选择性自噬是宿主清除病毒组分的关键防御机制,但冠状病毒已进化出多种策略逃逸这一监控。猪δ冠状病毒(PDCoV)作为新发肠道冠状病毒,可引起仔猪严重腹泻和高死亡率,造成重大经济损失,且具备跨种传播潜力。尽管已知冠状病毒主蛋白酶NSP5通过切割NBR1等自噬受体干扰宿主防御,但是否存在其他被靶向的自噬受体尚不清楚。
2026年3月12日,华中农业大学研究团队在mBio上发表题为“PDCoV NSP5 cleaves the selective autophagy receptor CCDC50 to disrupt autophagic degradation of the viral envelope protein”的最新研究论文。本研究揭示CCDC50通过识别病毒E蛋白的K63泛素化修饰介导其自噬降解,而PDCoV NSP5切割CCDC50于保守位点Q171使其失活。
IF:5.1 中科院1区 | JCR/Q1 微生物学 参考译文:PDCoV NSP5切割选择性自噬受体CCDC 50以破坏病毒包膜蛋白的自噬降解 第一作者:Ke Li 通讯作者:Rui Luo |
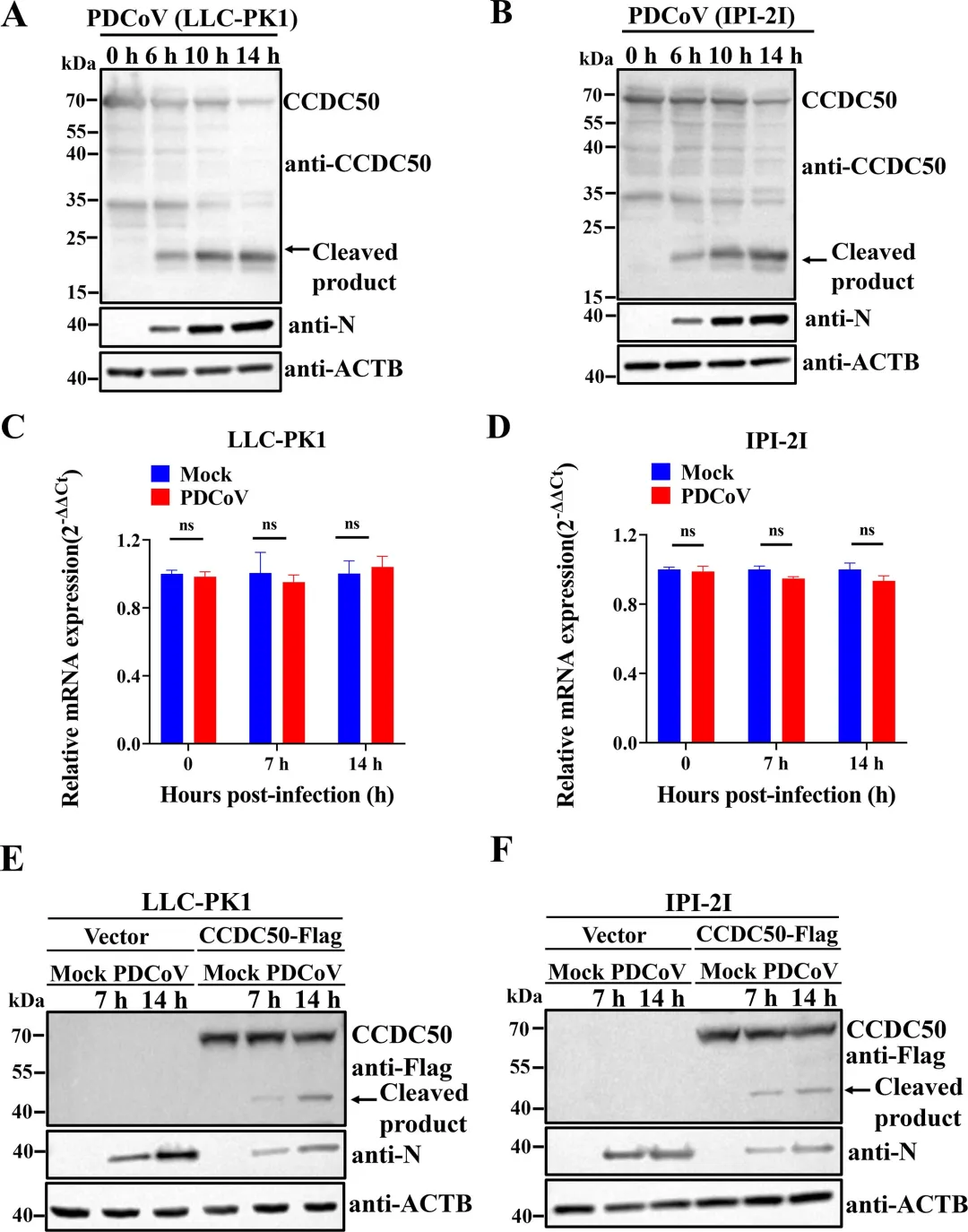
1.PDCoV感染诱导CCDC 50的蛋白水解切割
PDCoV感染LLC-PK1和IPI-2J细胞后,CCDC50蛋白水平呈时间依赖性下降,并伴随出现约20 kDa的N端切割片段。通过RT-qPCR检测,CCDC50的mRNA水平在感染后无明显变化,表明该切割发生在蛋白水平而非转录水平。为进一步验证,研究者构建了C端带有Flag标签的CCDC50表达载体,转染细胞后进行PDCoV感染,Western blot检测到约45 kDa的C端切割片段,进一步证实PDCoV感染可诱导CCDC50发生特异性切割。
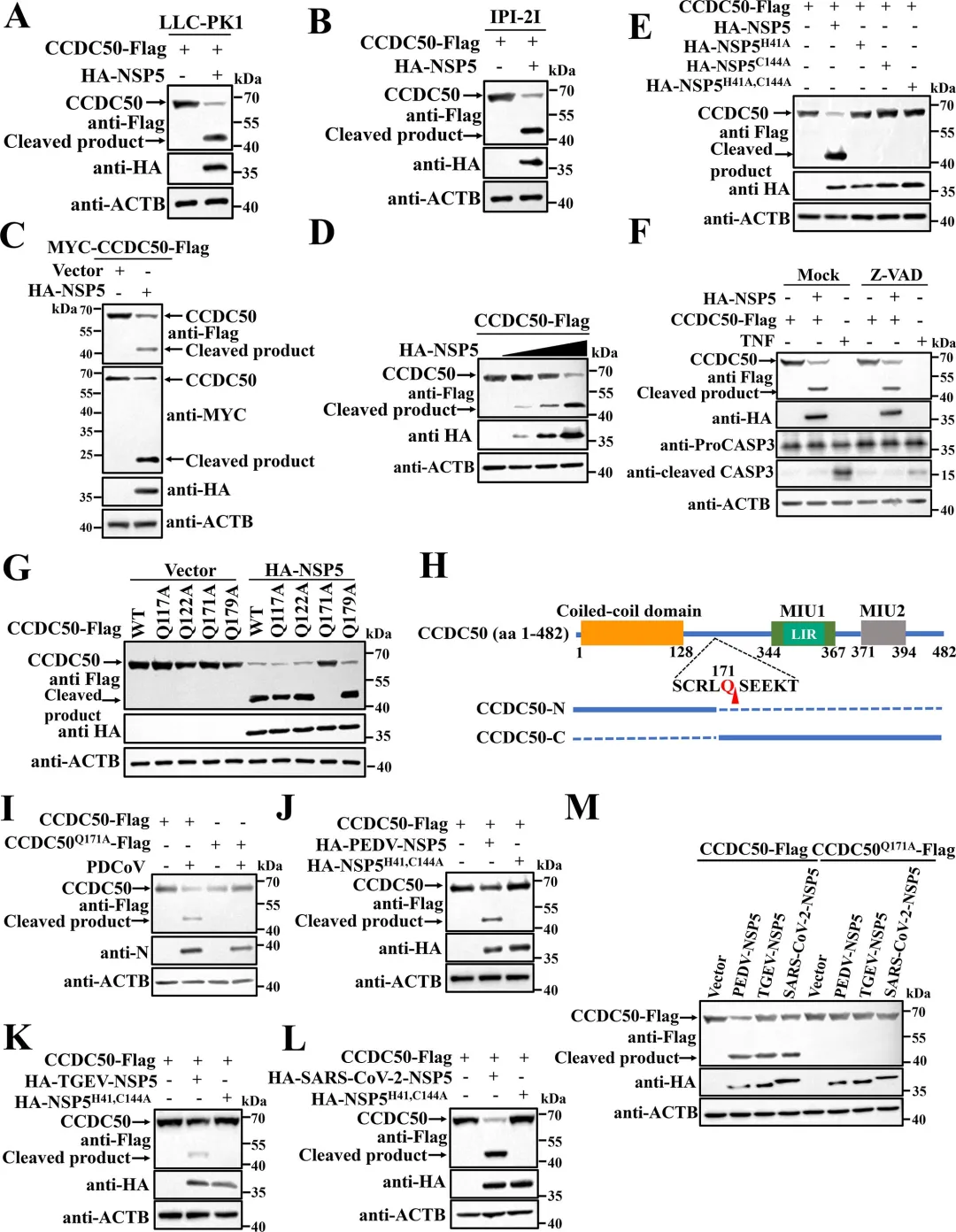
2.PDCoV NSP5在残基Q171处切割CCDC 50
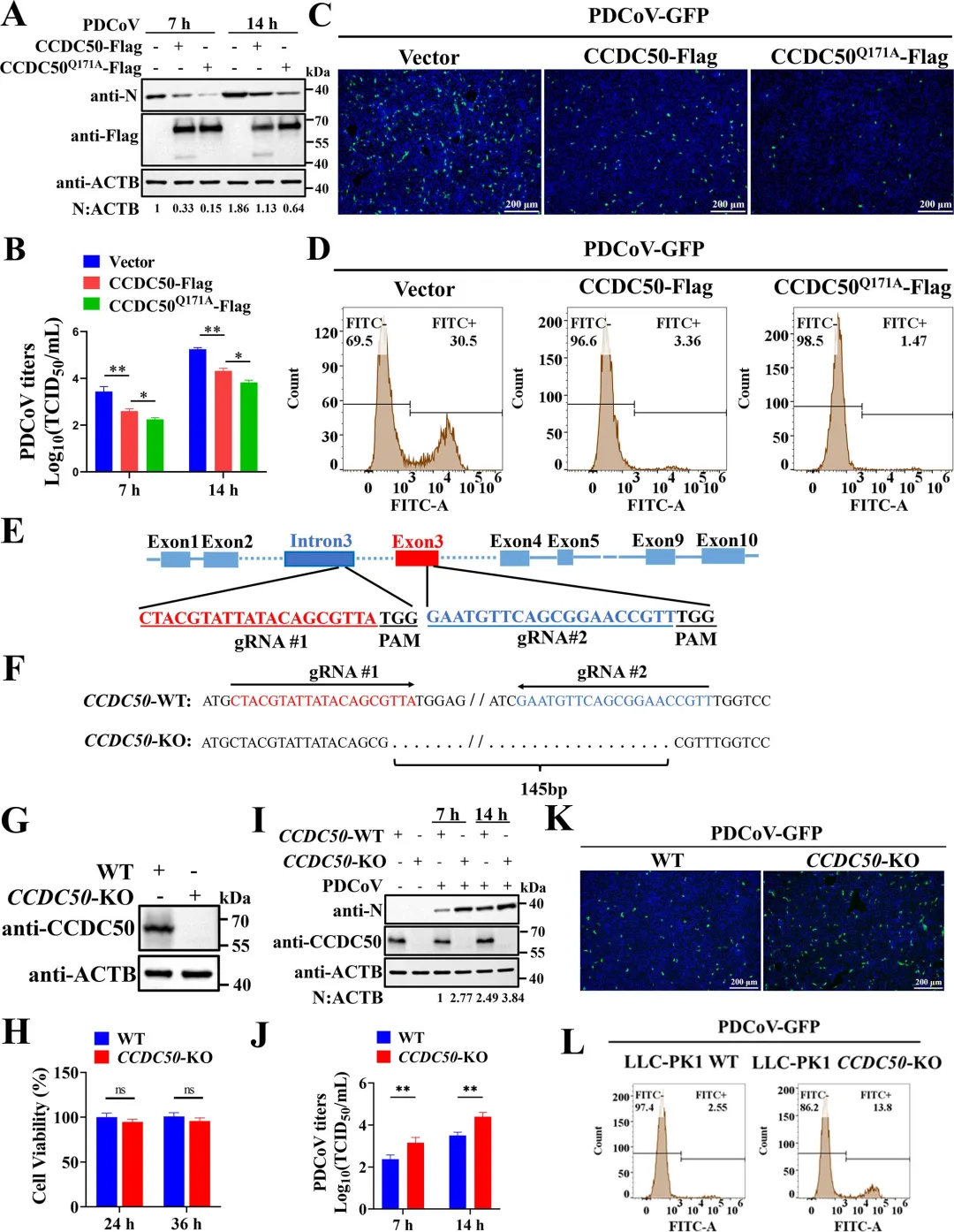
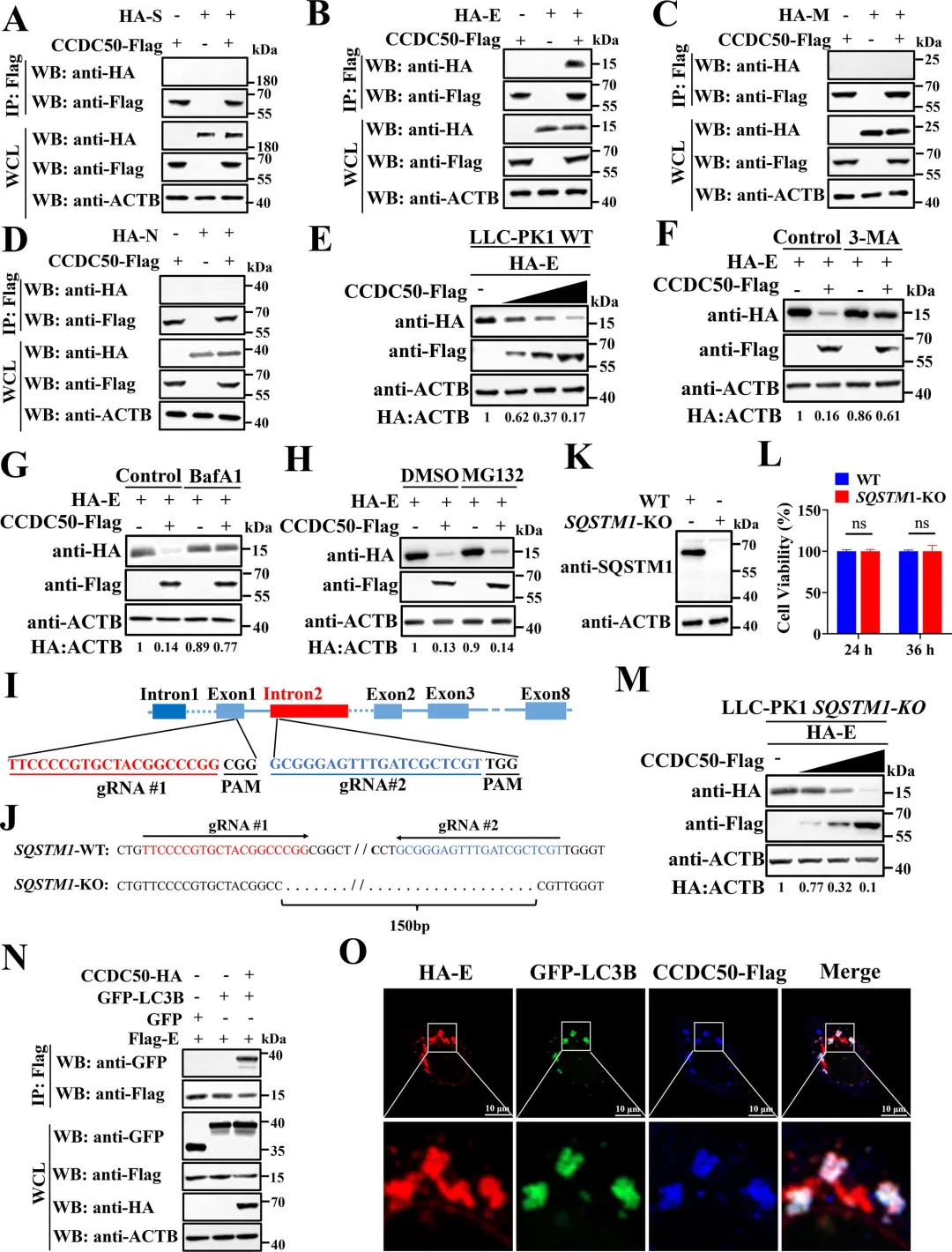
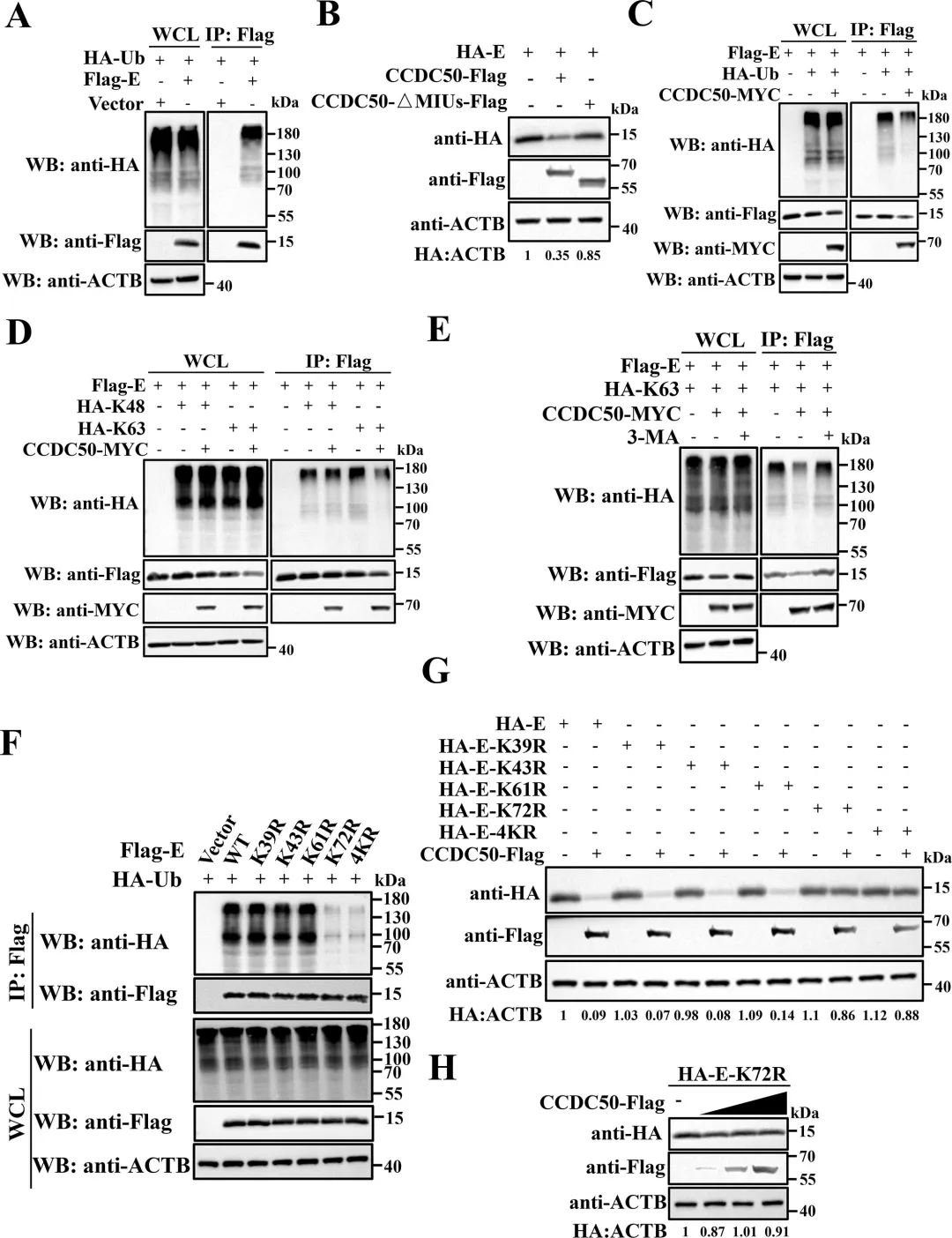
