Title:Rhizosphere Bacteria Help to Compensate for Pesticide-Induced Stress in Plants
Publication Date:2024/7/5江苏省农业科学院农产品质量安全与营养研究所余向阳研究员为通讯作者
https://newsjs.jaas.ac.cn/jgsz/xrld/art/2020/art_c4dd0e9bdd9c4dfead766338668ca8cd.html
摘 要
外源化学物质调控植物生长时往往呈现双相响应特征,即低剂量产生刺激效应、高剂量表现为抑制作用,但其内在作用机制尚未明确。本研究首次证实,根际微生物群具有补偿性功能,可助力植物抵御农药胁迫。研究发现,小白菜经低、高剂量吡虫啉叶面喷施处理后,根际细菌总数有所增加,且多种有益细菌得到富集,如诺卡氏菌属、短波单胞菌属、鞘氨醇单胞菌属等,这类细菌兼具植物促生和农药降解能力。受农药胁迫的小白菜会通过增加根系分泌物中初级代谢物(如氨基酸、脂肪酸、溶血磷脂酰胆碱)的释放,完成对上述有益细菌群落的招募。低剂量农药处理下,微生物的补偿效应可抵消农药胁迫作用,进而促进植物生长;而高剂量农药处理时,微生物的补偿效应不足以抗衡农药胁迫,最终导致植物生长受到抑制。本研究为制定优化的农药施用策略奠定了基础,也为深入理解如何利用根际微生物群、以生态友好的方式缓解作物的化学诱导胁迫提供了新的思路。
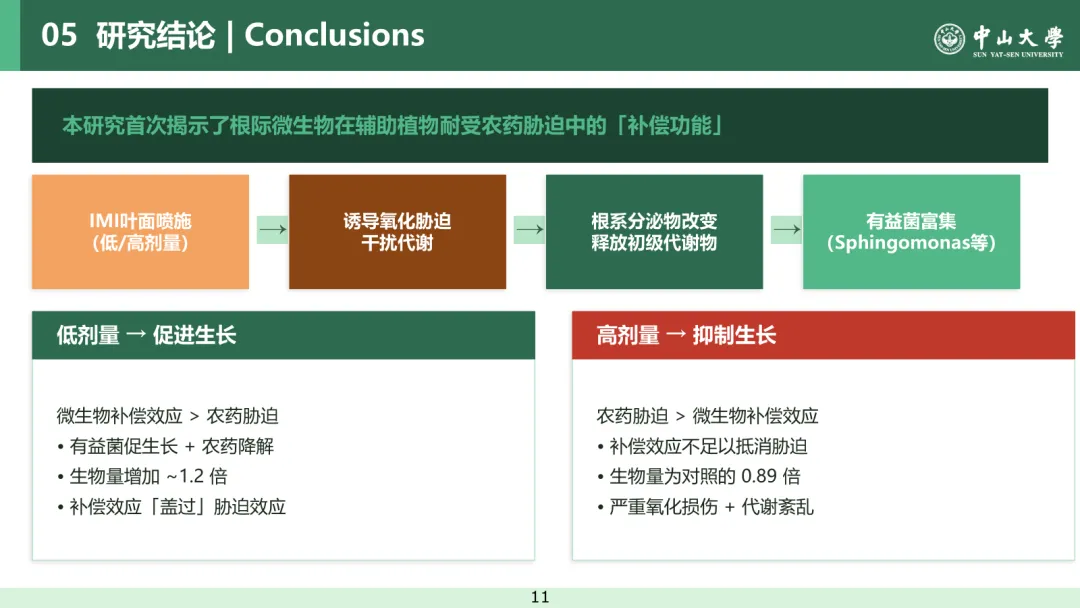
本研究以吡虫啉(IMI) 为模式农药、小白菜为研究对象,首次揭示了根际微生物群的补偿作用在缓解农药诱导植物胁迫中的核心机制:叶面喷施低、高剂量吡虫啉均会使小白菜根际细菌总数增加并富集诺卡氏菌属、短波单胞菌属、鞘氨醇单胞菌属等兼具促生和农药降解能力的有益菌,该过程由植物通过释放氨基酸、脂肪酸等根系分泌物初级代谢物驱动;低剂量下微生物补偿效应抵消农药胁迫并促进植物生长,高剂量下该效应不足以抗衡胁迫导致植物生长受抑,同时土壤系统相较水培系统能显著提升植物对吡虫啉胁迫的抗性,本研究为优化农药施用策略、利用根际微生物实现生态化缓解作物化学胁迫提供了理论依据。
引言
农药虽在防治作物病虫害方面不可或缺,却会诱导植物产生氧化胁迫,改变其生理代谢过程⁽¹⁾。草甘膦、百草枯、吡虫啉(IMI)、噻虫嗪等多种农药,在调控植物生长时均被证实会产生双相响应,呈现低剂量刺激生长、高剂量抑制生长的特征⁽²⁻⁴⁾,该现象被称为兴奋效应⁽⁵⁾。农药的低剂量刺激效应,通常被认为是直接的生长促进作用,或是植物的过度补偿性响应,这种效应或能增强植物对后续胁迫的抗逆能力⁽⁶⁾。例如,研究表明吡虫啉可激活植物与水杨酸相关的防御反应,进而提升作物长势及对生物胁迫的抗性⁽⁷⁾;而草甘膦则会通过调控莽草酸途径减少植物木质素的合成,最终促进作物生长、提高产量⁽⁸⁾。(研究背景及进展)但现有相关研究多仅聚焦于植物自身的响应机制,且研究结论仍存争议⁽⁹⁾。受农药胁迫的植物是否会借助外界因素抵御农药胁迫、调控自身生长,目前尚未明确。(研究空白)因此,亟需开展更全面的研究阐明这一过程,以此丰富植物环境适应性的理论体系。
本研究以吡虫啉为模式农药,系统阐释了农药施用量、植物生长与根际细菌群落三者间的关联。在此基础上,本研究验证了根际细菌对植物生长和农药降解的补偿效应,同时探究了这类细菌与根系分泌物代谢物之间的调控关系。随后,研究构建了一套严谨的假说以解释这一兴奋效应,该假说着重阐明了微生物补偿作用与农药剂量依赖性胁迫的联合作用机制。本研究的相关发现为优化农药施用方式开辟了新途径,为揭示植物与根际微生物群的互作规律提供了新线索,也为推动农业可持续栽培技术的发展奠定了新基础。(研究内容和解决的科学问题)
2.1 植株培养
2.1.1 土壤培养
试验材料包括小白菜(Brassica rapa var. perviridis)和农田表层土壤,均取自中国江苏省农业科学院。供试土壤的颗粒组成为砂粒 30%、黏粒 53%、粉粒 17%,有机质含量为 2%;采用土水比 1:2.5(质量体积比)测定土壤 pH,结果为 6.1⁽¹⁸⁾。
选取约 200 株小白菜幼苗,单株种植于装有 150 g 供试土壤的塑料花盆中,置于人工气候箱内培养,培养条件为:光照强度 190 微摩尔・米⁻²・秒⁻¹,相对湿度 68%,温度 28℃,每日光照 - 黑暗周期为 10-14 小时。幼苗培养 2 周后,对每株幼苗喷施 2.0 mL 含 0.1% 聚山梨酯 - 80 与吡虫啉的水溶液(吡虫啉为分析纯,购自美国密苏里州圣路易斯市西格玛 - 奥德里奇公司),吡虫啉浓度设置为 10~100 毫克 / 升;对照组喷施等体积含 0.1% 聚山梨酯 - 80 但不含吡虫啉的水溶液。喷施前用铝箔遮盖花盆口,防止农药落入培养基质。农药暴露 2 天后,采用日本美能达 SPAD-502 型叶绿素仪测定小白菜叶片叶绿素含量,同时采集小白菜地上部样品,用于生理指标和代谢组学分析;其中代谢组学分析用的地上部样品经液氮速冻、研磨成粉后,置于 - 80℃超低温冰箱中保存。剩余植株继续培养 20 天,培养结束后采集小白菜地上部、根系及根际土壤样品,用于吡虫啉及其降解产物的检测;记录植株鲜重,并采集根际土壤样品,开展细菌分析与微生物分离提取工作。此外,收集各处理组的栽培土壤,用于后续重植试验。
