 ZnO NPs处理的玉香油占的CAT活性均显著降低(图4e和f)。
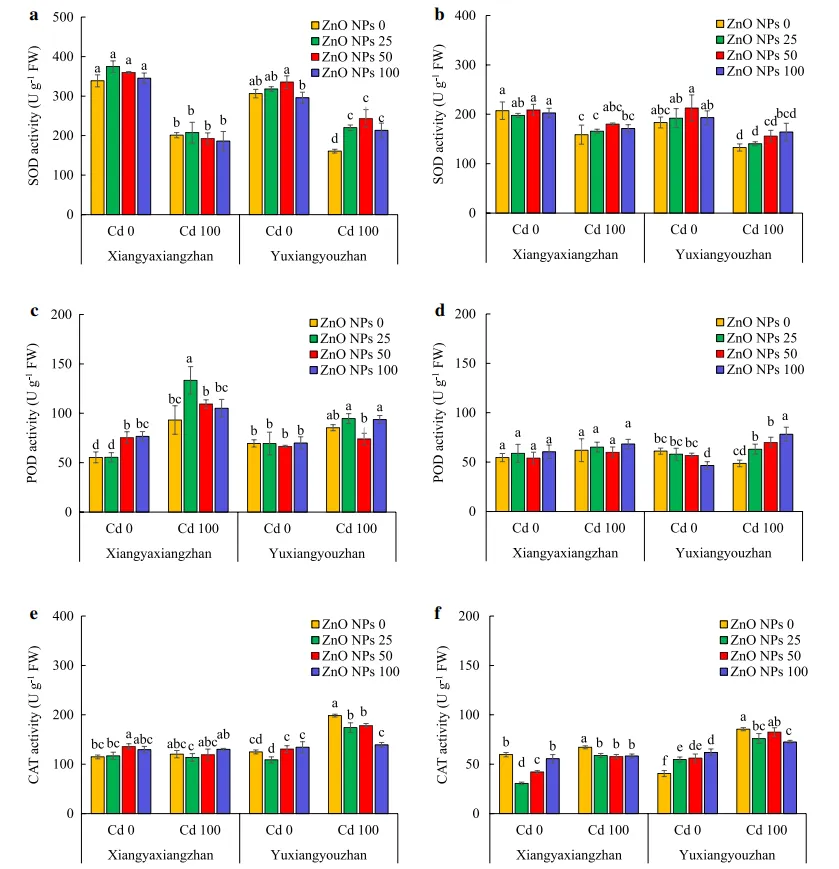
05
MDA含量和MT浓度
V、Cd、ZnO NPs和ZnO NPs×Cd显著影响MDA(丙二醛)含量和MT(金属硫蛋白)浓度。Cd胁迫下的象牙香占和玉香油占地上部MDA含量均未受到ZnO NPs处理的显著影响。在Cd胁迫下,ZnO NPs浓度为25、50和100mg·L-1时,MDA含量分别降低了26.94%、33.60%和15.88%。ZnO NPs处理对象牙香占(无Cd胁迫)和玉香油占(Cd胁迫和无Cd胁迫)幼苗MDA含量影响不显著,在Cd胁迫下,与未处理的对照(0mg·L-1 ZnO NPs)相比,50和100mg·L-1 ZnO NPs的MDA含量分别降低了16.63%和26.83%(图5a和b)。
在Cd胁迫下,25和50mg·L-1的ZnO NPs处理使象牙香占地上部的MT浓度分别增加了25.58%和23.90%,而100mg·L-1的ZnO NPs处理显著降低了玉香油占地上部的MT浓度。此外,在非胁迫及25和50mg·L-1ZnO NPs的情况下,玉香油占地上部的MT浓度分别增加了25.43%和19.68%。在正常条件下,50mg·L-1的ZnO NPs显着降低了象牙香占幼苗中的MT浓度,然而,在100mg·L-1的ZnO NPs显着增加了玉香油占幼苗中的MT浓度。此外,在Cd胁迫及50和100mg·L-1 ZnO NPs处理下,玉香油占幼苗中MT的浓度分别增加了21.77%和15.18%(图5c和d)。
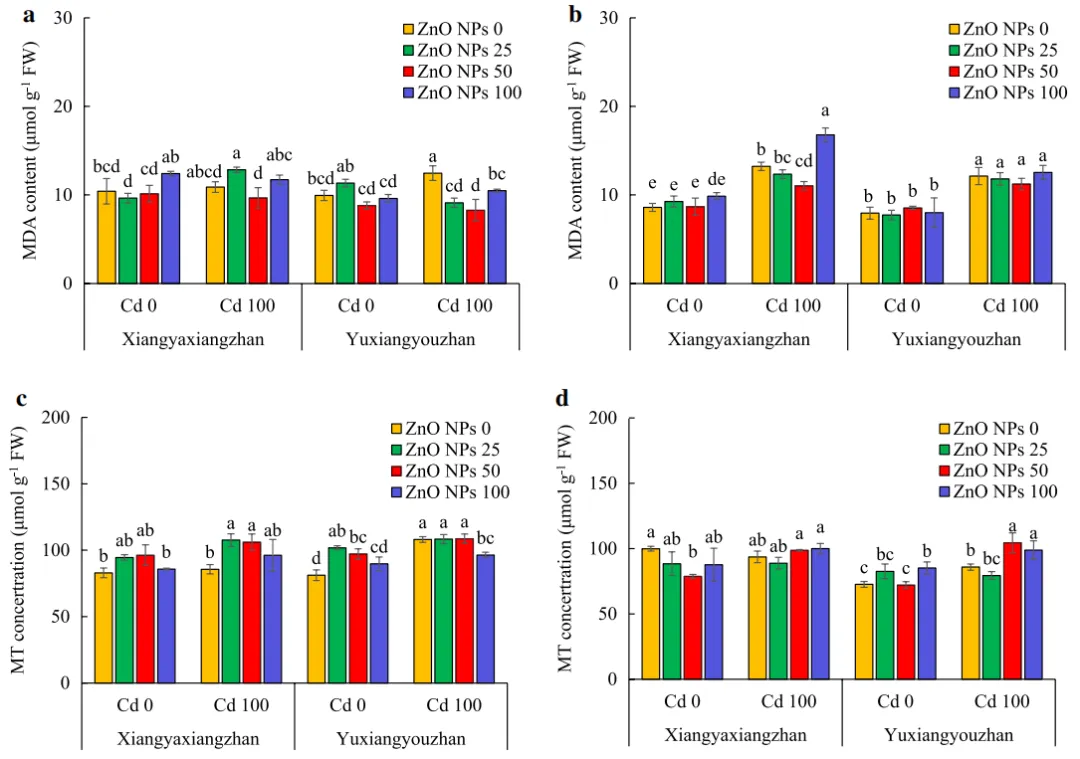
06
叶绿素含量、Zn和Cd的浓度