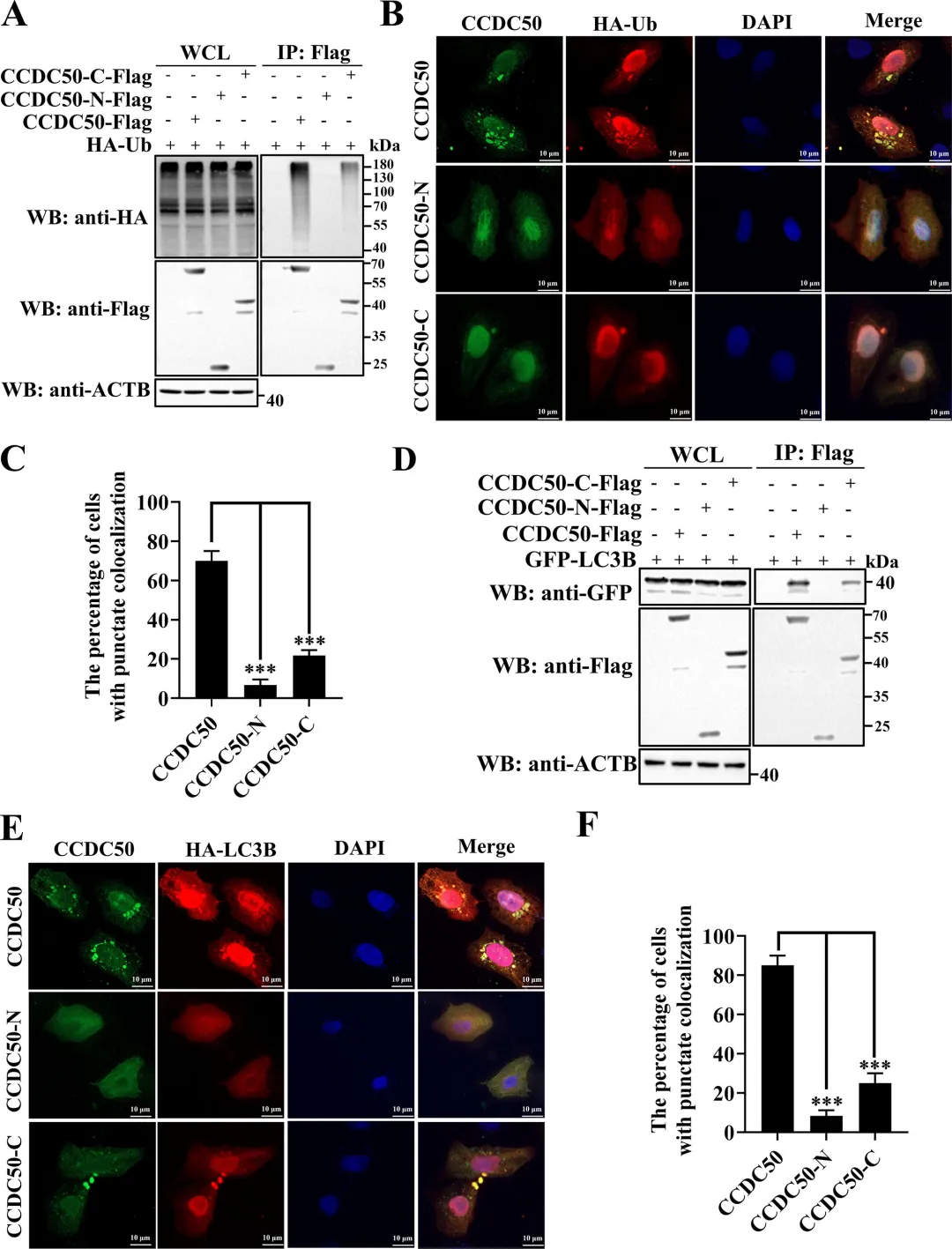
通过共表达实验发现,PDCoV的NSP5蛋白酶可诱导CCDC50产生与病毒感染时大小一致的C端切割片段,而催化失活的NSP5突变体则无此作用,表明切割依赖NSP5的蛋白酶活性。利用双标签构建体,研究者明确了切割产生约20 kDa的N端片段和约45 kDa的C端片段。结合冠状病毒NSP5切割底物需在P1位点为谷氨酰胺(Q)的特异性,研究者对CCDC50上多个候选Q残基进行点突变,结果显示仅Q171A突变可完全阻断NSP5介导的切割以及PDCoV感染诱导的切割,确定Q171为关键切割位点。图2.PDCoV NSP 5通过其蛋白酶活性在Q171处切割CCDC 50研究者通过过表达和基因敲除两方面验证了CCDC50对PDCoV复制的调控作用。过表达野生型CCDC50可显著降低PDCoV N蛋白表达水平及病毒滴度,而切割抗性突变体CCDC50𝑄171𝐴Q171A的抑制效果更强,表明NSP5介导的切割削弱了CCDC50的抗病毒功能。