2.1.2 水培培养
选取约 160 株小白菜幼苗,单株培养于装有 50 毫升木村 B 营养液的琥珀色玻璃瓶中,在人工气候箱内培养 20 天。吡虫啉的施用操作严格遵循 2.1.1 小节的试验方案进行。吡虫啉施用 2 天后,每组选取 6 株幼苗,用无菌水冲洗 3 次,随后转移至装有 50 毫升无菌去离子水的琥珀色玻璃容器中,继续培养 2 天。培养结束后,收集剩余水溶液(根系分泌物),经 0.4 微米孔径滤膜过滤后,于 - 20℃保存,用于代谢谱分析。其余不同处理组的幼苗继续培养 20 天,培养期间采集小白菜的地上部、根系及营养液样品,用于吡虫啉含量检测,并记录植株鲜重。
2.2 生理活性分析
选取 2.1.1 小节中吡虫啉施用 2 天后的小白菜地上部样品开展生理活性分析,样品涵盖 10 mg/L、100 mg/L 吡虫啉处理组及对照组。采用试剂盒并严格按照制造商操作说明,测定丙二醛(MDA)、总蛋白、细胞色素 P450 的含量,以及过氧化物酶、超氧化物歧化酶的活性⁽¹⁹⁾。
2.3 代谢组学分析
对 2.1.2 小节所述的根系分泌物溶液,采用德国 Christ 公司的 ALPHA 1-2 LD plus 型真空冷冻干燥机进行冷冻干燥处理。称取 0.50 g 干燥后的根系分泌物样品,与 2.1.1 小节指定的小白菜地上部样品混合,加入 3 mL 80%(体积体积比)甲醇溶液进行匀浆处理;涡旋振荡 10 分钟后,在 3802×g 离心力下继续离心 10 分钟,取上清液经 0.22 μm 滤膜过滤后,采用岛津 LC-20A 液相色谱仪联用 AB Sciex TripleTOF 5600 + 型四极杆飞行时间质谱仪进行检测分析。
质谱数据采用电喷雾双喷离子源,以信息依赖性采集(IDA)模式进行采集;色谱分离采用沃特世 XSelect HSS T3 色谱柱(规格:2.1 mm×100 mm,粒径 3.5 μm)。正离子模式下,流动相 A 为含 0.1% 甲酸的水溶液;负离子模式下,流动相 A 为 5 mmol/L 乙酸铵溶液。流动相 B 为乙腈,梯度洗脱程序设置如下:0~3.0 分钟,流动相 B 占比 1%;3.1~24.0 分钟,流动相 B 占比由 1% 线性升至 100%;24.1~32.0 分钟,流动相 B 占比保持 100%;32.1~37.0 分钟,流动相 B 占比恢复至 1%。仪器的其他操作参数参考本团队前期研究结果⁽²⁾。
采用 MS-DIAL 软件提取质谱峰信息,包括一、二级质谱数据、峰面积及保留时间。通过主成分分析(PCA)结合置换多元方差分析(PERMANOVA)对代谢谱进行统计分析,同时采用偏最小二乘判别分析(PLS-DA,变量投影重要性 > 1)和 t 检验(P<0.05)筛选差异显著的质谱峰。将一、二级质谱数据及保留时间与公共数据库中的条目进行匹配,完成代谢物鉴定,部分代谢物还通过化学标准品进行了验证⁽²⁰⁾。采用斯皮尔曼等级相关分析探究代谢物与吡虫啉施用量之间的关联,筛选出相关系数绝对值> 0.7 且 P<0.05 的显著性相关关系。
2.4 细菌分析
采用美国佐治亚州诺克罗斯市欧米茄生物科技公司的 E.Z.N.A. 土壤 DNA 提取试剂盒,从 2.1.1 小节所述的根际土壤样品中提取 DNA⁽²¹⁾。利用通用细菌引物,通过定量聚合酶链式反应(qPCR)测定细菌基因拷贝数,正向引物序列为 5′-CCTACGGGAGGCAGCAG-3′,反向引物序列为 5′-ATTACCGCGGCTGCTGG-3′。通过 PCR 扩增细菌 16S 核糖体 RNA(16S rRNA)基因的 V3~V4 高变区,随后采用美国加利福尼亚州圣地亚哥市因美纳公司的 Illumina MiSeq 测序平台对扩增产物进行测序。利用 DADA2 算法对扩增子序列进行去噪处理,获得扩增子序列变异体(ASVs),后续采用 mothur 软件进行分析。
采用 mothur 软件计算 Chao1 指数和 Sobs 指数这两项 α 多样性指数。通过主坐标分析(PCoA)分析不同处理组在扩增子序列变异体(ASV)水平上的细菌群落结构差异。借助基于 SILVA 138 数据库的 Classify-sklearn 朴素贝叶斯分类器,对各 16S rRNA 基因进行物种分类注释。利用线性判别分析效应量法(LEfSe)对比吡虫啉处理组与对照组的菌属差异,仅将 P<0.05 且线性判别分析(LDA)效应量绝对值>2 的类群判定为差异显著类群⁽²²⁾。开展斯皮尔曼等级相关分析,探究代谢物与菌属间的关联关系,分析前先将代谢物的相对丰度和菌属的丰度值均标准化为百万分比(ppm)单位,再进行以 10 为底的对数转换;仅将相关系数绝对值 | r|>0.7 且 P<0.05 的关联判定为具有统计学显著性的相关关系。此外,还采用基于京都基因与基因组百科全书(KEGG)数据库的 PICRUSt2 方法开展后续分析。