V、Cd、ZnO NPs和ZnO NPs×Cd显著影响叶绿素a、叶绿素b、叶绿素总含量和地上部类胡萝卜素的含量。ZnO NPs处理降低了正常和Cd胁迫下象牙香占和非胁迫下玉香油占地上部叶绿素a、叶绿素b、总叶绿素含量和类胡萝卜素含量。在Cd胁迫下,ZnO NPs处理提高了玉香油占地上部中的叶绿素a、叶绿素b、总叶绿素含量和类胡萝卜素。
V、ZnO NPs、ZnO NPs×Cd、V×ZnO NPs×Cd对幼苗Zn、Cd含量有显著影响。施用Cd显著提高了两个水稻品种幼苗中的Cd含量。两个品种的Zn含量在ZnO NPs处理下显著高于非ZnO NPs处理,而Cd含量在ZnO NPs处理下显著降低。在Cd胁迫下,100mg·L-1ZnO NPs显著增加了玉香油占幼苗中Cd的浓度(图6)。
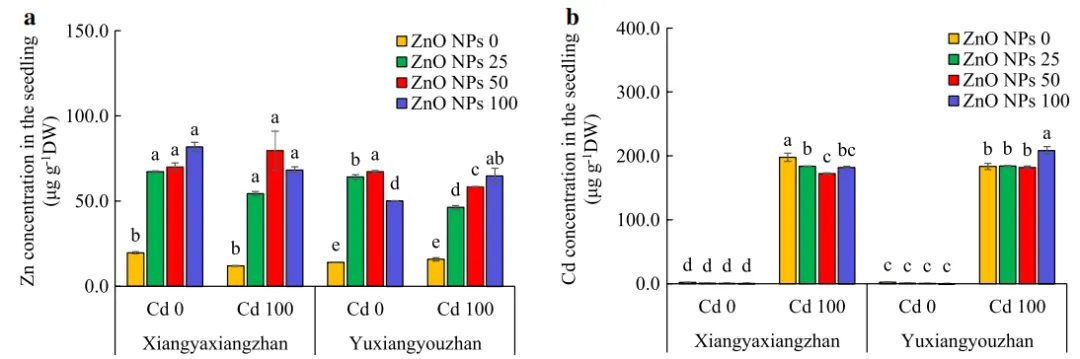
图6.水稻幼苗Zn、Cd含量。幼苗中Zn的浓度(a)和幼苗中Cd的浓度(b)。
07
代谢组检测和分析
在4种ZnO NPs处理中,50mg·L-1 ZnO NPs处理对Cd胁迫下两个水稻品种均有显著的缓解作用。取四种处理(A:ZnO NPs 0+Cd,B:ZnO NPs 0+Cd 100,C:ZnO NPs 50+Cd 0,和D:ZnO NPs 50+Cd 100)下的象牙香占和玉香油占的茎进行取样以分析代谢物。共检测到338种代谢物,包括72种氨基酸和衍生物、97种酚酸、57种脂质、44种有机酸、19种脂肪酸和醇、9种维生素和40种核苷酸和衍生物。