利用PDCoV-GFP报告病毒,荧光显微镜和流式细胞术进一步证实CCDC50过表达可减少病毒感染比例和荧光强度。相反,通过CRISPR-Cas9构建的CCDC50敲除细胞系中,PDCoV N蛋白积累、病毒滴度和GFP阳性细胞比例均显著升高,且细胞活力未受敲除影响。通过免疫共沉淀实验发现,CCDC50特异性结合PDCoV的E蛋白,而与S、M、N蛋白无显著互作。在LLC-PK1细胞中过表达CCDC50可呈剂量依赖性地降低E蛋白水平,且该降解作用可被自噬抑制剂3-甲基腺嘌呤和巴弗洛霉素A1阻断,但不受蛋白酶体抑制剂MG132影响,表明CCDC50通过自噬-溶酶体途径介导E蛋白降解。为明确CCDC50是否依赖经典自噬受体,研究者构建了SQSTM1/p62敲除细胞系,结果显示CCDC50仍能有效降低E蛋白水平,表明其功能独立于SQSTM1。进一步在NBR1/CCDC50双敲除细胞中回补实验表明,CCDC50与NBR1分别以相互独立的方式介导E蛋白降解。此外,CCDC50过表达可增强E蛋白与LC3B的互作及共定位,提示其作为桥接分子将E蛋白招募至自噬体。