2.5 改变后的细菌群落的反馈效应
2.5.1 植株生长及吡虫啉在植株地上部的降解
取对照组、10 mg/L 和 100 mg/L 吡虫啉处理组的栽培土壤(参见 2.1.1 小节),用于小白菜幼苗的重植试验。种植前,选取部分 10 mg/L 吡虫啉处理组的土壤,委托中国南京希悦科技有限公司采用 γ 射线辐照法进行灭菌处理⁽²⁴⁾。幼苗培养 20 天后,测定植株生物量、地上部叶绿素及总蛋白含量,同时测定根际土壤中的脱氢酶、脲酶、过氧化物酶活性,以及速效钾和有效磷含量。对剩余的每株幼苗,叶面喷施 2.0 mL 浓度为 10 mg/L 的吡虫啉水溶液,喷施前用铝箔遮盖花盆口。继续培养 10 天后,采集植株地上部样品,测定吡虫啉和细胞色素 P450 的含量,并检测过氧化物酶与超氧化物歧化酶的活性。
2.5.2 吡虫啉的体外降解试验
称取 2.1 小节所述对照组、10 mg/L 和 100 mg/L 吡虫啉处理组的根际土壤样品各 5 g,将其重悬于 20 mL 磷酸盐缓冲盐溶液中。将该悬液置于 37℃条件下,以 150 转 / 分钟的转速振荡培养半小时。随后在 3000×g 离心力下离心 10 分钟,小心吸取所得上清液,转移至装有 50 mL 1/20 浓度胰蛋白胨大豆肉汤培养基的三角瓶中,该培养基添加了 5 mg/L 吡虫啉。经稀释的胰蛋白胨大豆肉汤培养基可提供低浓度的必需营养物质,以此提高整个根际微生物群落的生物量。将混合体系继续培养 5 天后,收集培养液用于吡虫啉含量检测。
2.6 吡虫啉及其降解产物的检测
称取 0.50 g 小白菜组织或根际土壤样品,加入 4 mL 乙腈进行提取,涡旋混合 10 分钟后,在 3802×g 离心力下离心 10 分钟。若为小白菜组织样品,取 2 mL 上清液,加入含 50 mg 石墨化炭黑和 150 mg 硫酸镁的混合试剂进行净化,将所得溶液涡旋 2 分钟后,在相同离心条件下再次离心。随后,将小白菜组织和根际土壤样品的上清液均经 0.22 μm 滤膜过滤,再进行高效液相色谱(HPLC)检测。对于培养液样品,取 2 mL 待测液,与等体积乙腈混合,涡旋 10 分钟后经 0.22 μm 滤膜过滤,再进行高效液相色谱检测。本研究的色谱检测采用安捷伦 1260 高效液相色谱仪,配备二极管阵列检测器,色谱柱为安捷伦 ZORBAX SB-C18 分析柱(规格:4.6 mm×250 mm,粒径 5 μm)。流动相为含 30% 乙腈的水溶液,流速设置为 0.5 mL/min。以信噪比的 10 倍计算定量限(LOQ),该方法检测培养液中吡虫啉的定量限为 0.03 mg/L,检测小白菜组织和根际土壤中吡虫啉的定量限为 0.04 mg/kg。此外,针对吡虫啉的两种重要降解产物 ——5 - 羟基吡虫啉和烯烃吡虫啉(均为分析纯,购自美国密苏里州圣路易斯市西格玛 - 奥德里奇公司),采用安捷伦 6410 三重四极杆质谱仪联用安捷伦 1260 高效液相色谱仪进行检测,色谱柱为 ZORBAX SB-C18 分析柱(规格:2.1 mm×100 mm)。
本研究采用土壤培养和水培培养双体系对比,结合生理、代谢组、微生物组和农药检测技术,辅以重植和体外降解验证实验,具体关键参数如下表:
| | |
|---|
| | 土壤组成为砂 30%、黏土 53%、粉砂 17%,有机质 2%,pH6.1;培养条件 28℃、光照 190μmol m⁻²s⁻¹、湿度 68% |
| | |
| | IMI 浓度 10/20/50/100mg/L,每株 2.0mL 含 0.1% 吐温 - 80 的水溶液,铝箔遮盖花盆防止农药入土壤 |
| | 测定 MDA、叶绿素、总蛋白、细胞色素 P450 及过氧化物酶(POD)、超氧化物歧化酶(SOD)活性 |
| | 采用 LC-QTOF/MS 检测,PCA/PLS-DA 筛选差异代谢物(VIP>1、P<0.05) |
| | 16S rRNA V3-V4 区测序,qPCR 检测细菌基因拷贝数,LEfSe 筛选差异菌属(P<0.05、LDA>2) |
| | HPLC 检测 IMI(LOQ:培养基 0.03mg/L、植株 / 土壤 0.04mg/kg),LC-MS 检测降解产物 5 - 羟基 IMI(LOQ0.5μg/kg)、烯烃 IMI(LOQ0.3μg/kg) |
| | 采用对照组、10/100mg/L IMI 处理组土壤及 10mg/L IMI 组灭菌土壤重植小白菜 |
| | 根际土壤悬液在含 5mg/L IMI 的 1/20TSB 培养基中 37℃、150rpm 培养 5 天 |
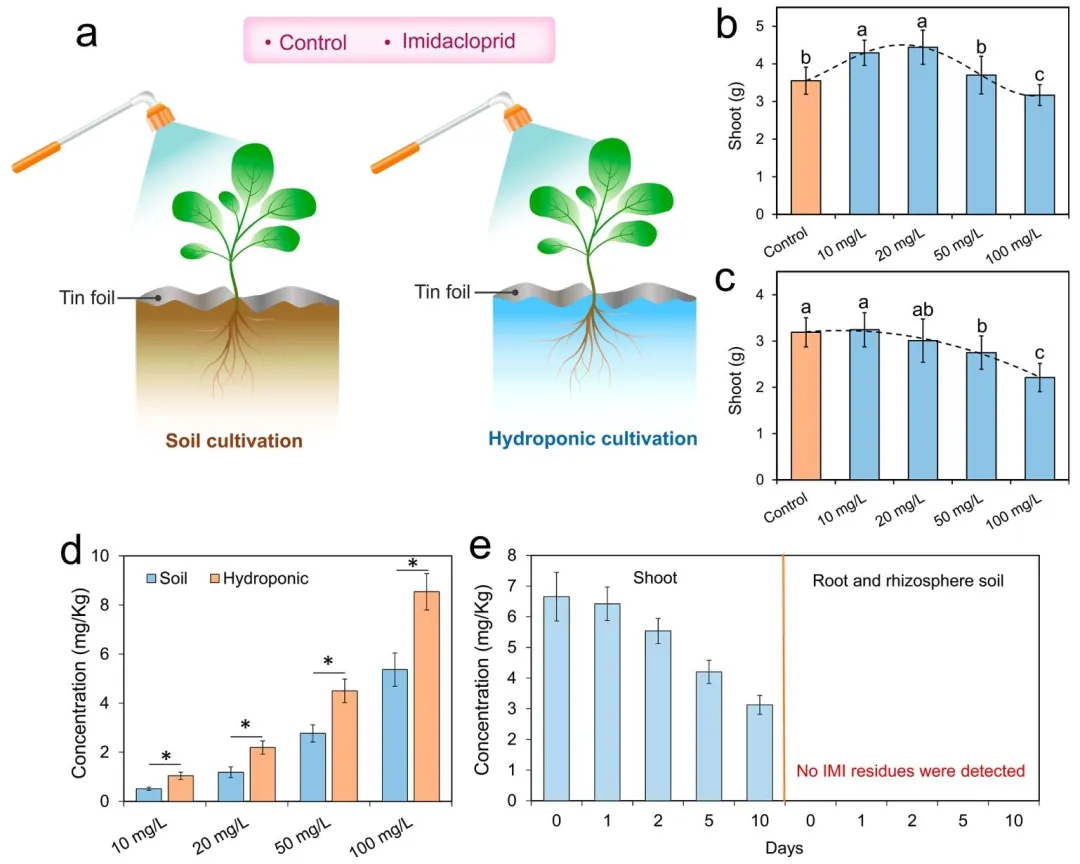
3.1 土壤培养体系提升植株对叶面喷施吡虫啉的抗性
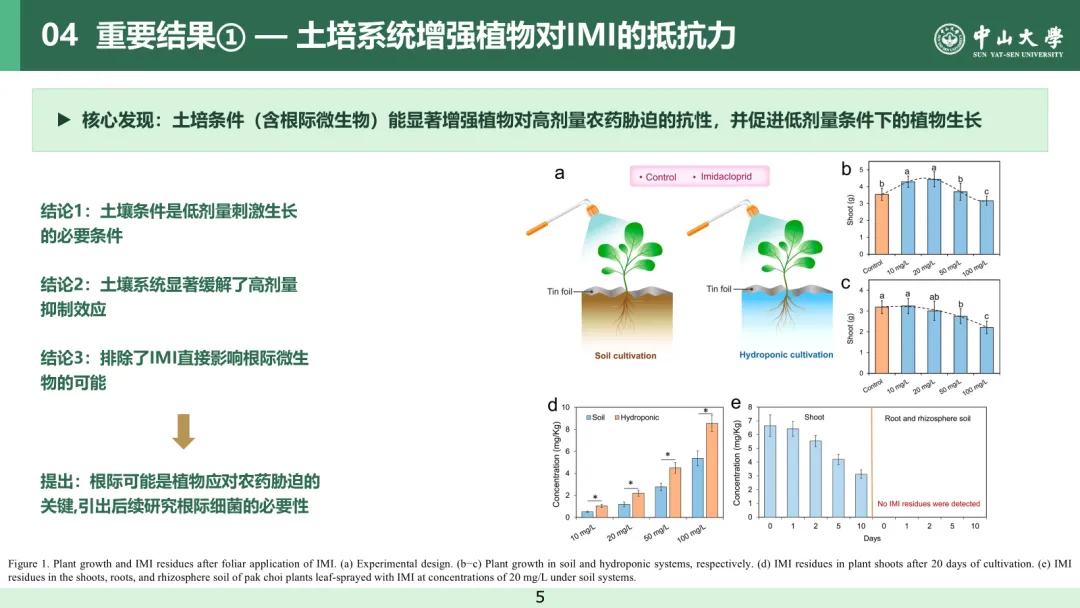