通过PCA(图7a)和PLS-DA(图7b)分析了四种处理下的两个水稻品种的茎,其中两个主成分共同解释了75%以上的方差,VIP值大于1的代谢物有26种,19种代谢物在处理组A中显示出相对含量最高,处理组B具有pmf 0585(dl-脯氨酸)和mws 0396(反式油酸)的相对含量最高,而处理组C中pmn 001627(没食子鞣质)是相对含量最高。处理组D的pme 0040(腺嘌呤)、pme 2634(dl-正缬氨酸)、mws 0256(L-缬氨酸)和mws 0001(L-天冬酰胺)值最高(图7c)。
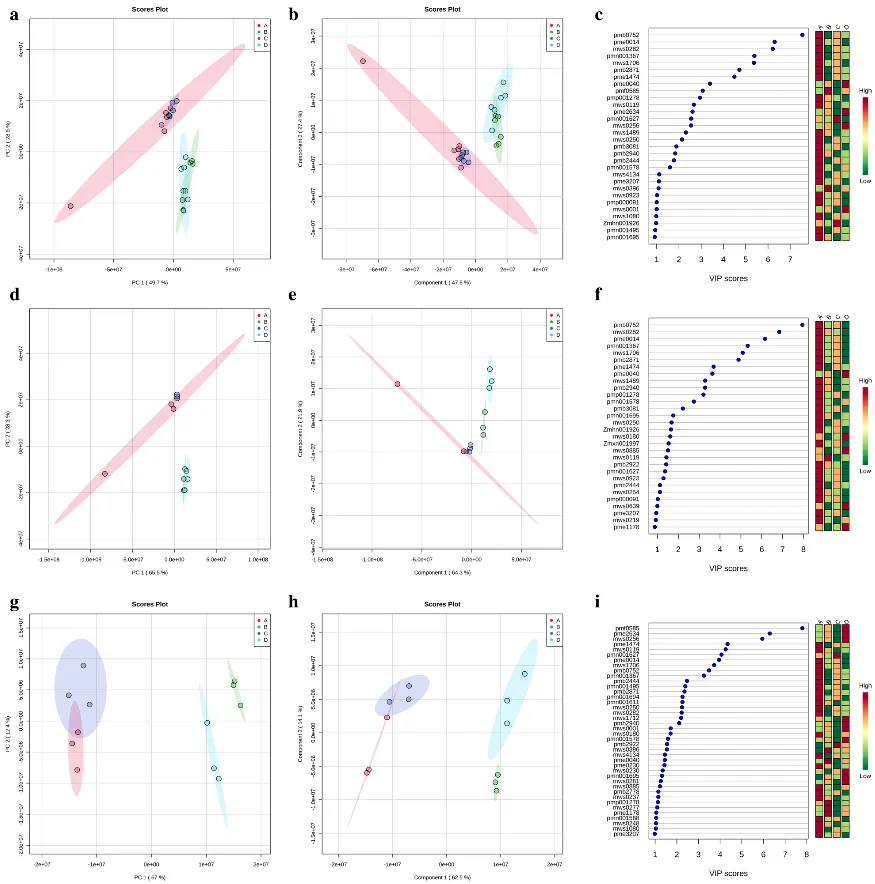
图7.代谢物的多变量分析。两个品种的代谢物的PCA分析(a),两个品种的代谢物的PLS-DA分析(b),比较两个品种的实验处理的PLS-DA的变量重要性投影(c)。象芽香占代谢产物的主成分分析(d),象芽香占代谢产物的PLS-DA分析(e),象芽香占试验处理的PLS-DA变量重要性投影比较(f)。玉香油占代谢产物的主成分分析(g),玉香油占代谢产物的PLS-DA分析(h),玉香油占各试验处理间的PLS-DA变量重要性投影比较(i)。A:ZnO NP 0+Cd 0;B:ZnO NP 0+Cd 100;C:ZnO NP 50+Cd 0;D:ZnO NP 50+Cd 100。
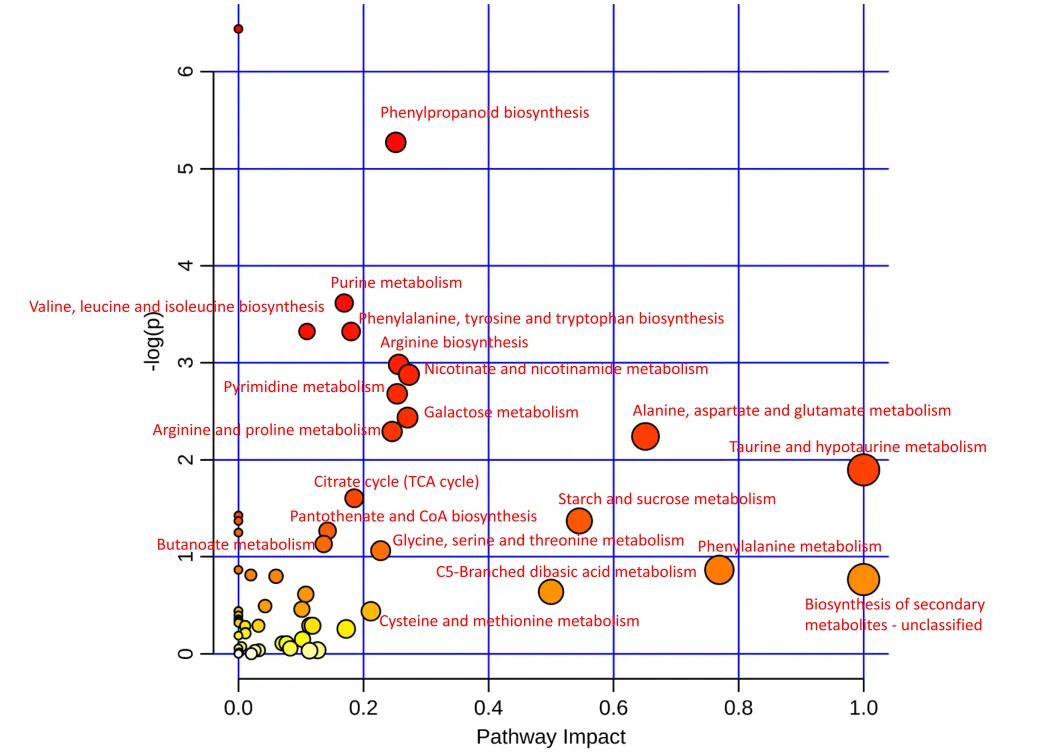
此外,“苯丙烷生物合成”、“嘌呤代谢”、“缬氨酸、亮氨酸和异亮氨酸生物合成”、“苯丙氨酸、酪氨酸和色氨酸生物合成”、“精氨酸生物合成”、“烟酸和烟酰胺代谢”、“嘧啶代谢”、“半乳糖代谢”、“精氨酸和脯氨酸代谢”、“丙氨酸、天冬氨酸和谷氨酸代谢”、“牛磺酸和亚牛磺酸代谢”等途径富集,检测到的代谢物中还涉及到“柠檬酸循环(TCA循环)”、“淀粉和蔗糖代谢”、“泛酸和CoA生物合成”、“丁酸代谢”、“甘氨酸、丝氨酸和苏氨酸代谢”、“苯丙氨酸代谢”、和“半胱氨酸和甲硫氨酸代谢”等代谢途径(图8)。
08