图4.CCDC 50与PDCoV E相互作用并介导其自噬降解5.CCDC50识别PDCoV E蛋白的K63连接的泛素化研究者探究了CCDC50识别E蛋白的分子基础。由于CCDC50通过其MIU结构域结合泛素,研究者首先验证了E蛋白存在泛素化修饰,并发现缺失MIU结构域的CCDC50突变体无法降解E蛋白,表明泛素结合能力是其介导降解的前提。进一步实验显示,CCDC50过表达可降低E蛋白的K63连接泛素化水平,但对K48连接泛素化无显著影响,且该效应可被自噬抑制剂3-甲基腺嘌呤阻断,提示CCDC50选择性地识别K63泛素化修饰并通过自噬途径清除底物。为精确定位E蛋白上的泛素化位点,研究者构建了多个赖氨酸突变为精氨酸的点突变体,发现K72R突变及四个赖氨酸位点联合突变(4KR)均显著削弱E蛋白的泛素化水平。图5.CCDC50识别PDCoV E蛋白的K63连接的泛素化6.NSP5介导的CCDC50切割损害其选择性自噬功能研究者探讨了切割对CCDC50作为自噬受体功能的影响。通过免疫共沉淀实验发现,全长CCDC50能够与泛素及LC3B发生强效结合;而N端切割片段(CCDC50-N)由于缺失MIU和LIR结构域,完全丧失了与泛素和LC3B的相互作用;C端切割片段(CCDC50-C)虽保留了MIU结构域,但其结合能力较全长蛋白显著减弱。免疫荧光共定位分析进一步证实,全长CCDC50与泛素及LC3B呈现明显的点状共定位,而CCDC50-N几乎无共定位,CCDC50-C的共定位程度亦明显降低。这些结果表明,NSP5介导的切割破坏了CCDC50的模块化结构,使其无法同时有效结合泛素化底物和自噬体膜蛋白LC3B,从而削弱了其作为选择性自噬受体的货物递送功能。图6.CCDC50的裂解削弱了其在选择性自噬中的功能7.NSP5介导的切割片段减弱CCDC 50抗病毒活性
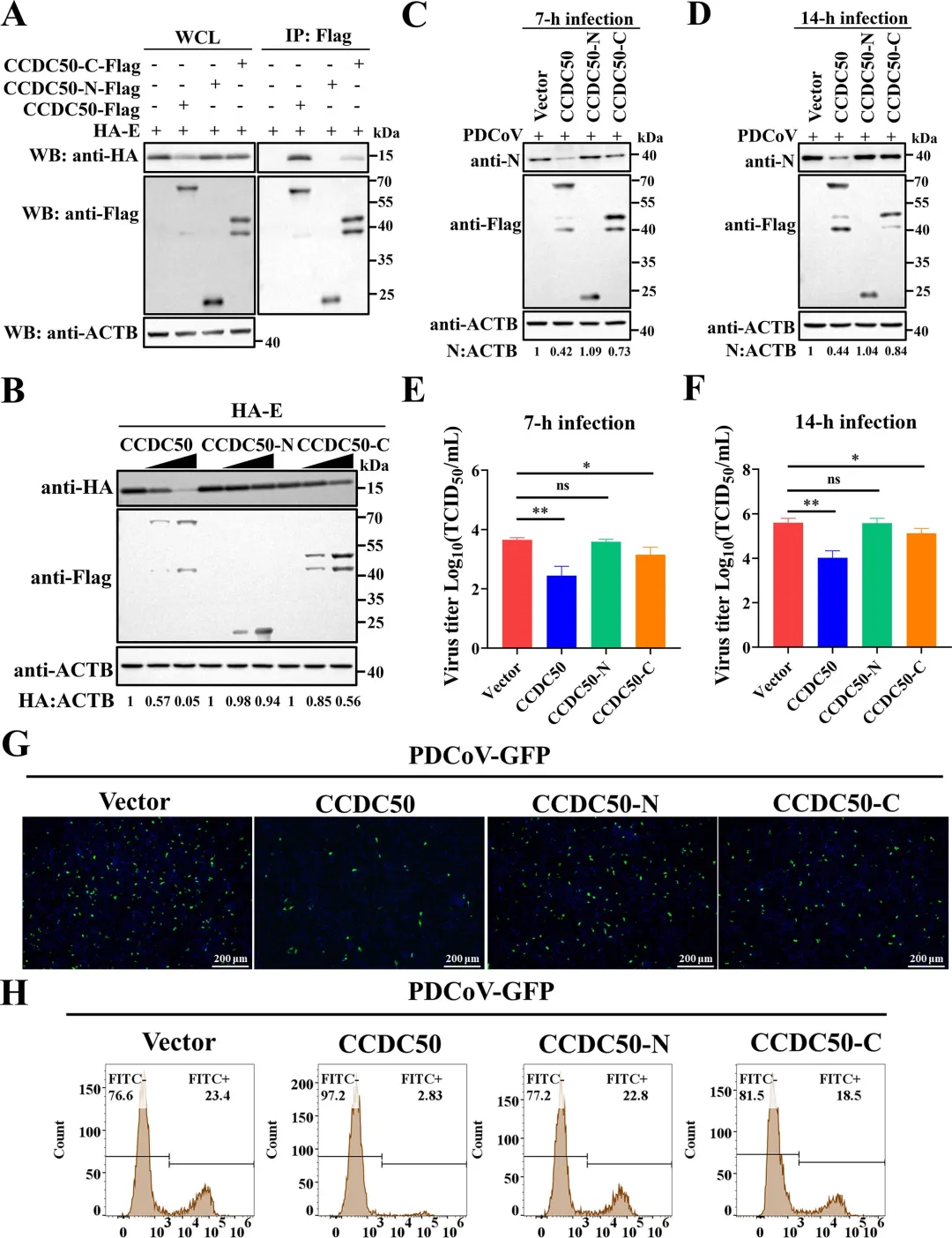
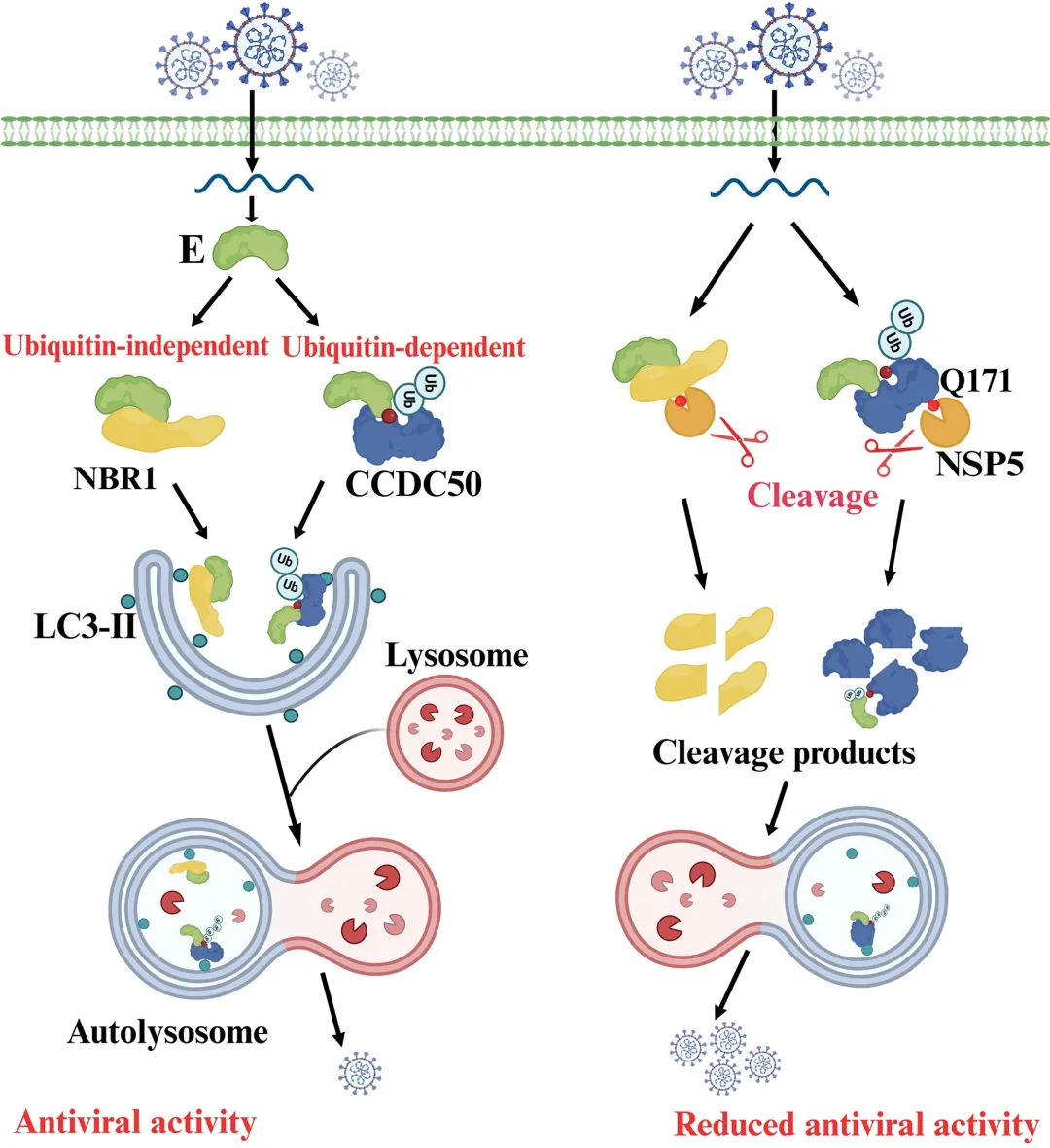
免疫共沉淀结果显示,全长CCDC50可与PDCoV E蛋白高效结合;CCDC50-N端片段完全丧失与E蛋白的结合能力;而CCDC50-C端片段虽保留微弱互作,但结合效率较全长显著降低。