本研究分别在土壤培养和水培体系中,探究了叶面喷施吡虫啉对植株生长的影响。经 20 天培养后,所有植株均生长良好,土壤培养和水培体系中对照组植株的叶片叶绿素含量无显著差异(t 检验,P > 0.05,图 S1)。在土壤培养体系中,10 mg/L 和 20 mg/L 吡虫啉处理组的植株生物量较对照组分别显著提升约 1.22 倍和 1.26 倍(方差分析结合杜凯检验,P < 0.05,图 1b);而在水培体系中,吡虫啉处理组的植株生物量与对照组相比无显著差异(方差分析结合杜凯检验,P > 0.05,图 1c)。这表明土壤环境是低剂量吡虫啉实现植株生长促进效应的关键条件。
在 100 mg/L 高剂量吡虫啉处理下,水培和土壤培养体系中的植株生长均较对照组受到抑制(方差分析结合杜凯检验,P < 0.05),说明吡虫啉对植株生长产生了胁迫作用。值得注意的是,吡虫啉对土壤培养植株的抑制效应显著弱于水培植株,二者生物量分别为对照组的 0.89 倍和 0.70 倍(方差分析结合杜凯检验,P < 0.05),这表明与水培体系相比,土壤培养体系能增强植株对高剂量农药胁迫的抗性。此外,与水培体系相比,土壤培养体系还能促进吡虫啉在蔬菜地上部的降解,相同吡虫啉处理条件下,土壤培养体系中蔬菜地上部的吡虫啉残留量降至水培体系的 0.58~0.68 倍(t 检验,P < 0.05,图 1d)。培养期间,在蔬菜根系和根际土壤中均未检测到吡虫啉及其降解产物 5 - 羟基吡虫啉、烯烃吡虫啉的残留,这基本排除了叶面喷施吡虫啉对根际环境产生直接刺激的可能性(图 1e、图 S2)。上述结果表明,吡虫啉对蔬菜地上部的剂量依赖性胁迫与土壤环境之间存在潜在关联,进而影响植株的生长状况与抗逆能力。土壤培养和水培体系中植株的生长差异。其核心原因在于根际环境的差异。与水培体系相比,土壤培养体系的根际环境中富集着多样的微生物群落。根际微生物组常被称作植物的 “第二基因组”,在维持植株健康方面发挥着关键作用⁽²⁶⁾。据此,本研究提出假说:根际微生物在调控植株对农药胁迫的适应性过程中具有重要作用。
图 1 叶面喷施吡虫啉后植株的生长状况及吡虫啉残留分布(a) 试验设计;(b)~(c) 分别为土壤培养体系、水培体系下的植株生长情况;(d) 培养 20 天后植株地上部的吡虫啉残留量;(e) 土壤培养体系下,经 20 mg/L 吡虫啉叶面喷施处理的小白菜其地上部、根系及根际土壤中的吡虫啉残留量。柱形图上不同字母表示组间存在统计学显著差异(方差分析结合杜凯检验,P < 0.05)。植株生长指标测定设置 20 次重复,吡虫啉残留量测定设置 6 次重复,误差棒代表标准差。
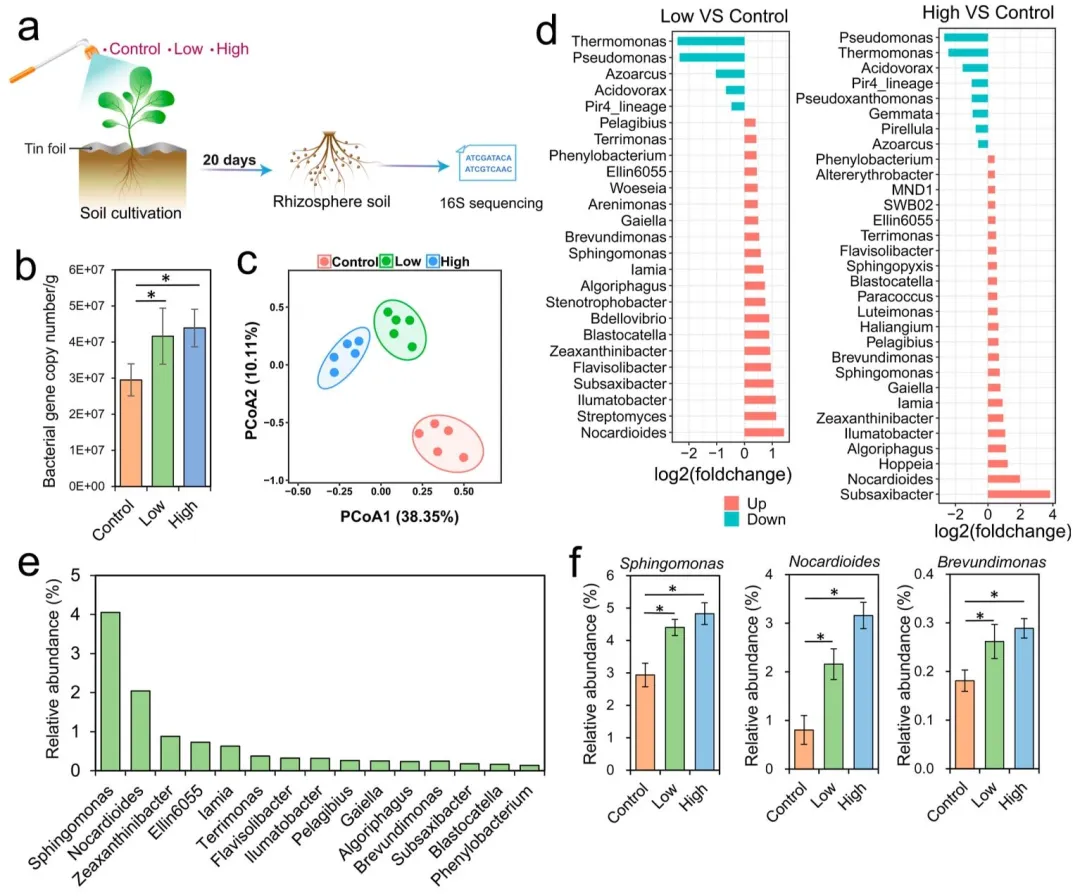
3.2 叶面喷施吡虫啉重塑植物根际细菌群落结构
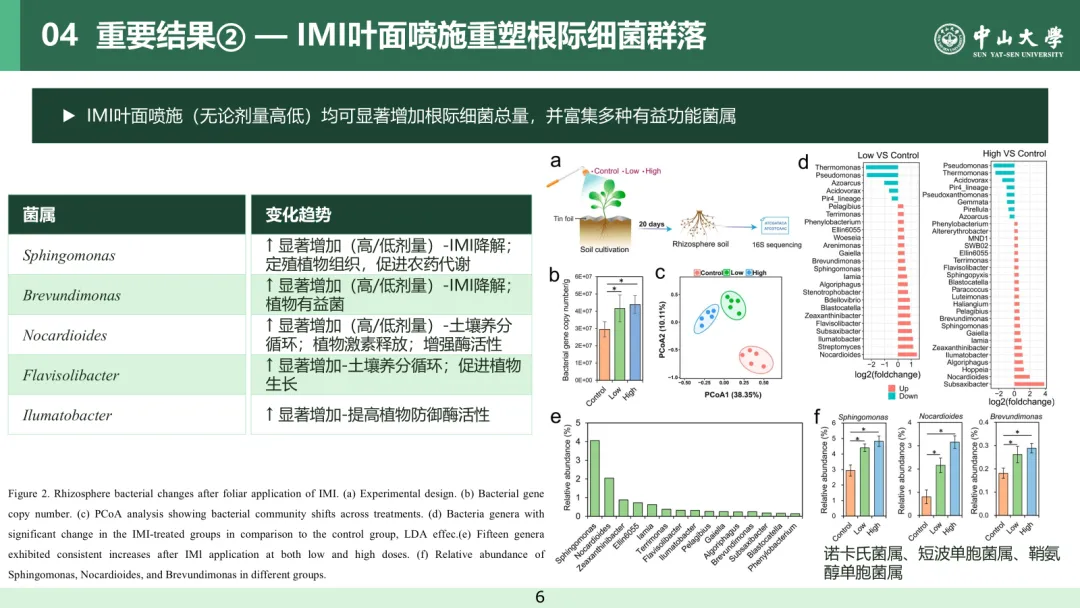
本研究进一步采用 16S rRNA 扩增子测序技术,探究了叶面喷施吡虫啉对根际土壤中细菌数量及群落结构的影响(图 2a)。与对照组相比,10 mg/L(低剂量)和 100 mg/L(高剂量)吡虫啉处理组的细菌基因拷贝数分别提升 1.41 倍和 1.49 倍(威尔科克森秩和检验,P<0.05,图 2b)。从根际土壤样品中共鉴定得到 1698648 条细菌序列,包含 3117 条细菌扩增子序列变异体(ASV)序列,以及 364 个共有菌属。主坐标分析(PCoA)结果显示,叶面喷施吡虫啉可显著影响根际细菌群落的组成结构(置换多元方差分析,P<0.05,图 2c)。吡虫啉处理后,根际细菌的 Sobs 指数和 Chao1 指数均显著高于对照组(威尔科克森秩和检验,P<0.05,图 S3),共现网络分析结果则表明其细菌群落的连通性和复杂程度均有所提升(图 S4)。在 76 个平均相对丰度大于 0.1% 的共有菌属中,有 36 个菌属在吡虫啉处理组与对照组间存在显著差异(线性判别分析效应量法,P<0.05,线性判别分析效应量绝对值> 2,表 S1)。其中,低剂量吡虫啉处理组有 20 个菌属显著富集、5 个菌属显著减少;高剂量吡虫啉处理组有 23 个菌属显著富集、8 个菌属显著减少(图 2d)。此外,有 15 个菌属在低、高剂量吡虫啉处理后均呈现显著富集的趋势,包括诺卡氏菌属、黄溶杆菌属、突柄杆菌属、鞘氨醇单胞菌属和短波单胞菌属等(威尔科克森秩和检验,P<0.05,图 3e-f)。
已有部分研究表明,污染土壤中的农药会对土壤微生物产生不利甚至严重的影响⁽²⁷⁾,而本研究结果显示,遭受叶面喷施吡虫啉胁迫的蔬菜植株,其根际细菌的数量和群落多样性均有所提升。造成该差异的主要原因是,本研究的实验体系中无农药残留在土壤中,排除了农药对根际微生物产生直接作用的可能性。在叶面喷施吡虫啉的胁迫下,土壤中多种土著细菌在植物根际显著富集,且其中多个菌属已被证实具有促植物生长的功能。例如,鞘氨醇单胞菌属和短波单胞菌属的细菌具备吡虫啉降解潜力⁽²⁸⁾⁽²⁹⁾,这类细菌还能定殖于植物组织内部,进而促进吡虫啉在植物体内的降解⁽³⁰⁾;诺卡氏菌属、黄溶杆菌属、突柄杆菌属等其他有益菌,则可参与土壤养分循环、促进植物激素合成释放,并提升植物防御酶的活性⁽³¹⁾⁽³²⁾。上述结果表明,蔬菜植株在遭受吡虫啉这一非生物胁迫时,能够特异性地招募多种有益细菌,而这些细菌在调控植株生长和提升抗逆能力方面具有潜在作用。