代谢组的差异分析和功能富集
构建了在所有实验处理下的象牙香占和玉香油占的水稻幼苗的地上部中的代谢谱的PCA。比较X轴(第一主成分)差异,X_A与X_B、X_A与X_C、X_A与X_D、X_B与X_D和X_C与X_D比较组中共鉴定出68个、7个、67个、33个和78个显著不同的代谢物;比较Y轴(第二主成分)的差异,分析Y_A与Y_B、Y_A与Y_C、Y_A与Y_D、Y_B与Y_D和Y_C与Y_D比较组,分别鉴定出55个、26个、42个、4个和28个显著不同的代谢物。此外,X_A与X_B、X_A与X_C、X_A与X_D、X_B与X_D和X_C与X_D比较组中分别鉴定出36个、5个、43个、9个和18个下调的代谢物,以及32个、2个、24个、24个和18个上调的代谢物;Y_A与Y_B、Y_A与Y_C、Y_A与Y_D、Y_B与Y_D和Y_C与Y_D比较组中分别鉴定出24个、22个、20个、2个和6个下调的代谢物,以及31个、4个、22个、2个和22个上调的代谢物。
对已鉴别的显著差异代谢物的KEGG富集分析进行了评估,并对不同处理象牙香占和玉香油占中已鉴别的显著差异代谢物根据变化倍数(FC)进行了排序和KEGG富集分析。
不同差异分组中差异明显的物质略有不同,代谢物如Hmfn 00531(L-抗坏血酸)、pme 3007(尿苷5'-二磷酸)、pme 0040(腺嘌呤)、pme 1474(5'-脱氧-5'-(甲硫基)腺苷)、pmb 0998(鸟苷5'-单磷酸)和mws 4134(氧化型谷胱甘肽)等在X_A和Y_A与X_B和Y_A与Y_B的FC值较大,而pme 0256(黄嘌呤)的代谢产物在X_A和X_C、Y_A和Y_C处均检测到FC值较大的代谢产物。