功能实验进一步表明,在LLC-PK1细胞中过表达全长CCDC50可呈剂量依赖性降低E蛋白水平,并有效抑制病毒N蛋白表达及病毒滴度;CCDC50-N完全丧失上述抑制作用;CCDC50-C仅表现出部分抑制效果。利用PDCoV-GFP报告病毒进行的荧光显微镜及流式细胞术分析进一步证实,全长CCDC50显著减少病毒感染比例,而CCDC50-N无抑制作用,CCDC50-C的抑制效果亦明显减弱。本研究首次揭示CCDC50作为一种新型选择性自噬受体,在猪丁型冠状病毒(PDCoV)感染中发挥重要的抗病毒作用。机制研究表明,CCDC50通过其MIU结构域特异性识别E蛋白K72位点的K63连接多聚泛素化修饰,并与LC3B相互作用,将病毒E蛋白靶向自噬-溶酶体途径进行降解,从而限制病毒复制。这一过程独立于经典自噬受体SQSTM1/p62和NBR1,揭示了宿主抗冠状病毒防御的新机制。
与此同时,PDCoV演化出精密的免疫逃逸策略。病毒的NSP5蛋白酶通过其水解酶活性,在CCDC50蛋白的Q171位点对其进行特异性切割,产生N端和C端两个片段。切割后的CCDC50-N完全丧失与泛素、LC3B及E蛋白的结合能力,CCDC50-C虽保留部分互作但功能显著减弱,导致CCDC50介导的选择性自噬降解功能受损,抗病毒活性大幅下降。
值得注意的是,这一切割机制在冠状病毒中高度保守。不仅PDCoV的NSP5,来自PEDV、TGEV甚至SARS-CoV-2的NSP5同样能够识别并切割CCDC50的Q171位点。该发现揭示了冠状病毒通过NSP5蛋白酶靶向宿主自噬受体、逃避选择性自噬清除的共同进化策略,为理解冠状病毒与宿主互作机制提供了新视角,也为抗病毒药物研发提供了潜在靶点。
源论文链接:https://journals.asm.org/journal/mbio on 29 March 2026 by 58.58.135.3
编辑:九九
投稿合作:shouyiyanquan@163.com
可为课题组代发文章宣传、新闻通稿、招聘等
| 标注“原创”仅代表原创编译,原文版权归原作者所有。本文仅用作学术交流分享,如有错误或侵权请后台私信订正或删除。 |


 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