图 2 叶面喷施吡虫啉后根际细菌的变化特征。(a) 试验设计;(b) 细菌基因拷贝数;(c) 主坐标分析(PCoA)展示不同处理组的细菌群落结构变化;(d) 吡虫啉处理组与对照组间存在显著差异的菌属(线性判别分析效应量法,P<0.05,线性判别分析效应量> 2);(e) 低、高剂量吡虫啉处理后均呈显著富集的 15 个菌属;(f) 不同处理组中鞘氨醇单胞菌属、诺卡氏菌属和短波单胞菌属的相对丰度。星号表示组间存在统计学显著差异(威尔科克森秩和检验,P<0.05)。柱形代表 5 次重复的平均值,误差棒代表标准差。
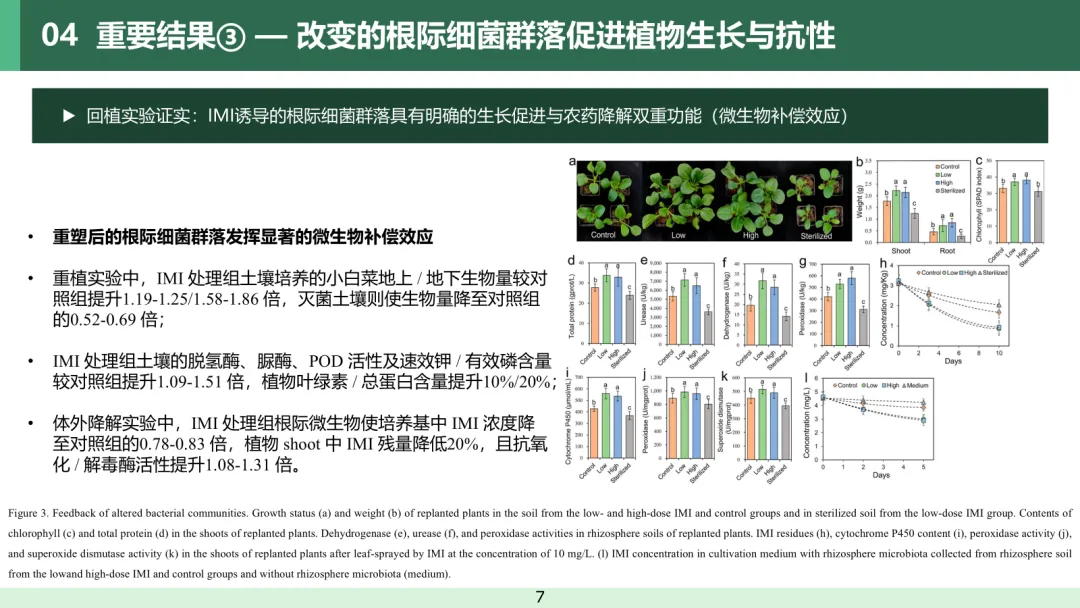
3.3 根际细菌群落的改变促进植株生长并增强抗逆性
为探究吡虫啉诱导形成的根际细菌群落对植株生长的影响,本研究将小白菜幼苗重植于取自低剂量、高剂量吡虫啉处理组及对照组的土壤,同时设置低剂量吡虫啉处理组的土壤灭菌处理作为对照。培养 2 周后,种植于吡虫啉处理组土壤中的植株,其生长状况显著优于对照组土壤中的植株,地上部鲜重与根系鲜重分别为对照组的 1.19~1.25 倍和 1.58~1.86 倍(方差分析,杜凯检验,P<0.05;图 3a−b)。与之相反,种植于吡虫啉处理组灭菌土壤中的植株生长受到显著抑制,生物量仅为另外三组的 0.52~0.69 倍(方差分析,杜凯检验,P<0.05;图 3a−b)。吡虫啉处理组土壤中生长的植株,其地上部叶绿素与总蛋白含量显著提升,分别比对照组土壤植株高约 10% 和 20%(方差分析,杜凯检验,P<0.05;图 3c−d)。同时,该组植株根际土壤中的脱氢酶、脲酶、过氧化物酶活性以及速效钾、有效磷含量也呈现相似的上升趋势,较对照组提升 1.09~1.51 倍(方差分析,杜凯检验,P<0.05;图 3e−g、图 S5)。
对种植于对照组、吡虫啉处理组及灭菌土壤中的小白菜统一喷施 10 mg/L 吡虫啉溶液。10 天后,吡虫啉处理组土壤中植株地上部的吡虫啉含量较对照组与灭菌土壤组低约 20%(方差分析,杜凯检验,P<0.05;图 3h)。此外,该组植株体内的解毒酶(细胞色素 P450)与抗氧化酶(过氧化物酶、超氧化物歧化酶)活性显著升高,较对照组提升 1.08~1.31 倍(方差分析,杜凯检验,P<0.05;图 3i−k)。本研究收集了吡虫啉处理组与对照组根际土壤中的微生物,开展吡虫啉体外降解试验。培养 5 天后,吡虫啉处理组的根际微生物可显著提升培养液中吡虫啉的降解效率,其吡虫啉浓度仅为对照组的 0.78~0.83 倍(方差分析,杜凯检验,P<0.05;图 3)。现有研究表明,农药可诱导植株产生氧化胁迫⁽³³⁾并改变根际微生物群落组成⁽³⁴⁻³⁵⁾,但此类微生物群落变化对植株生长与健康的反馈作用尚不明确。本研究证实,叶面喷施吡虫啉后重塑的根际细菌群落,不仅能通过降解植株体内的农药有效缓解胁迫,还可发挥促生功能,增强植株抗逆性,这类有益效应可定义为微生物补偿作用。该结果与植株从土壤中招募特定微生物以抵御病原菌或昆虫引发的生物胁迫这一策略相一致⁽³⁶⁻³⁷⁾。值得注意的是,低、高剂量吡虫啉诱导形成的细菌群落,在促进植株生长、提升酶活性及降解吡虫啉等方面未表现出显著差异,表明吡虫啉可作为一种 “开关”,激活根际微生物的补偿效应,以抵御农药对植株造成的胁迫。上述研究结果为深入理解将根际微生物作为生态友好型手段缓解作物农药胁迫提供了理论依据。
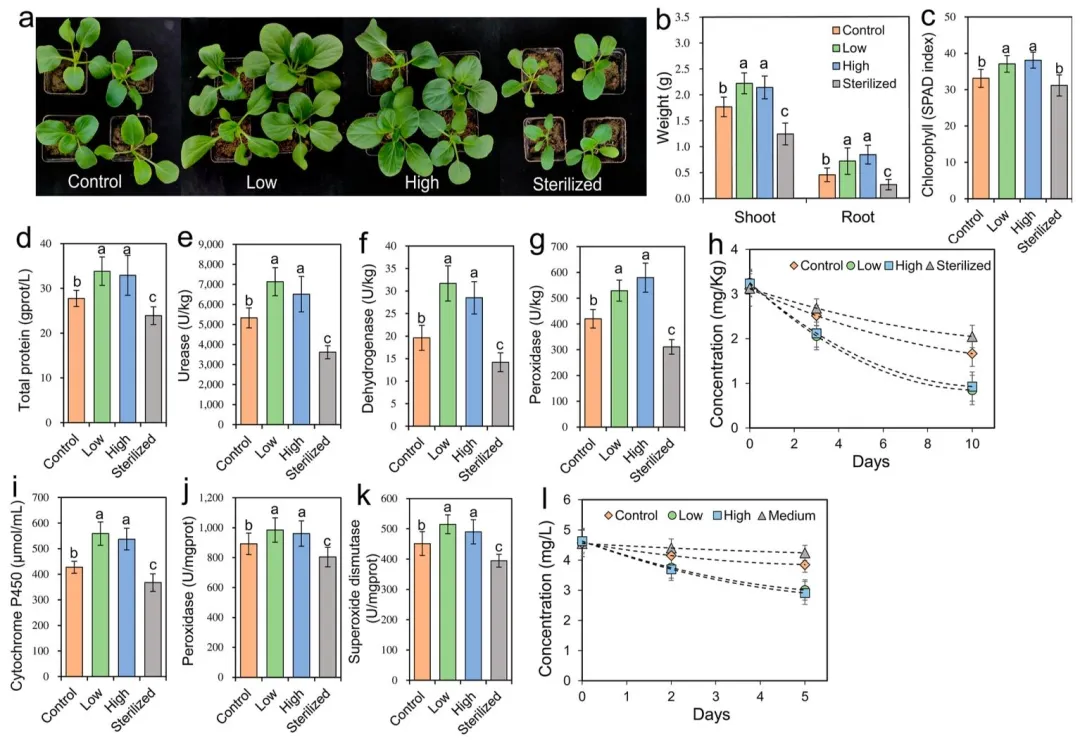
图 3 根际细菌群落结构改变后的反馈效应。(a) 重植植株的生长状况、(b) 重植植株的生物量(土壤分别取自低 / 高剂量吡虫啉处理组、对照组及低剂量吡虫啉处理组的灭菌土壤);(c) 重植植株地上部的叶绿素含量、(d) 重植植株地上部的总蛋白含量;重植植株根际土壤中的 (e) 脱氢酶活性、(f) 脲酶活性、(g) 过氧化物酶活性;经 10 mg/L 吡虫啉叶面喷施后,重植植株地上部的 (h) 吡虫啉残留量、(i) 细胞色素 P450 含量、(j) 过氧化物酶活性、(k) 超氧化物歧化酶活性;(l) 含低 / 高剂量吡虫啉处理组、对照组根际微生物的培养基及无添加根际微生物的空白培养基中的吡虫啉浓度。柱形图上不同字母表示组间存在统计学显著差异(方差分析结合杜凯检验,P<0.05)。叶绿素含量测定设置 20 次重复,其余指标测定均设置 6 次重复,误差棒代表标准差。
3.4 叶面喷施吡虫啉改变植株根系分泌物代谢谱
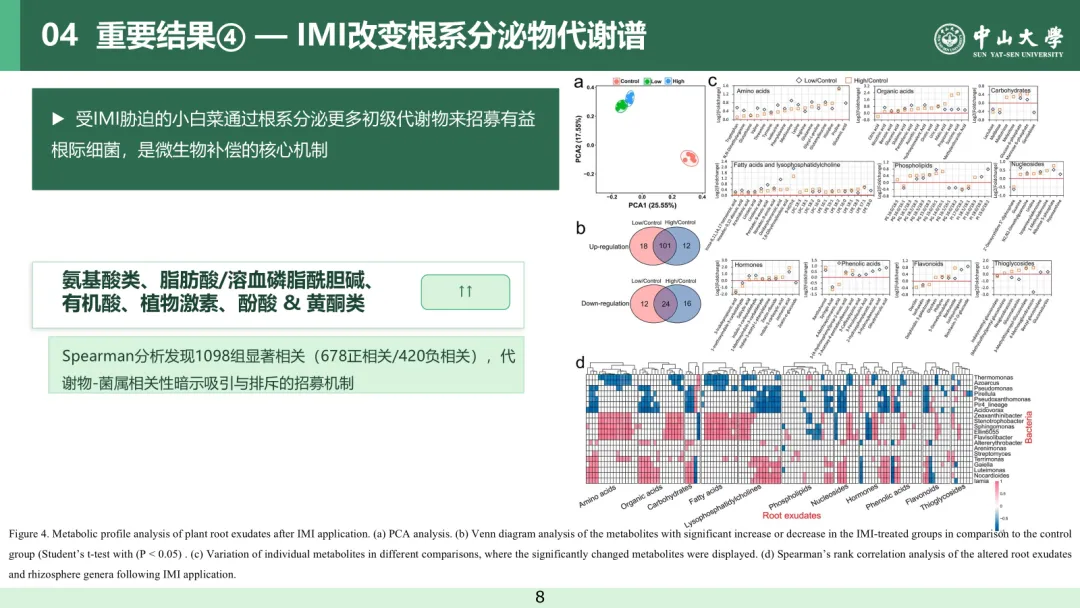