KEGG富集分析显示,ZnO NPs和Cd影响的代谢物参与代谢途径,例如“玉米素生物合成”、“嘌呤代谢”、“嘧啶代谢”、“谷胱甘肽代谢”、“半胱氨酸和甲硫氨酸代谢”、“精氨酸和脯氨酸代谢”、“不饱和脂肪酸的生物合成”、“咖啡因代谢”和“类黄酮生物合成”、“芪类化合物”。
此外,不同处理组中差异物质不同,与ZnO NPs 0+Cd 0处理相比,ZnO NPs 0+Cd 100处理引起5′-单磷酸腺苷、5′-脱氧-5′-(甲硫基)腺苷,鸟苷5'-单磷酸,胞苷5'-单磷酸(胞苷酸)和尿苷5’-单磷酸,以及上调的腺嘌呤、尿苷5’-二磷酸、腺嘌呤、黄嘌呤、尿苷5’-二磷酸,L-抗坏血酸、二十碳烯酸、二十碳二烯酸和黄嘌呤。ZnO NPs 50+Cd 0处理导致黄嘌呤的减少。此外,ZnO NPs 50+Cd 100处理上调了腺嘌呤、黄嘌呤、L-抗坏血酸、二十碳烯酸和二十碳二烯酸。且代谢途径的代谢物因品种和ZnO NPs处理而异,与玉香油占相比,检测到象牙香占的D/B和D/C中更多基于代谢物的变化。玉香油占中的(5-L-谷氨酰)-L-氨基酸高于象牙香占。
讨论与总结
综上所述,ZnO NPs对水稻种子萌发无明显影响,但通过提高Cd胁迫下水稻幼苗的相关生理生化反应(如淀粉酶和抗氧化酶的活性)、调节Zn浓度和幼苗对ZnO NPs的吸收以及其他代谢产物的吸收,显著促进了幼苗的生长。今后仍需通过田间试验进一步研究ZnO NPs对镉毒害条件下水稻早期生长的影响机制。总体而言,研究结果可能有助于制定在农业种植经济重要的作物与高浓度的重金属污染方面可行的战略。



END
推 荐 阅 读
1、开学季狂欢,好礼享不停!特价、免单、预存全都有!快来捕捉属于你的惊喜礼包~
2、合作文章丨平顶山学院张志录团队揭示了海桐叶斑对低温的保护作用及其生理生化和分子机制
3、助力科研丨普奈斯水体检测指标介绍