采用代谢组学技术分析叶面喷施吡虫啉对植物根系分泌物释放的影响。主成分分析(PCA)结果表明,叶面喷施吡虫啉可显著改变植株根系分泌物的代谢谱特征(置换多元方差分析,P < 0.05;图 4a)。与对照组相比,吡虫啉处理组共有 160 种根系分泌物代谢物发生显著变化,其中 101 种代谢物在低、高剂量处理下均显著上调,24 种均显著下调(t检验,P<0.05;偏最小二乘判别分析,变量投影重要度 VIP>1;表 S2、图 S6、图 4b)。在这些差异代谢物中,上调物质以初级代谢产物为主,包括氨基酸、有机酸、脂肪酸、溶血磷脂酰胆碱以及糖磷酸酯等,其含量变化呈现吡虫啉剂量依赖性(t检验,P < 0.05;偏最小二乘判别分析,VIP>1;图 4c、图 S7)。茉莉酸、水杨酸、玉米素核苷、吲哚 - 3 - 甲醛等植物激素在吡虫啉处理后均显著升高(t检验,P<0.05;偏最小二乘判别分析,VIP>1)。与对照组相比,低剂量吡虫啉处理组还富集了大量酚酸和类黄酮物质(t检验,P<0.05;偏最小二乘判别分析,VIP>1;图 4c)。新葡萄糖芸苔素、葡萄糖芥苷、4 - 甲氧基葡萄糖芸苔素等硫代糖苷类物质,在低剂量吡虫啉处理下含量降低,而在高剂量处理下含量升高(t检验,P < 0.05;偏最小二乘判别分析,VIP>1;图 4c)。本研究结果表明,蔬菜植株在应对吡虫啉胁迫时,会促进根系分泌物中多种初级代谢物的释放。吡虫啉对植物根系分泌物的影响模式与病原菌侵染较为相似⁽³⁷⁾,但与养分缺乏胁迫存在显著差异——后者仅会引起香豆素和酚类物质含量上升⁽²⁰,³⁸⁾。值得注意的是,根系分泌物主要由光合产物组成,经由根系分泌至根际环境中⁽³⁹⁾。这意味着在农药胁迫下,植物可能通过向根际释放更多光合产物来做出适应性响应。吡虫啉处理后,上述根系分泌物代谢物的变化可从多个方面对根际环境产生调控作用。例如,氨基酸和小分子有机酸已被证实能够影响土壤结构、养分循环以及植物—微生物互作关系⁽⁴⁰⁾;硫代葡萄糖苷与酚酸类物质均能够抑制病原微生物的生长繁殖⁽⁴¹,⁴²⁾。本研究结果揭示了农药可对根系分泌物变化产生潜在影响,有助于更深入地理解农药对根际内植物互作关系的作用机制。
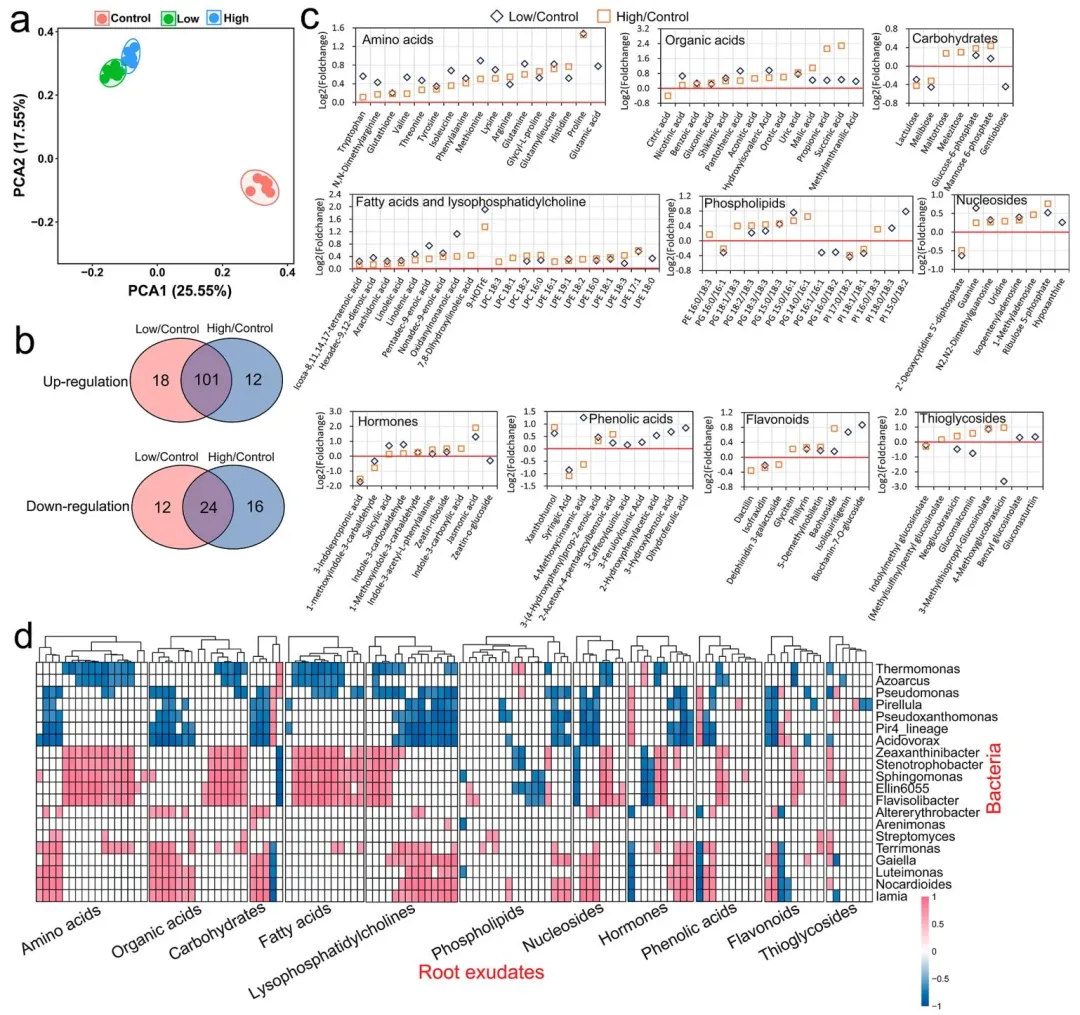
图 4 吡虫啉处理后植株根系分泌物代谢谱分析。(a) 主成分分析(PCA);(b) 吡虫啉处理组与对照组相比显著上调或下调代谢物的韦恩图分析(t 检验,P<0.05;偏最小二乘判别分析,VIP>1);(c) 不同比较组中单个代谢物的变化情况,图中展示差异显著的代谢物;(d) 吡虫啉处理后差异根系分泌物与根际菌属的斯皮尔曼等级相关分析(斯皮尔曼相关,|r|>0.7,P<0.05)。每组设置 5 次重复,误差棒表示标准差。
3.5 根系分泌物可驱动根际细菌群落及功能的改变
吡虫啉处理后,差异根系分泌物与根际菌属之间共鉴定出1098 条显著相关关系,其中正相关 678 条、负相关 420 条(斯皮尔曼等级相关,|r|>0.7,P<0.05;图 4d)。在这些关联中,溶血磷脂酰胆碱、有机酸、氨基酸、糖类及植物激素类代谢物与菌属的显著相关数量最多,每类物质平均关联 8.4~12.3 个菌属。诺卡氏菌属、浮游杆菌属、黄溶杆菌属、嗜冷杆菌属、伊利亚菌属、鞘氨醇单胞菌属和热单胞菌属与根系分泌物代谢物的显著相关数量最多,每个菌属平均关联 33~48 种代谢物。此外,本研究发现不同类型的根系分泌物代谢物倾向于调控特定的菌属。芳香族氨基酸(酪氨酸、色氨酸、苯丙氨酸)和酚酸类物质(二氢阿魏酸、3 - 羟基苯甲酸、4 - 甲氧基肉桂酸)与黄溶杆菌属呈显著正相关(斯皮尔曼等级相关,P<0.05)。低分子量有机酸(乌头酸、琥珀酸、丙酸、苹果酸)、18 碳脂肪酸相关脂类(磷脂酰甘油、溶血磷脂酰胆碱、溶血磷脂酰乙醇胺)以及糖磷酸酯(6 - 磷酸甘露糖、6 - 磷酸葡萄糖)与诺卡氏菌属和盖勒氏菌属呈显著正相关(斯皮尔曼等级相关,P<0.05)。类黄酮物质异嗪皮啶与热单胞菌属呈正相关,却与诺卡氏菌属呈负相关(斯皮尔曼等级相关,P<0.05)。基于 KEGG 数据库的 PICRUSt2 功能预测分析显示,与对照组相比,低、高剂量吡虫啉处理组中与氨基酸、碳水化合物及脂类代谢相关的细菌功能均显著上调(LEfSe,P<0.05)。这些细菌功能与根系分泌物中对应类别的代谢物呈显著正相关(斯皮尔曼等级相关,P<0.05;图 S8a−b)。与环境信息处理相关的功能,如膜转运、信号转导和环境适应等,在吡虫啉处理组中也较对照组显著提升(LEfSe,P<0.05;图 S8c)。上述结果表明,小白菜植株在应对吡虫啉胁迫时,可通过改变根系分泌物来调控根际细菌群落结构及其功能。通常,根系分泌物代谢物可通过影响根际微生物的生长生态位、运动能力及生物膜形成等方式调控微生物群落结构⁽⁴³,⁴⁴⁾。吡虫啉胁迫下根系分泌物代谢物与根际菌属之间的正负相关关系,可能分别对应微生物的招募富集与排斥抑制。这些关联关系仍有待后续试验进一步验证。本研究结果有助于更深入地理解农药胁迫下根际环境的动态变化机制。
图 5 吡虫啉处理后植株生理代谢变化。(a)、(b) 分别为蔬菜地上部丙二醛(MDA)含量和叶绿素含量。柱形图上不同字母表示组间差异具有统计学意义(方差分析,杜凯检验,P<0.05)。(c) 植株地上部代谢谱的主成分分析(PCA)。(d) 吡虫啉处理组与对照组相比显著上调或下调代谢物的韦恩图分析(t 检验,P<0.05;偏最小二乘判别分析,VIP>1)。(e) 不同比较组中单个代谢物的变化情况。叶绿素含量测定设置 20 次重复,其余指标均设置 6 次重复。误差棒代表标准差。
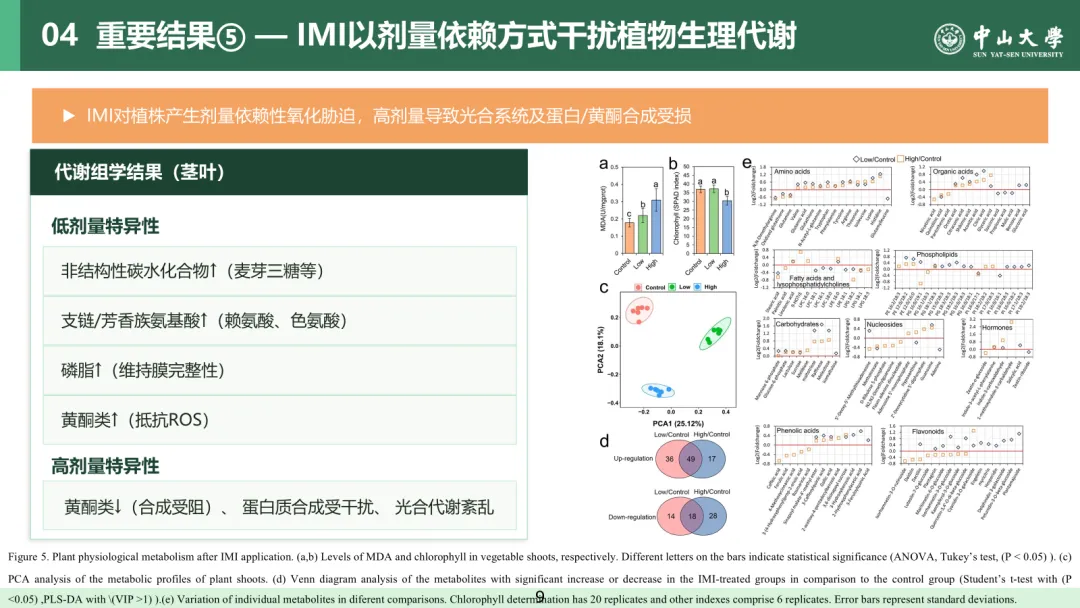
3.6 吡虫啉以剂量依赖方式干扰植株生理代谢
以往研究多关注农药种类对作物生理代谢的影响,却忽视了农药施用剂量的作用⁽¹⁾。本研究测定了不同剂量叶面喷施吡虫啉对小白菜地上部生理代谢的影响。植株地上部丙二醛(MDA,膜脂过氧化指标)含量以及过氧化物酶、超氧化物歧化酶活性均随吡虫啉施用剂量升高而显著上升(斯皮尔曼等级相关,r=0.568~0.718,P<0.05;图 5a、图 S9),表明吡虫啉可对小白菜植株产生剂量依赖性氧化胁迫。此外,与对照组相比,高剂量吡虫啉处理使植株蛋白和叶绿素含量分别显著下降 11.2% 和 15.4%(方差分析,杜凯检验,P<0.05;图 5b、图 S9)。基于液相色谱 - 四极杆飞行时间质谱(LC-QTOF/MS)的代谢组学结果显示,吡虫啉可扰乱小白菜地上部的代谢谱(主成分分析,置换多元方差分析,P<0.05;图 5c)。与对照组相比,低、高剂量吡虫啉处理组分别有 85 种和 66 种代谢物显著上调,同时分别有 28 种和 46 种代谢物显著下调(t检验,P<0.05;偏最小二乘判别分析,VIP>1;图 5d、图 S10 及表 S3)。在这些差异代谢物中,多种非结构性碳水化合物(麦芽三糖、棉子糖、松三糖等)、支链氨基酸与芳香族氨基酸(赖氨酸、色氨酸以及酪氨酸等)和小分子有机酸(莽草酸、乌头酸、柠檬酸等)在低、高剂量吡虫啉处理下均出现明显积累(t 检验,P<0.05;偏最小二乘判别分析,VIP>1;图 5e)。
此外,低剂量吡虫啉还可促进磷脂(PG 18:2/18:3、PG 18:3/18:3、PI 18:2/18:3 等)和类黄酮(海葵黄素 - 6-O - 葡萄糖苷、异鼠李素 - 3-O - 葡萄糖苷、山奈酚 - 3-O - 葡萄糖苷等)的积累,而高剂量吡虫啉则使植株地上部多种类黄酮含量下降(t 检验,P<0.05;偏最小二乘判别分析,VIP>1;图 5e)。本研究结果表明,吡虫啉可在小白菜植株中引发剂量依赖性氧化胁迫,并显著改变其代谢谱。低、高剂量吡虫啉胁迫下,植株地上部积累的碳水化合物与氨基酸可作为能量来源或渗透调节物质,以适应逆境⁽⁵⁾。同时,低剂量吡虫啉提升了与抗逆相关的磷脂和类黄酮含量,前者可维持细胞膜完整性与流动性,后者可清除活性氧⁽⁴⁵,⁴⁶⁾。高剂量吡虫啉则会对植株造成严重的氧化损伤,干扰光合作用及蛋白质、类黄酮的合成,进而可能影响植株生长发育与抗逆能力。本研究阐明了农药施用剂量与植株代谢之间的关联,为指导农业生产中农药的科学合理使用提供了重要依据。
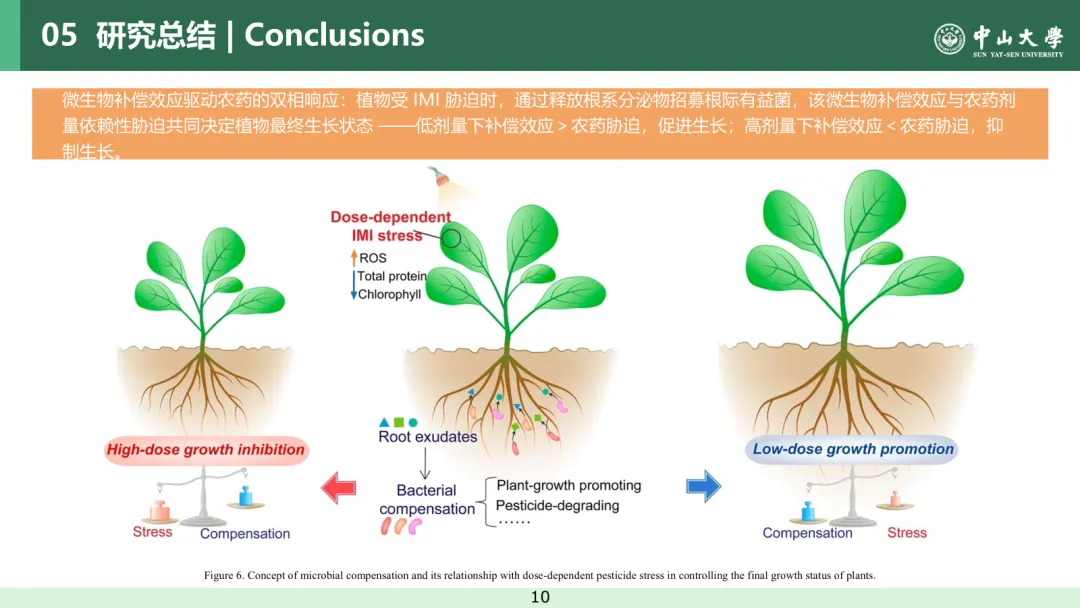
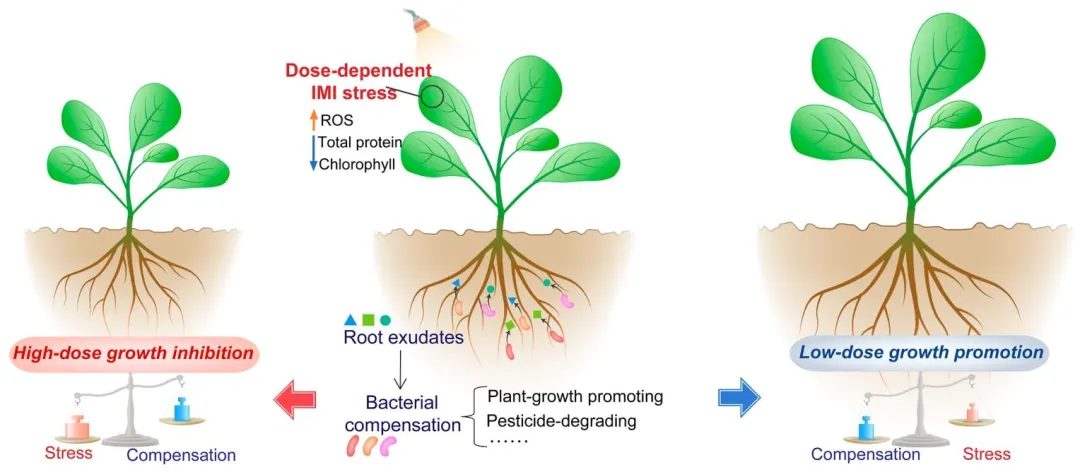
图 6 微生物补偿作用概念及其与剂量依赖性农药胁迫在调控植株最终生长状态中的关联机制。
植物在整个生长发育过程中会遭遇多种生物胁迫与非生物胁迫⁽⁴⁷⁾。尽管温度、土壤湿度、光照以及众多化学物质(农药、重金属、纳米材料、抗生素、多环芳烃等)这类环境因子在调控植物生长时常表现出双相效应⁽⁴⁸,⁴⁹⁾,但其内在机制仍不明确。
基于原文的研究空白与科学问题拓展
科学问题 1:吡虫啉诱导的关键有益菌的功能机制未通过无菌 / 回接实验验证
原文依据:“Subsequent studies are essential for isolating rhizosphere bacteria and validating their functions on plants through reintroduction experiments or sterile seedling systems.”
核心未解决点:仅通过相关性证明 Sphingomonas、Nocardioides 等富集,但是否真正降解农药、促生长、提升抗性缺乏直接因果证据。
科学问题 2:土壤残留农药(非叶面喷施)对植物 — 微生物互作的影响仍未研究
原文依据:“many pesticides remained in the environment at concentrations lower than those used for foliar applications… could directly affect the rhizosphere environment… need further investigation.”
核心未解决点:本研究仅叶面喷施;土壤低剂量长期残留农药是否通过根系吸收改变互作模式尚不明确。
科学问题 3:植物内生菌在农药胁迫响应中的作用未被纳入研究
原文依据:“plants host various endophytes that could participate in the bioremediation of organic pollutants within plant tissues… Further research is needed to investigate the effects of pesticides on plant endophytic communities.”
核心未解决点:只关注根际菌群,内生菌是否协同降解农药、调控代谢、增强抗性完全空白。
文章结论
本研究结果表明,小白菜植株在应对叶面吡虫啉胁迫时,可通过改变根系分泌物代谢谱,富集根际有益微生物。经重塑的根际微生物群落能够发挥补偿作用,协助植株抵御农药胁迫。结合土壤体系中不同剂量吡虫啉胁迫下植株的生长变化,本研究提出假说:微生物补偿效应与剂量依赖性农药胁迫共同调控植株最终的生长状态(图 6)。低剂量农药处理下,微生物补偿效应强于农药胁迫,从而促进植株生长;而高剂量农药处理下,微生物补偿效应不足以抵消农药胁迫,最终导致植株生长受抑。上述研究结果有助于深入理解农药施用剂量与微生物补偿作用之间的平衡关系,对优化农药施用、降低其对非靶标生物及环境的危害具有重要意义。综上,本研究揭示了微生物补偿效应与剂量依赖性农药胁迫互作调控植株生长及抗逆性的内在机制,为相关领域研究提供了重要参考。后续研究需进一步分离根际细菌,并通过回接试验或无菌苗体系验证其对植株的功能。
值得注意的是,本实验采用农业生产中最常用的叶面喷施方式施用农药,而环境中残留的许多农药浓度往往低于叶面喷施剂量。与叶面处理不同,土壤中残留的农药可被植株从根际吸收,并直接影响土壤性质、微生物群落等根际环境⁽⁵⁰⁾。在此条件下,农药剂量 — 植株 — 根际微生物三者间的互作关系仍有待深入探究。尽管本研究聚焦于根际微生物群落,但植株体内还存在多种内生菌,可参与植物体内有机污染物的生物修复⁽⁵¹,⁵²⁾。未来需进一步研究农药对植物内生菌群落的影响,明确植株 — 内生菌互作是否能够增强植株对农药胁迫的抗性。






1)如何从文献实验方法中明确判断是靶向还是非靶向代谢组学?
需要阅读文献的《材料与方法》中关于代谢组学分析的部分(通常标题里带有 "Metabolomic analysis", "LC-MS analysis" 或类似字眼)。 如何区分:
靶向代谢组学 (Targeted Metabolomics): 核心特征:研究者在实验前就已经知道要测哪些具体的化合物(例如:20种特定的氨基酸,或者几种特定的激素)。 关键词寻找: - 文献中会明确列出目标化合物的清单。
- 使用了标准品 (Standards) 制作标准曲线进行绝对定量(例如浓度单位是 ng/g FW,而不是相对峰面积)。 - 仪器通常使用三重四极杆质谱 (QqQ-MS),并使用 MRM(多反应监测)或 SRM 模式。 一句话判断:如果文章说“我们测定了以下30种特定物质的浓度”,那就是靶向。
--- 非靶向代谢组学 (Untargeted Metabolomics / Global Metabolomics): 核心特征:研究者不知道会发现什么,目的是尽可能多地检测样本中的所有小分子,寻找差异代谢物。
关键词寻找: - 强调尽可能多地检测代谢特征 (Metabolic features)。 - 使用相对定量(比较峰面积大小,而不是具体浓度)。 - 仪器通常使用高分辨质谱,如 Q-TOF 或 Orbitrap。 - 数据处理使用 XCMS、MS-DIAL 等软件进行峰对齐和提取。 - 分析方法里会提到 PCA、PLS-DA 等多元统计分析来寻找“差异代谢物 (Differential metabolites)”。 一句话判断:如果文章说“我们检测到了2000个代谢特征,并通过数据库比对鉴定出其中300个”,那就是非靶向。
2)非靶标作物是什么?
定义:在农药试验中,“靶标作物”是你要保护、故意喷洒农药的目标(比如为了防治棉花蚜虫而喷洒棉花,棉花就是靶标作物)。非靶标作物 (Non-target crops) 是指那些不是农药预期处理对象,但可能无意中接触到农药的植物。 情境: 它们可能是生长在目标田块旁边的其他作物(受农药漂移影响),或者是与目标作物轮作的后茬作物(受土壤残留影响),甚至是田间的杂草。 在这篇文献中: 你需要看实验设计部分。研究者可能特意种植了一种作物(如玉米)作为主要研究对象,同时种植了另一种作物(如大豆)在旁边,或在处理过的土壤中种植另一种作物,用来评估吡虫啉的潜在环境风险。那个用来评估风险的“配角”作物,就是非靶标作物。
3)本次实验的MDA和P450分别表征什么,怎么测定?
IMI处理土壤回植实验和重植实验是同一个吗?MDA(丙二醛, Malondialdehyde)表征什么: 它是一种生物标志物,表征脂质过氧化程度。简单说,它反映了植物细胞膜受损程度和受到的氧化胁迫 (Oxidative stress)大小。MDA含量越高,植物受到的伤害越大。
怎么测定: 最常用方法是硫代巴比妥酸 (TBA) 比色法。提取植物组织液,加入TBA试剂,在酸性和高温下反应生成红色产物,用分光光度计测定吸光度。
---P450(细胞色素P450酶系, Cytochrome P450) 表征什么: P450是生物体内非常重要的一类解毒酶。在植物中,它参与许多外源物质(包括农药吡虫啉)的代谢和解毒过程。P450活性升高通常意味着植物正在启动自身防御机制来降解毒素。
怎么测定: - 酶活测定:提取植物蛋白液,加入特定底物,反应后测定产物生成量(常用分光光度法或荧光法)。 - 基因表达测定 (qPCR):测定编码P450酶的基因(如CYP家族)的转录水平。
--- “IMI处理土壤回植实验”和“重植实验”是同一个吗?是一回事。 在这种语境下,“重植 (Replanting)”通常就是指在已经进行过某种处理(这里是IMI处理)的土壤中再次种植作物,以评估土壤残留的影响。 确认方法:仔细核对文章实验设计流程图或文字描述,看两个词语是指向同一实验步骤:先向土壤施加IMI →(可能经过一段时间老化)→ 在该土壤中种植植物。
4)IMI降解体外实验是怎么做的,目的和意义是什么?
体外实验 (In vitro): 意思是“在玻璃器皿中”,即在试管、锥形瓶、培养皿中进行,脱离完整活体植物或土壤环境。 怎么做的(根据本文逻辑推测): - 在受控容器(如锥形瓶)中模拟降解环境。 - 反应体系:已知浓度吡虫啉(IMI) + 缓冲液 + 降解启动因子。 - “降解启动因子”通常是:特定降解菌株、土壤/植物粗酶液、纯化酶等。 - 在特定时间点取样,用 HPLC 或 LC-MS 检测IMI剩余浓度及降解产物。 目的和意义: - 分离机制:排除植物、土壤复杂干扰,确定到底是谁(哪种菌/哪种酶)在降解IMI。
- 鉴定产物:更容易富集、鉴定中间产物,阐明降解途径。
- 动力学研究:精确计算降解速率、半衰期。
- 因果验证:直接证明微生物/酶确实能降解IMI。
5)实验中喷洒吡虫啉(IMI)是单次喷洒还是多次喷洒?每个实验处理几个重复?
必须看原文《材料与方法》中“Plant materials and treatments”或“Experimental design”部分。其中看每个图的标题的下标可知道:植株生长指标测定设置 20 次重复,吡虫啉残留量测定设置 6 次重复。关于喷洒频次: 寻找 "sprayed once"、"single application"、"sprayed every 7 days" 等描述。 单次喷洒 = 急性暴露;多次喷洒 = 慢性累积暴露。 关于重复 (Replicates):寻找 "n=3"、"three biological replicates"、"performed in triplicate"。 一般至少3次生物学重复。
6)MS-DIAL 和 PLS-DA 分析叫做什么?
MS-DIAL是一款代谢组学数据处理软件。 功能:对质谱原始数据进行噪音过滤、峰识别 (Peak picking)、样本间峰对齐、初步代谢物鉴定。PLS-DA(偏最小二乘判别分析, Partial Least Squares-Discriminant Analysis) 属于多元统计分析方法**,是一种**有监督的模式识别方法**。 在代谢组学中,用于区分不同处理组,并找出导致组间差异的关键代谢物(根据VIP值筛选)。
7)图5为什么只做茎叶的代谢组学?不做根际的?茎和叶是分开做还是合并表征?意义何在? 为什么只做茎叶,不做根际?
- 吡虫啉是叶面喷施,首先作用于地上部,研究重点是地上部的吸收、转运、胁迫响应。 - 根际(土壤+根系+微生物)过于复杂,会干扰植物自身代谢信号。 - 根系/根际微生物可能已在其他图表单独分析。茎和叶是分开还是合并?
看图例或方法: - 标注为Shoots / Aerial parts→ 茎叶合并。 - 分别标注 Stem、Leaf → 分开检测。 通常代谢组为了整体表征和节约成本,多采用合并地上部。 意义: - 揭示吡虫啉对植物地上部光合、能量、氨基酸、脂质、次生代谢的影响。 - 阐明植物应对农药胁迫的解毒、抗氧化、耐受机制。
- 对于叶菜类作物,直接反映可食用部位的代谢安全与胁迫响应。
注:本文作为文献阅读分享记录和供科研人员学术交流使用,未涉及任何商业用途。由于小编水平有限,文中如有不当或不准确之处,敬请批评指正。如有疑问,欢迎随时后台联系小编交流探讨,若有侵权和冒犯请联系删除:)
点击文末👇“阅读原文”即可通过超链接跳转阅读在线文献




 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
